
Effects of the Bradyrhizobium japonicum waaL (rfaL) Gene on Hydrophobicity, Motility, Stress Tolerance, and Symbiotic Relationship with Soybeans

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the B. japonicum waaL (rfaL) Gene
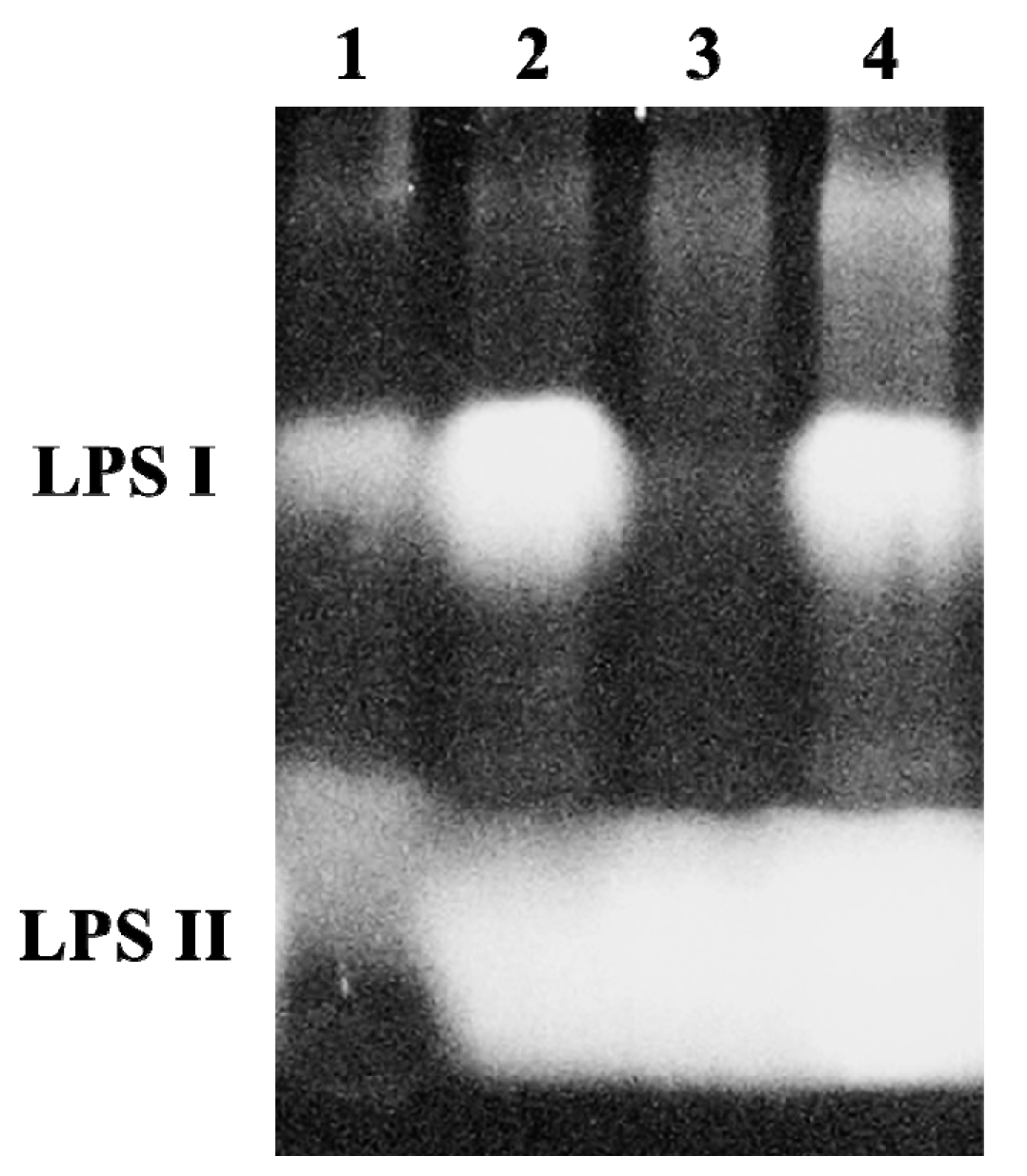
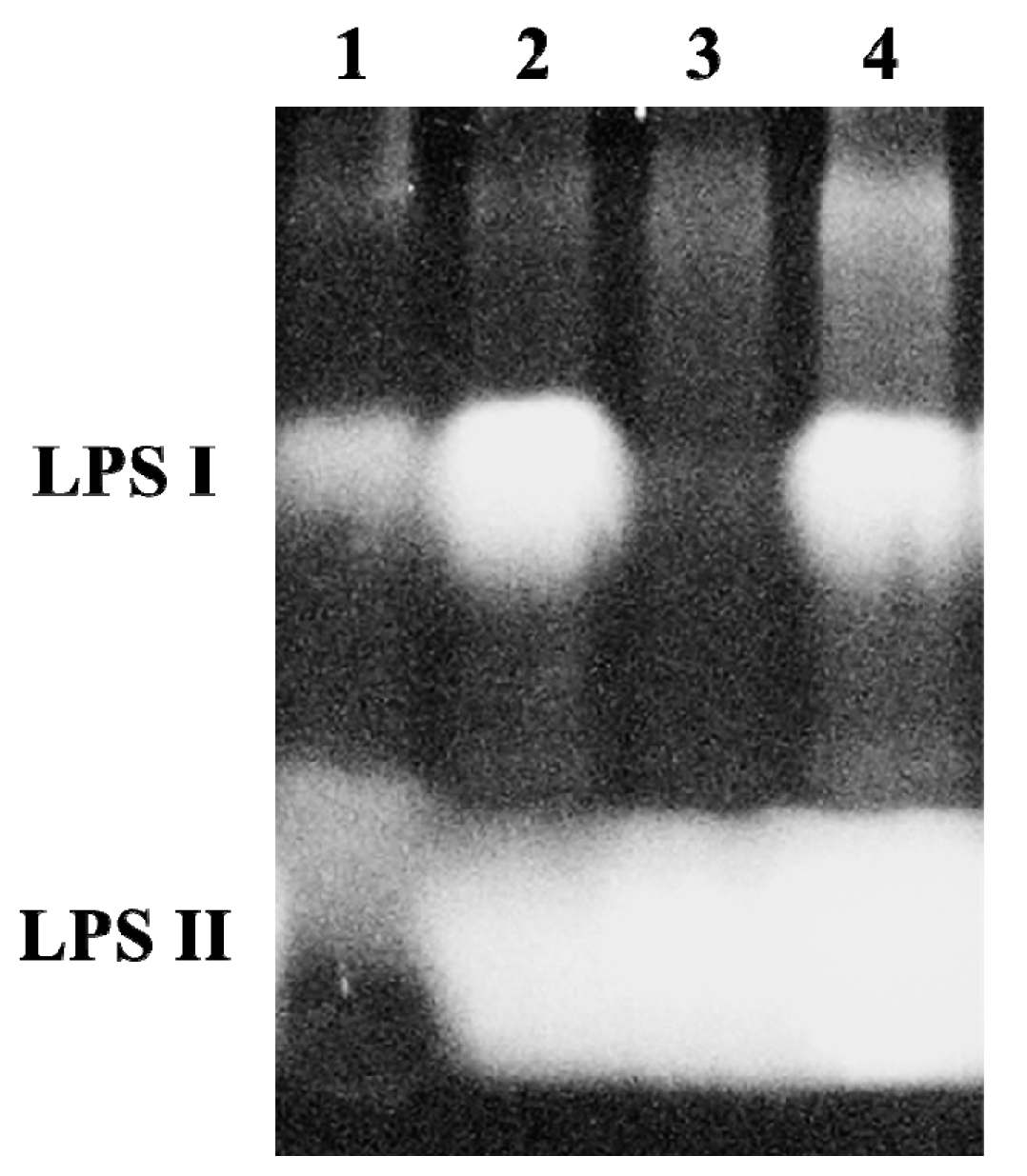
2.2. A B. japonicum waaL Mutant Shows Different LPS Profiles
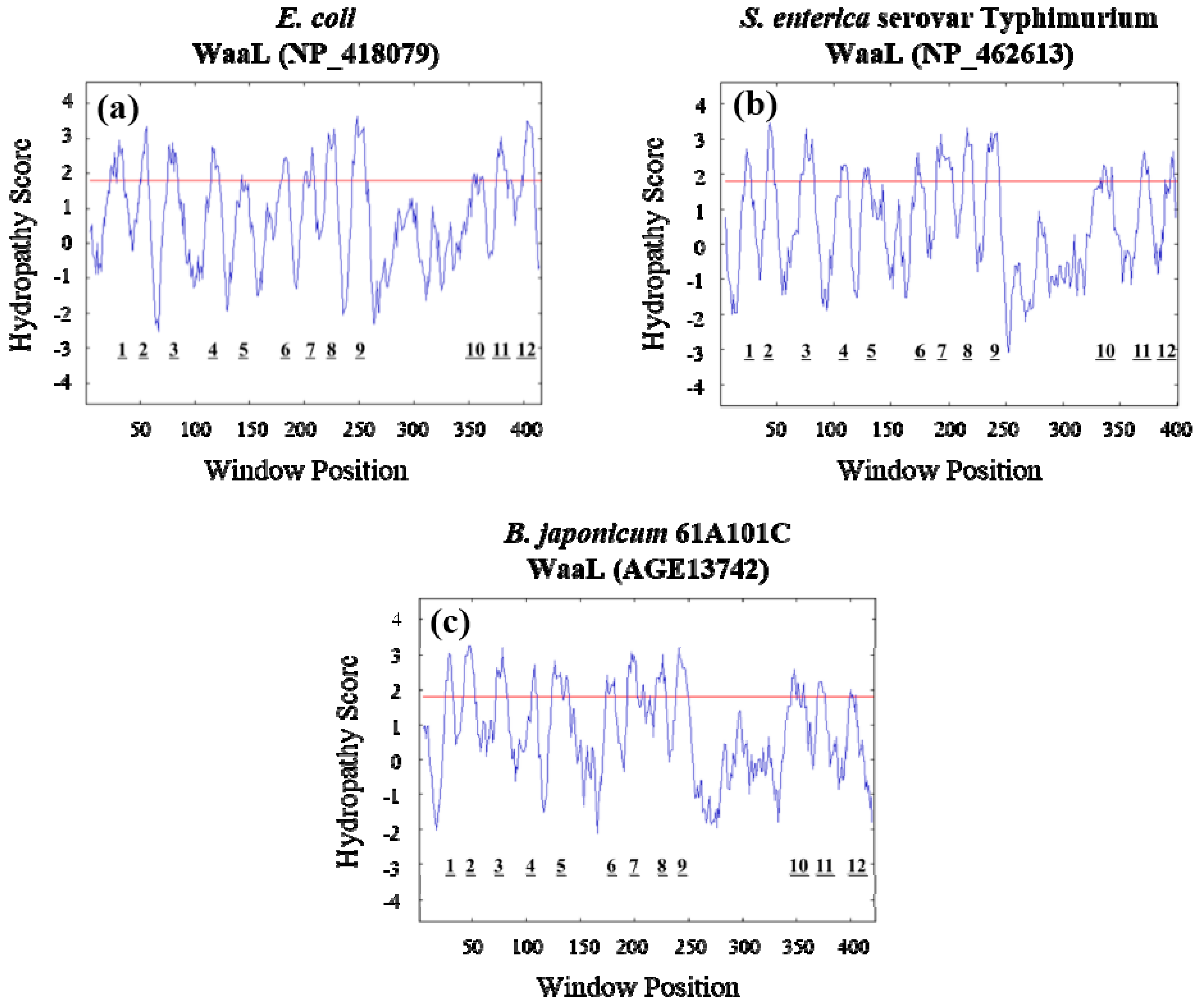
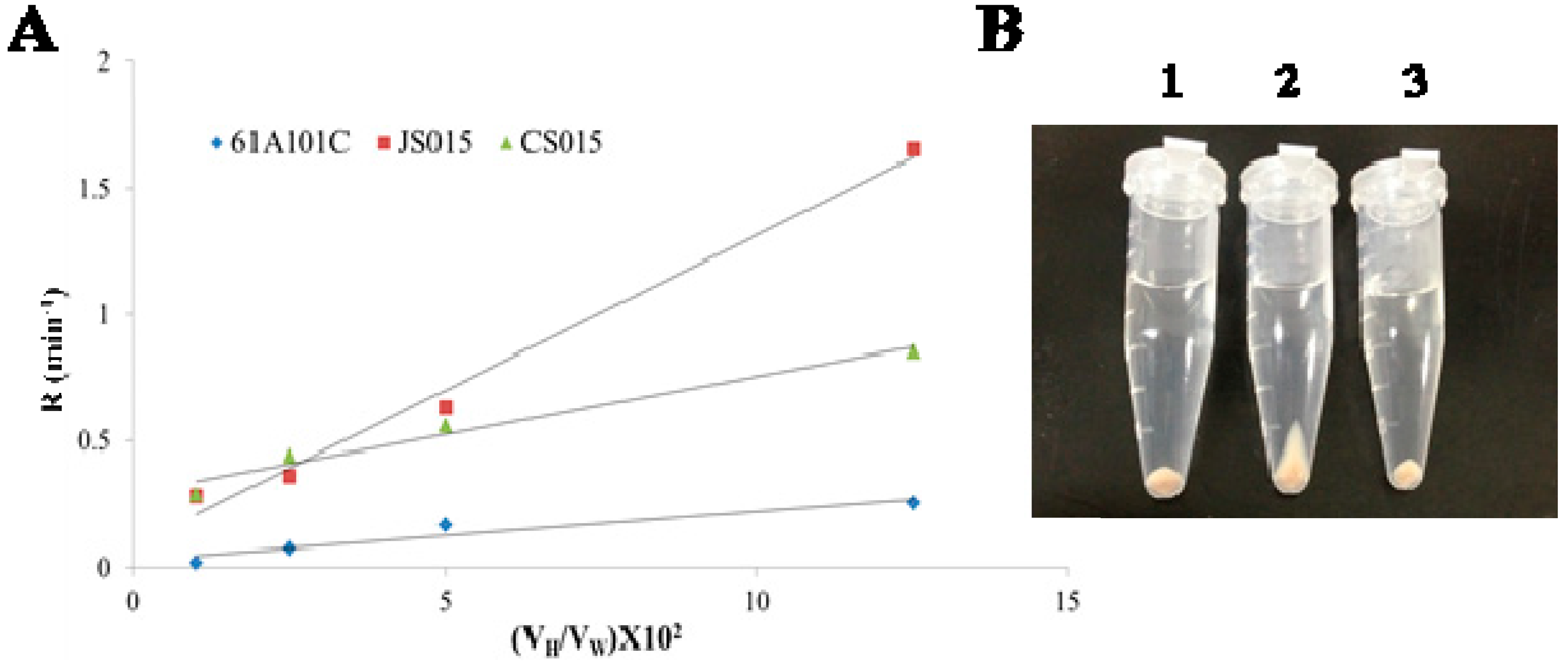
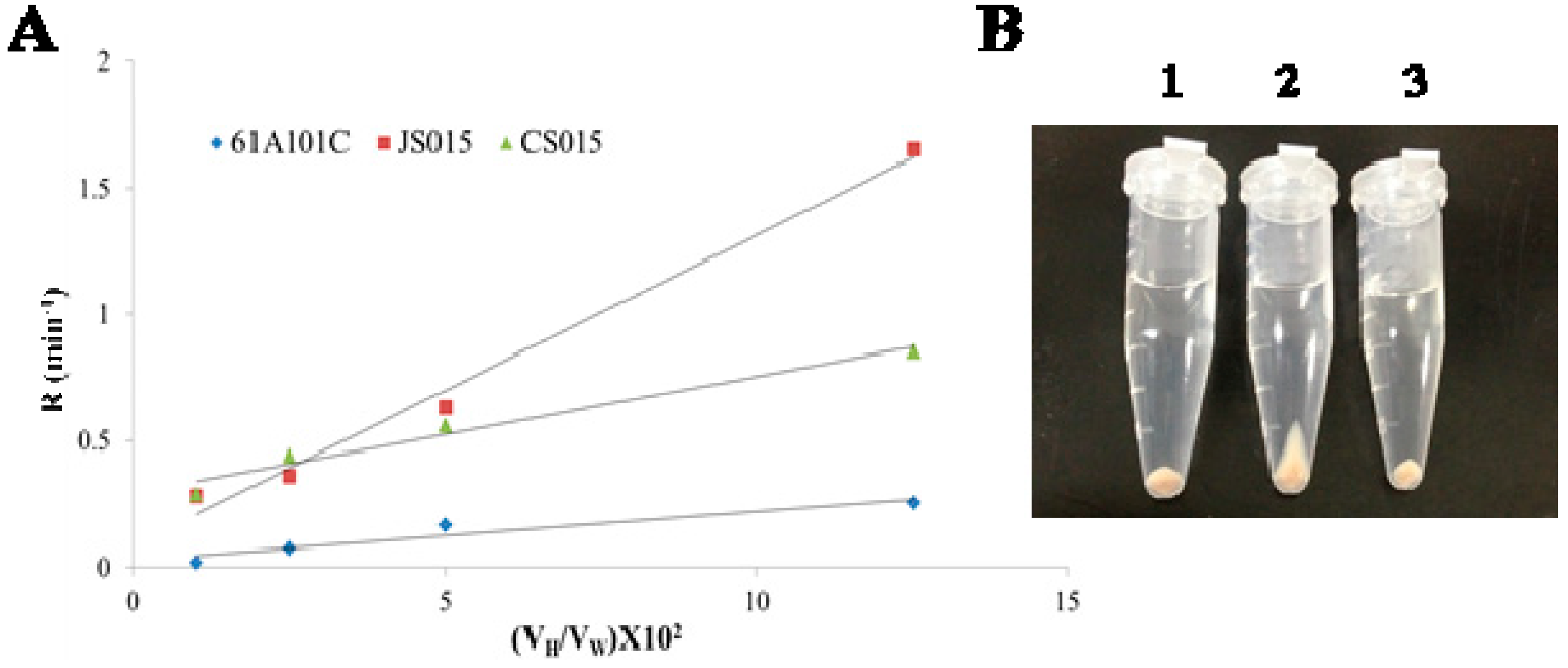
2.3. Lack of O-Antigen Increases the Cell Surface Hydrophobicity
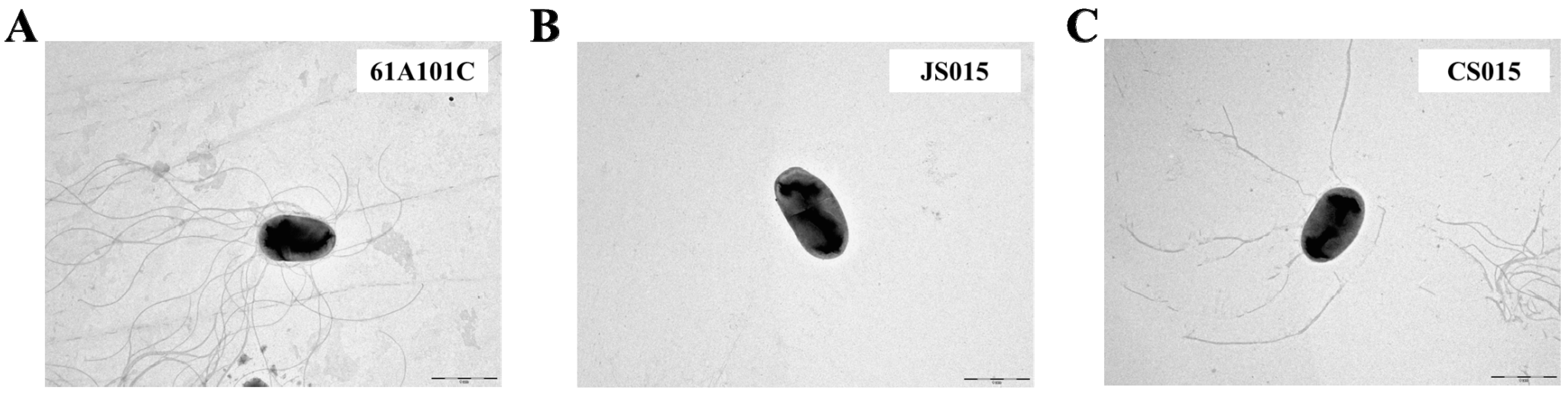
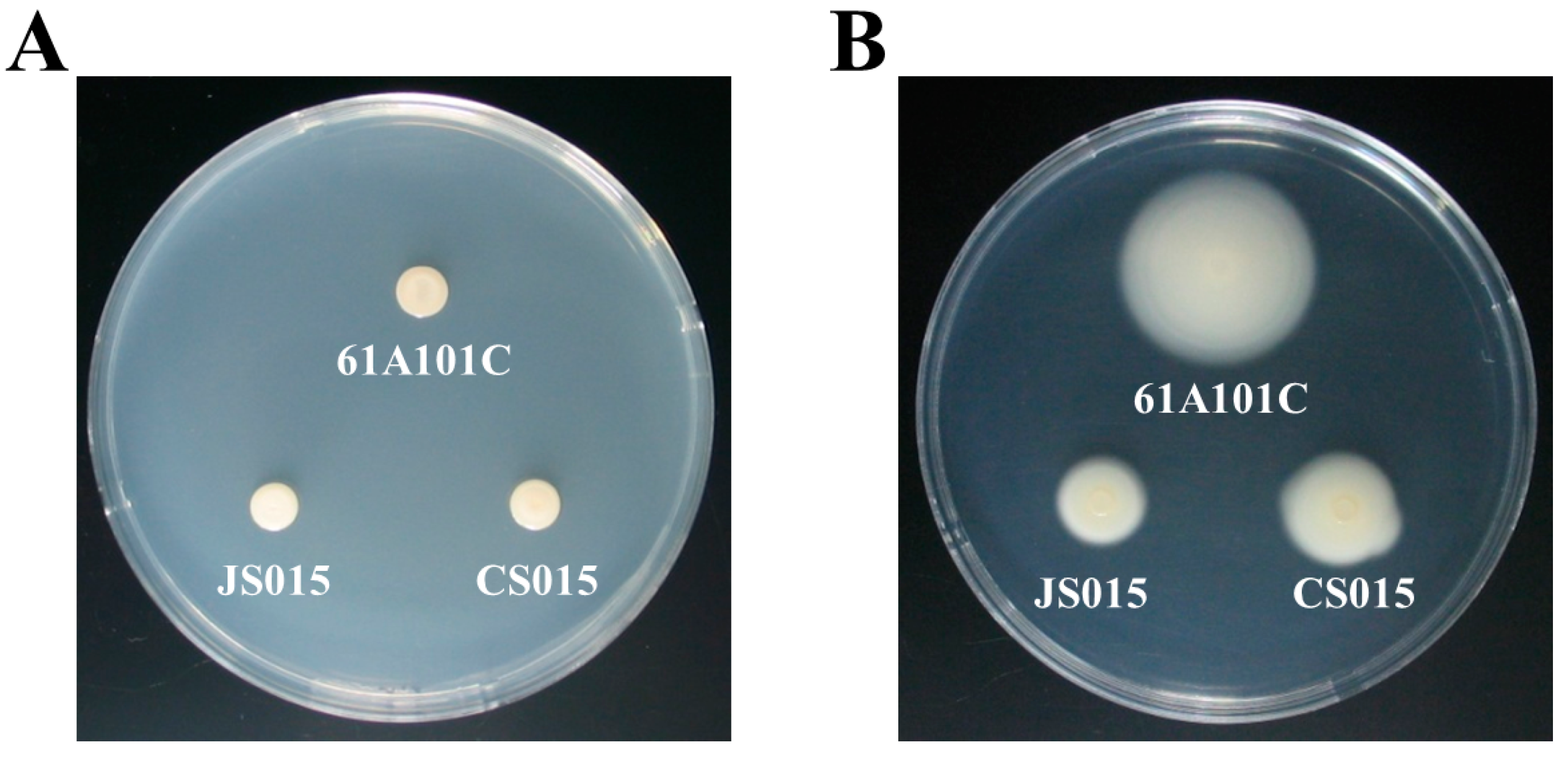
2.4. The waaL Mutant Is Less Motile

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Motility (mm in Diameter) | Osmotic Stress a (% Survivability) | Oxidative Stress b (Zone Inhibition Size in mm) | Novobiocin a (% Survivability) | Nodulation c |
|---|---|---|---|---|---|
| 61A101C | 33.0 ± 1.5 A | 106.4 ± 5.1 A | 30.7 ± 0.4 A | 92.5 ± 3.4 A | + |
| JS015 | 12.3 ± 0.9 B | 94.2 ± 4.2 B | 39.0 ± 0.7 B | 64.7 ± 9.0 B | − |
| CS015 | 20.8 ± 0.6 C | 109.2 ± 7.1 A | 34.7 ± 2.2 A | 88.7 ± 7.1 A | + |
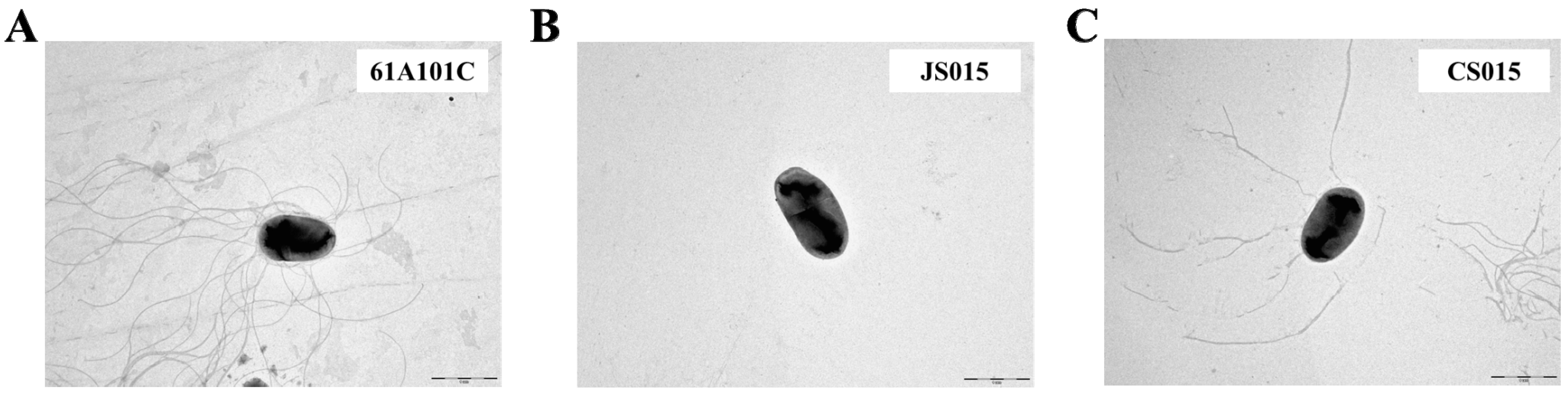
2.5. The waaL Mutant Is More Sensitive to Oxidative, Osmotic, and Novobiocin Stresses
2.6. waaL Is Required for Successful Nodule Formation on Soybean Roots
3. Experimental Section
3.1. Bacterial Strains, Plasmids, and Growth Conditions
| Strains/Plasmids | Relevant Characteristics | Reference or Source |
|---|---|---|
| B. japonicum | ||
| 61A101C | Wild type, Cmr | Nitragin (Milwaukee, WI, USA) |
| JS015 | Strain 61A101C waaL::Km, Kmr | This study |
| CS015 | JS015/pBBRL, Smr, Kmr | This study |
| E. coli | ||
| DH5α | supE44, ΔlacU169 (Φ80 lacZΔM15), hsdR17, recA1, endA1, gyrA96, thi-1, relA1 | Invitrogen (Carlsbad, CA, USA) |
| S17-1 | recA pro (RP4-2Tet::Mu Kan::Tn7) | [39] |
| Plasmids | ||
| pGEM-T easy | PCR cloning vector, Ampr | Promega (Madison, WI, USA) |
| p34s-Km | Vector containing Kmr cassette, Kmr | [40] |
| pTRL | pGEM T easy vector containing 1.5 kb fragment of waaL gene, Ampr | - |
| pTRLK | pTRL derivative waaL gene disruption by deletion of BssHII fragment and insertion of Kmr cassette, Kmr | - |
| pJQ200SK | Suicide vector, sacB, Gmr | [41] |
| pJQRLK | pJQ200SK-waaL::Km, Gmr, Kmr | This study |
| pIJ778 | pBluescript KS(+) derivative containing the streptomycin/spectinomycin resistance gene aadA, Smr | [42] |
| pBBR1MCS-2 | Broad host range expression vector, mod, rep, lacZ, Kmr | [43] |
| pBBRL | pBBR1MCS-2 carrying the 1.5 kb fragment of waaL gene, Kmr | This study |
| pBBRL-Sm | pBBRL derivative containing the Smr gene, Kmr gene disruption by deletion of NcoI fragment, Smr | This study |
3.2. DNA Sequence Analysis
3.3. Construction of a waaL Mutant and Its Complemented Strain
3.4. LPS Profile Analysis
3.5. Cell Surface Hydrophobicity Analysis
3.6. Motility and Transmission Electron Microscope (TEM) Analysis
3.7. H2O2 Inhibition Zone Assay
3.8. Osmotic Stress and Novobiocin Sensitivity Assay
3.9. Nodulation Assay
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stacey, G.; So, J.-S.; Roth, L.E.; Lakshmi, S.B.; Carlson, R.W. A lipopolysaccharide mutant of Bradyrhizobium japonicum that uncouples plant from bacterial differentiation. Mol. Plant Microbe Interact. 1991, 4, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; So, J.-S. Altered cell surface hydrophobicity of lipopolysaccharide-deficient mutant of Bradyrhizobium japonicum. J. Microbiol. Methods 2000, 41, 219–226. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwal, K.; Marzec, A.; Grzadziel, J.; Palusinska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- Kobayashi, H.; Naciri-Graven, Y.; Broughton, W.J.; Perret, X. Flavonoids induce temporal shifts in gene-expression of nod-box controlled loci in Rhizobium sp. NGR234. Mol. Microbiol. 2004, 51, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Lang, K.; Lindemann, A.; Hauser, F.; Göttfert, M. The genistein stimulon of Bradyrhizobium japonicum. Mol. Genet. Genomics 2008, 279, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Halverson, L.J.; Stacey, G. Signal exchange in plant-microbe interactions. Microbiol. Rev. 1986, 50, 193–225. [Google Scholar] [PubMed]
- Hirsch, A.M.; Fujishige, N.A. Molecular signals and receptors: Communication between nitrogen-fixing bacteria and their plant hosts. In Biocommunication of Plants; Witzany, G., Baluska, F., Eds.; Springer: Berlin, Germany, 2012; pp. 255–280. [Google Scholar]
- Hassan, S.; Mathesius, U. The role of flavonoids in root-rhizosphere signaling: Opportunities and challenges for improving plant-microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [PubMed]
- Carrion, M.; Bhat, U.R.; Reuhs, B.; Carlson, R.W. Isolation and characterization of the lipopolysaccharides from Bradyrhizobium japonicum. J. Bacteriol. 1990, 172, 1725–1731. [Google Scholar] [PubMed]
- Carlson, R.W.; Kalembasa, S.; Turowski, D.; Pachori, P.; Noel, K.D. Characterization of the lipopolysaccharide from a Rhizobium phaseoli mutant that is defective in infection thread development. J. Bacteriol. 1987, 169, 4923–4928. [Google Scholar] [PubMed]
- Campbell, G.R.; Reuhs, B.L.; Walker, G.C. Chronic intracellular infection of alfalfa nodules by Sinorhizobium meliloti requires correct lipopolysaccharide core. Proc. Natl. Acad. Sci. USA 2002, 99, 3938–3943. [Google Scholar] [CrossRef] [PubMed]
- Cava, J.R.; Elias, P.M.; Turowski, D.A.; Noel, K.D. Rhizobium leguminosarum CFN42 genetic regions encoding lipopolysaccharide structures essential for complete nodule development on bean plants. J. Bacteriol. 1989, 171, 8–15. [Google Scholar] [PubMed]
- Noel, K.D.; Duelli, D.M. Rhizobium lipopolysaccharide and its role in symbiosis. In Prokaryotic Nitrogen Fixation: A Model System for Analysis of a Biological Process; Triplett, E.W., Ed.; Horizon Scientific Press: Wymondham, UK, 2000; pp. 415–431. [Google Scholar]
- So, J.-S. Molecular cloning of a gene region from Bradyrhizobium japonicum essential for lipopolysaccharide synthesis. FEMS Microbiol. Lett. 1991, 83, 299–303. [Google Scholar] [CrossRef]
- So, J.-S.; Kim, W.-S.; Stacey, G. Molecular characterization of a gene region involved in lipopolysaccharide biosynthesis in Bradyrhizobium japonicum: Cloning, sequencing and expression of rfaF gene. FEMS Microbiol. Lett. 2000, 190, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.; Kim, D.-H.; Oh, E.-T.; So, J.-S. Characterization of the rfaD gene region of Bradyrhizobium japonicum 61A101C. J. Microbiol. Biotechnol. 2002, 12, 826–828. [Google Scholar]
- Chang, W.-S.; Park, K.-M.; Koh, S.-C.; So, J.-S. Characterization of the Bradyrhizobium japonicum galE gene: Its impact on lipopolysaccharide profile and nodulation of soybean. FEMS Microbiol. Lett. 2008, 280, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-I.; In, Y.-H.; Jeong, S.-Y.; Jeon, J.-M.; So, J.-S.; Chang, W.-S. Inactivation of the lpcC gene alters surface-related properties and symbiotic capability of Bradyrhizobium japonicum. Lett. Appl. Microbiol. 2014, 59, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Schnaitman, C.A.; Klena, J.D. Genetics of lipopolysaccharide biosynthesis in enteric bacteria. Microbiol. Rev. 1993, 57, 655–682. [Google Scholar] [PubMed]
- Klena, J.D.; Pradel, E.; Schnaitman, C.A. Comparison of lipopolysaccharide biosynthesis genes rfaK, rfaL, rfaY, and rfaZ of Escherichia coli K-12 and Salmonella typhimurium. J. Bacteriol. 1992, 174, 4746–4752. [Google Scholar] [PubMed]
- Ormeno-Orrillo, E.; Menna, P.; Almeida, L.G.P.; Ollero, F.J.; Nicolas, M.F.; Rodrigues, E.P.; Nakatani, A.S.; Batista, J.S.S.; Chueire, L.M.O.; Souza, R.C.; et al. Genomic basis of broad host range and environmental adaptability of Rhizobium tropici CIAT 899 and Rhizobium sp. PRF 81 which are used in inoculants for common bean (Phaseolus vulgaris L.). BMC Genomics. 2012, 13, 735. [Google Scholar] [PubMed]
- Post, D.-M.; Yu, L.; Krasity, B.C.; Choudhury, B.; Mandel, M.J.; Brennan, C.A.; Ruby, E.G.; McFall-Ngai, M.J.; Gibson, B.W.; Apicella, M.A. O-antigen and core carbohydrate of Vibrio fischeri lipopolysaccharide composition and analysis of their role in Euprymna scolopes light organ colonization. J. Biol. Chem. 2012, 287, 8515–8530. [Google Scholar] [CrossRef] [PubMed]
- Abeyrathne, P.D.; Daniels, C.; Poon, K.K.; Matewish, M.J.; Lam, J.S. Functional characterization of WaaL, a ligase associated with linking O-antigen polysaccharide to the core of Pseudomonas aeruginosa lipopolysaccharide. J. Bacteriol. 2005, 187, 3002–3012. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.C.; McGhee, G.C.; Zhao, Y.; Sundin, G.W. Effect of a waaL mutation on lipopolysaccharide composition, oxidative stress survival, and virulence in Erwinia amylovora. FEMS Microbiol. Lett. 2009, 291, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, D.E.; Yethon, J.A.; Whitfield, C. Molecular basis for structural diversity in the core regions of the lipopolysaccharides of Escherichia coli and Salmonella enterica. Mol. Microbiol. 1998, 30, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Morgenstein, R.-M.; Clemmer, K.M.; Rather, P.N. Loss of the WaaL O-antigen ligase prevents surface activation of the flagellar gene cascade in Proteus mirabilis. J. Bacteriol. 2010, 192, 3213–3221. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-W.; Jeong, S.-Y.; In, Y.-H.; kim, K.-Y.; So, J.-S.; Chang, W.-S. Lack of O-polysaccharide enchances biofilm formaion by Bradyrhizobium japonicum. Lett. Appl. Microbiol. 2010, 50, 452–456. [Google Scholar]
- Chang, W.-S.; Franck, W.L.; Cytryn, E.; Jeong, S.; Joshi, T.; Emerich, D.W.; Sadowsky, M.J.; Xu, D.; Stacey, G. An oligonucleotide microarray resource for transcriptional profiling of Bradyrhizobium japonicum. Mol. Plant Microbe Interact. 2007, 20, 1298–1309. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.A.; Armario, P.; Pajuelo, E.; Burgos, A.; Caviedes, M.A.; Lopez, R.; Chamber, M.A.; Palomares, A.J. Isolation and characterisation of symbiotically effective Rhizobium resistant to arsenic and heavy metals after the toxic spill at the Aznalcollar pyrite mine. Soil Biol. Biochem. 2005, 37, 1131–1140. [Google Scholar] [CrossRef]
- Donati, A.J.; Jeon, J.-M.; Sangurdekar, D.; So, J.-S.; Chang, W.-S. The genome-wide transcriptional and physiological responses of Bradyrhizobium japonicum to paraquat-mediated oxidative stress. Appl. Environ. Microbiol. 2011, 77, 3633–3643. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-M.; Lee, H.-I.; Donati, A.J.; So, J.-S.; Emerich, D.W.; Chang, W.-S. Whole-genome expression profiling of Bradyrhizobium japonicum in response to hydrogen peroxide. Mol. Plant Microbe Interact. 2011, 24, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Vriezen, J.A.C.; de Bruijn, F.J.; Nusslein, K. Responses of rhizobia to desiccation in relation to osmotic stress, oxygen, and temperature. Appl. Environ. Microbiol. 2007, 73, 3451–3459. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.A.W.; Bending, G.D.; White, P.J. Biological costs and benefits to plant-microbe interactions in the rhizosphere. J. Exp. Bot. 2005, 56, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Munoz, V.; Ibanez, F.; Tordable, M.; Megias, M.; Fabra, A. Role of reactive oxygen species generation and Nod factors during the early symbiotic interaction between bradyrhizobia and peanut, a legume infected by crack entry. J. Appl. Microbiol. 2014, 118, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Singleton, P.W.; Bohlool, B.B. Effect of salinity on nodule formation by soybean. Plant Physiol. 1984, 74, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.; Robert, G.; Melchiorre, M.; Racca, R.; Lascano, R. Saline and osmotic stress differentially affects apoplastic and intracellular reactive oxygen species production, curling and death of root hair during Glycine max L.–Bradyrhizobium japonicum interaction. Environ. Exp. Bot. 2012, 78, 76–83. [Google Scholar] [CrossRef]
- Tamaki, S.; Sato, T.; Matsuhashi, M. Role of lipopolysaccharides in antibiotic resistance and bacteriophage adsorption of Escherichia coli K-12. J. Bacteriol. 1971, 105, 968–975. [Google Scholar] [PubMed]
- Sadowsky, M.J.; Tully, R.E.; Cregan, P.B.; Keyser, H.H. Genetic diversity in Bradyrhizobium japonicum serogroup 123 and its relation to genotype-specific nodulation of soybean. Appl. Environ. Microbiol. 1987, 53, 2624–2630. [Google Scholar] [PubMed]
- Simon, R.; Priefer, U.; Puhler, A. A braod host range mobilization system for in vivo genetic engineering—Transcposon mutagenesis in Gram-negative bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Dennis, J.J.; Zylstra, G.J. Plasposons: Modular self-cloning minitransposon derivatives for rapid genetic analysis of gram-negative bacterial genomes. Appl. Environ. Microbiol. 1998, 64, 2710–2715. [Google Scholar] [PubMed]
- Quandt, J.; Hynes, M.F. Versatile suicide vectors which allow direct selection for gene replacement in gram-negative bacteria. Gene 1993, 127, 15–21. [Google Scholar] [CrossRef]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M., II; Peterson, K. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons—A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Lee, H.-I.; Lee, J.-S.; Park, K.-H.; Sangurdekar, D.; Chang, W.-S. Effect of soybean coumestrol on Bradyrhizobium japonicum nodulation ability, biofilm formation, and transcriptional profile. Appl. Environ. Microbiol. 2012, 78, 2896–2903. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.; Dilworth, M. Control of leghaemoglobin synthesis in snake beans. Biochem. J. 1971, 125, 1075–1080. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, J.-G.; Jeon, H.-E.; So, J.-S.; Chang, W.-S. Effects of the Bradyrhizobium japonicum waaL (rfaL) Gene on Hydrophobicity, Motility, Stress Tolerance, and Symbiotic Relationship with Soybeans. Int. J. Mol. Sci. 2015, 16, 16778-16791. https://doi.org/10.3390/ijms160816778
Noh J-G, Jeon H-E, So J-S, Chang W-S. Effects of the Bradyrhizobium japonicum waaL (rfaL) Gene on Hydrophobicity, Motility, Stress Tolerance, and Symbiotic Relationship with Soybeans. International Journal of Molecular Sciences. 2015; 16(8):16778-16791. https://doi.org/10.3390/ijms160816778
Chicago/Turabian StyleNoh, Jun-Gu, Han-Eul Jeon, Jae-Seong So, and Woo-Suk Chang. 2015. "Effects of the Bradyrhizobium japonicum waaL (rfaL) Gene on Hydrophobicity, Motility, Stress Tolerance, and Symbiotic Relationship with Soybeans" International Journal of Molecular Sciences 16, no. 8: 16778-16791. https://doi.org/10.3390/ijms160816778
APA StyleNoh, J.-G., Jeon, H.-E., So, J.-S., & Chang, W.-S. (2015). Effects of the Bradyrhizobium japonicum waaL (rfaL) Gene on Hydrophobicity, Motility, Stress Tolerance, and Symbiotic Relationship with Soybeans. International Journal of Molecular Sciences, 16(8), 16778-16791. https://doi.org/10.3390/ijms160816778