
Oxidized LDL Is Strictly Limited to Hyperthyroidism Irrespective of Fat Feeding in Female Sprague Dawley Rats

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Triglycerides, Free Fatty Acids, Phospholipids, HDL, Cholesterol
2.2. Malondialdehyde (MDA), 4-Hydroxynonenal (HNE)
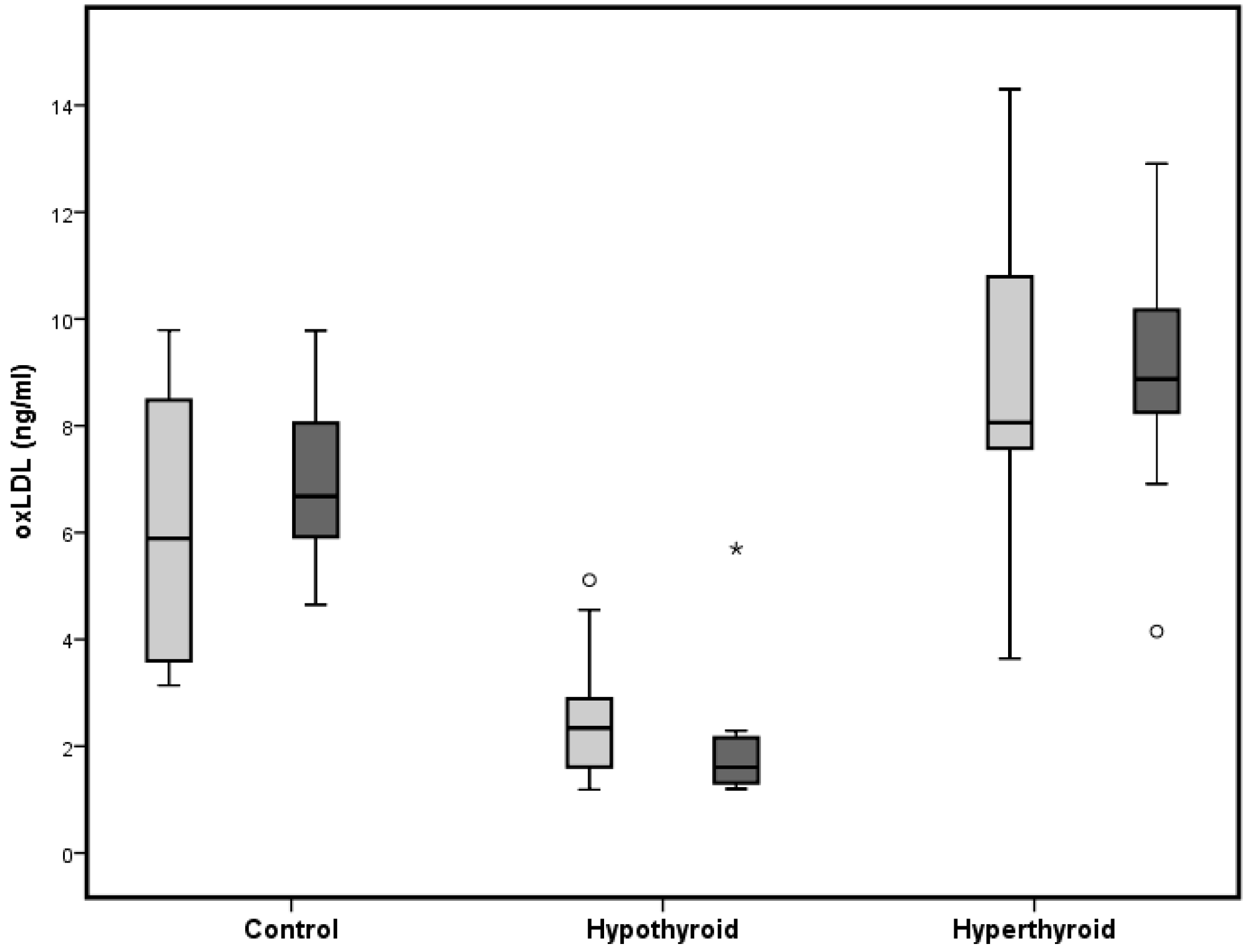
2.3. Oxidized Low Dense Lipoprotein (oxLDL)

{kind=link}
| Normal Diet Group (n = 30) | High Fat Diet Group (n = 36) | |||||
|---|---|---|---|---|---|---|
| Control (n = 10) | Hypothyroid (n = 10) | Hyperthyroid (n = 10) | Control (n = 10) | Hypothyroid (n = 10) | Hyperthyroid (n = 16) | |
| Body weight change (g) | 36 ± 24 | −35 ± 8 *** | 31 ± 7 | 56 ± 24 | −42 ± 16 *** | 49 ± 17 †† |
| Body weight change (%) | 14 ± 10 | −13 ± 2 *** | 12 ± 3 | 21 ± 9 | −15 ± 5 *** | 19 ± 7 |
| Food intake/rate/day (g) | 19 | 10 | 30 | 13 | 8 | 18 |
| Calorie intake/rat/day (kcal) | 49 | 26 | 80 | 61 | 38 | 87 |
| T3 (pg/mL) | 417 ± 85 | 269 ± 74 ** | 951 ± 454 ** | 450 ± 58 | 446 ± 176 †† | 1708 ± 1447 |
| Oxidized LDL (ng/mL) | 6.1 ± 2.5 | 2.6 ± 1.3 ** | 8.8 ± 3.0 * | 6.9 ± 1.6 | 2.1 ± 1.3 *** | 9.5 ± 3.0 * |
| MDA (nmol/mL) | 1.4 ± 0.2 | 1.4 ± 0.3 | 1.0 ± 0.2 *** | 1.0 ± 0.1 ººº | 0.7 ± 0.1 ††† | 1.0 ± 0.3 |
| HNE (nmol/mL) | 0.09 ± 0.02 | 0.09 ± 0.03 | 0.12 ± 0.05 | 0.08 ± 0.02 | 0.07 ± 0.02 | 0.11 ± 0.04 |
| Triglyceride (mg/dL) | 62 ± 8 | 46 ± 4 *** | 61 ± 36 | 50 ± 12 º | 32 ± 6 ††† | 88 ± 14 |
| Cholesterol (mg/dL) | 86 ± 13 | 87 ± 14 | 69 ± 17 * | 81 ± 14 | 185 ± 23 ††† | 64 ± 13 |
| Free cholesterol (mg/dL) | 24 ± 4 | 26 ± 5 | 19 ± 6 * | 20 ± 5 | 59 ± 8 ††† | 18 ± 7 |
| Phospholipids (mg/dL) | 156 ± 15 | 120 ± 19 ** | 141 ± 21 | 139 ± 22 | 209 ± 21 ††† | 129 ± 26 |
| Free fatty acids (mmol/L) | 1.1 ± 0.2 | 0.7 ± 0.2 ** | 1.0 ± 0.3 | 0.8 ± 0.1 ºº | 0.5 ± 0.1 | 1.2 ± 0.5 |
| HDL cholesterol (mg/dL) | 44 ± 5 | 39 ± 6 | 41 ± 7 | 44 ± 5 | 66 ± 5 ††† | 40 ± 9 |
3. Discussion
4. Experimental Section
4.1. Animals
4.2. Experimental Design and Treatment
4.3. Blood Collection
4.4. Laboratory Procedures
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Highlights
- (1)
- LDL oxidation is strictly driven by hyperthyroidism.
- (2)
- The above effect persists independently of diet.
Conflicts of Interest
References
- Kratky, J.; Vitkova, H.; Bartakova, J.; Telicka, Z.; Antosova, M.; Limanova, Z.; Jiskra, J. Thyroid nodules: Pathophysiological insight on oncogenesis and novel diagnostic techniques. Physiol. Res. 2014, 63, S263–S275. [Google Scholar] [PubMed]
- Medici, M.; Porcu, E.; Pistis, G.; Teumer, A.; Brown, S.J.; Jensen, R.A.; Rawal, R.; Roef, G.L.; Plantinga, T.S.; Vermeulen, S.H.; et al. Identification of novel genetic Loci associated with thyroid peroxidase antibodies and clinical thyroid disease. PLoS Genet. 2014, 10, e1004123. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, L.B.; Stewart, C.; Gaitonde, D.Y. Thyroiditis: An integrated approach. Am. Fam. Physician 2014, 90, 389–396. [Google Scholar] [PubMed]
- Yen, P.M. Physiological and molecular basis of thyroid hormone action. Physiol. Rev. 2001, 81, 1097–1142. [Google Scholar] [PubMed]
- Iwen, K.A.; Schroder, E.; Brabant, G. Thyroid hormones and the metabolic syndrome. Eur. Thyroid J. 2013, 2, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Peppa, M.; Betsi, G.; Dimitriadis, G. Lipid abnormalities and cardiometabolic risk in patients with overt and subclinical thyroid disease. J. Lipids 2011, 2011. [Google Scholar] [CrossRef]
- Santi, A.; Duarte, M.M.; Moresco, R.N.; Menezes, C.; Bagatini, M.D.; Schetinger, M.R.; Loro, V.L. Association between thyroid hormones, lipids and oxidative stress biomarkers in overt hypothyroidism. Clin. Chem. Lab. Med. 2010, 48, 1635–1639. [Google Scholar] [CrossRef] [PubMed]
- Tejovathi, B.; Suchitra, M.M.; Suresh, V.; Reddy, V.S.; Sachan, A.; Srinivas Rao, P.V.; Bitla, A.R. Association of lipid peroxidation with endothelial dysfunction in patients with overt hypothyroidism. Exp. Clin. Endocrinol. Diabetes 2013, 121, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Nanda, N.; Bobby, Z.; Hamide, A. Association of thyroid stimulating hormone and coronary lipid risk factors with lipid peroxidation in hypothyroidism. Clin. Chem. Lab. Med. 2008, 46, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, I.; Alva-Sanchez, C.; Pacheco-Rosado, J. The role of thyroid hormones as inductors of oxidative stress and neurodegeneration. Oxidative Med. Cell. Longev. 2013, 2013, 218145. [Google Scholar] [CrossRef]
- Venditti, P.; Balestrieri, M.; di Meo, S.; de Leo, T. Effect of thyroid state on lipid peroxidation, antioxidant defences, and susceptibility to oxidative stress in rat tissues. J. Endocrinol. 1997, 155, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Costantini, F.; Pierdomenico, S.D.; de Cesare, D.; de Remigis, P.; Bucciarelli, T.; Bittolo-Bon, G.; Cazzolato, G.; Nubile, G.; Guagnano, M.T.; Sensi, S.; et al. Effect of thyroid function on LDL oxidation. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Ittermann, T.; Baumeister, S.E.; Volzke, H.; Wasner, C.; Schminke, U.; Wallaschofski, H.; Nauck, M.; Ludemann, J. Are serum TSH levels associated with oxidized low-density lipoprotein? Results from the Study of Health in Pomerania. Clin. Endocrinol. 2012, 76, 526–532. [Google Scholar] [CrossRef]
- Lampka, M.; Junik, R.; Nowicka, A.; Kopczynska, E.; Tyrakowski, T.; Odrowaz-Sypniewska, G. Oxidative stress markers during a course of hyperthyroidism. Endokrynol. Pol. 2006, 57, 218–222. [Google Scholar] [PubMed]
- Messarah, M.; Boumendjel, A.; Chouabia, A.; Klibet, F.; Abdennour, C.; Boulakoud, M.S.; Feki, A.E. Influence of thyroid dysfunction on liver lipid peroxidation and antioxidant status in experimental rats. Exp. Toxicol. Pathol. 2010, 62, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Hussein Ael, A.; Abbas, A.M.; El Wakil, G.A.; Elsamanoudy, A.Z.; El Aziz, A.A. Effect of chronic excess iodine intake on thyroid function and oxidative stress in hypothyroid rats. Can. J. Physiol. Pharmacol. 2012, 90, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Rizos, C.V.; Elisaf, M.S.; Liberopoulos, E.N. Effects of thyroid dysfunction on lipid profile. Open Cardiovasc. Med. J. 2011, 5, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Kaliaperumal, R.; William, E.; Selvam, T.; Krishnan, S.M. Relationship between lipoprotein (a) and thyroid hormones in hypothyroid patients. J. Clin. Diagn. Res. 2014, 8, 37–39. [Google Scholar] [PubMed]
- Ragginer, C.; Bernecker, C.; Ainoedhofer, H.; Pailer, S.; Kieslinger, P.; Truschnig-Wilders, M.; Gruber, H.J. Treatment with the nitric oxide donor SNP increases triiodothyronine levels in hyper- and hypothyroid Sprague-Dawley rats. Horm. Metab. Res. 2013, 45, 808–812. [Google Scholar] [CrossRef] [PubMed]
- McAllister, R.M.; Albarracin, I.; Price, E.M.; Smith, T.K.; Turk, J.R.; Wyatt, K.D. Thyroid status and nitric oxide in rat arterial vessels. J. Endocrinol. 2005, 185, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Messarah, M.; Saoudi, M.; Boumendjel, A.; Boulakoud, M.S.; Feki, A.E. Oxidative stress induced by thyroid dysfunction in rat erythrocytes and heart. Environ. Toxicol. Pharmacol. 2011, 31, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Weltman, N.Y.; Ojamaa, K.; Savinova, O.V.; Chen, Y.F.; Schlenker, E.H.; Zucchi, R.; Saba, A.; Colligiani, D.; Pol, C.J.; Gerdes, A.M. Restoration of cardiac tissue thyroid hormone status in experimental hypothyroidism: A dose-response study in female rats. Endocrinology 2013, 154, 2542–2552. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, D.; Song, G.; Zuo, C.; Qiao, X.; Qin, S. The effect of combination therapy of allicin and fenofibrate on high fat diet-induced vascular endothelium dysfunction and liver damage in rats. Lipids Health Dis. 2010, 9. [Google Scholar] [CrossRef]
- Bhandari, U.; Kumar, V.; Khanna, N.; Panda, B.P. The effect of high-fat diet-induced obesity on cardiovascular toxicity in Wistar albino rats. Hum. Exp. Toxicol. 2011, 30, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Casellas, A.; Proenza, A.M.; Gianotti, M.; Llad, I. Brown adipose tissue redox status in response to dietary-induced obesity-associated oxidative stress in male and female rats. Stress 2011, 14, 174–184. [Google Scholar] [PubMed]
- Zelzer, S.; Oberreither, R.; Bernecker, C.; Stelzer, I.; Truschnig-Wilders, M.; Fauler, G. Measurement of total and free malondialdehyde by gas-chromatography mass spectrometry—Comparison with high-performance liquid chromatography methology. Free Radic. Res. 2013, 47, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.P.; Yazdanpanah, M.; Bhooi, N.; Lehotay, D.C. Determination of aldehydes and other lipid peroxidation products in biological samples by gas chromatography-mass spectrometry. Anal. Biochem. 1995, 228, 294–298. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zelzer, S.; Mangge, H.; Pailer, S.; Ainoedhofer, H.; Kieslinger, P.; Stojakovic, T.; Scharnagl, H.; Prüller, F.; Weghuber, D.; Datz, C.; et al. Oxidized LDL Is Strictly Limited to Hyperthyroidism Irrespective of Fat Feeding in Female Sprague Dawley Rats. Int. J. Mol. Sci. 2015, 16, 11689-11698. https://doi.org/10.3390/ijms160511689
Zelzer S, Mangge H, Pailer S, Ainoedhofer H, Kieslinger P, Stojakovic T, Scharnagl H, Prüller F, Weghuber D, Datz C, et al. Oxidized LDL Is Strictly Limited to Hyperthyroidism Irrespective of Fat Feeding in Female Sprague Dawley Rats. International Journal of Molecular Sciences. 2015; 16(5):11689-11698. https://doi.org/10.3390/ijms160511689
Chicago/Turabian StyleZelzer, Sieglinde, Harald Mangge, Sabine Pailer, Herwig Ainoedhofer, Petra Kieslinger, Tatjana Stojakovic, Hubert Scharnagl, Florian Prüller, Daniel Weghuber, Christian Datz, and et al. 2015. "Oxidized LDL Is Strictly Limited to Hyperthyroidism Irrespective of Fat Feeding in Female Sprague Dawley Rats" International Journal of Molecular Sciences 16, no. 5: 11689-11698. https://doi.org/10.3390/ijms160511689
APA StyleZelzer, S., Mangge, H., Pailer, S., Ainoedhofer, H., Kieslinger, P., Stojakovic, T., Scharnagl, H., Prüller, F., Weghuber, D., Datz, C., Haybaeck, J., Obermayer-Pietsch, B., Trummer, C., Gostner, J., & Gruber, H.-J. (2015). Oxidized LDL Is Strictly Limited to Hyperthyroidism Irrespective of Fat Feeding in Female Sprague Dawley Rats. International Journal of Molecular Sciences, 16(5), 11689-11698. https://doi.org/10.3390/ijms160511689