
Chemical Profiles and Protective Effect of Hedyotis diffusa Willd in Lipopolysaccharide-Induced Renal Inflammation Mice

Abstract
:1. Introduction
2. Results and Discussion
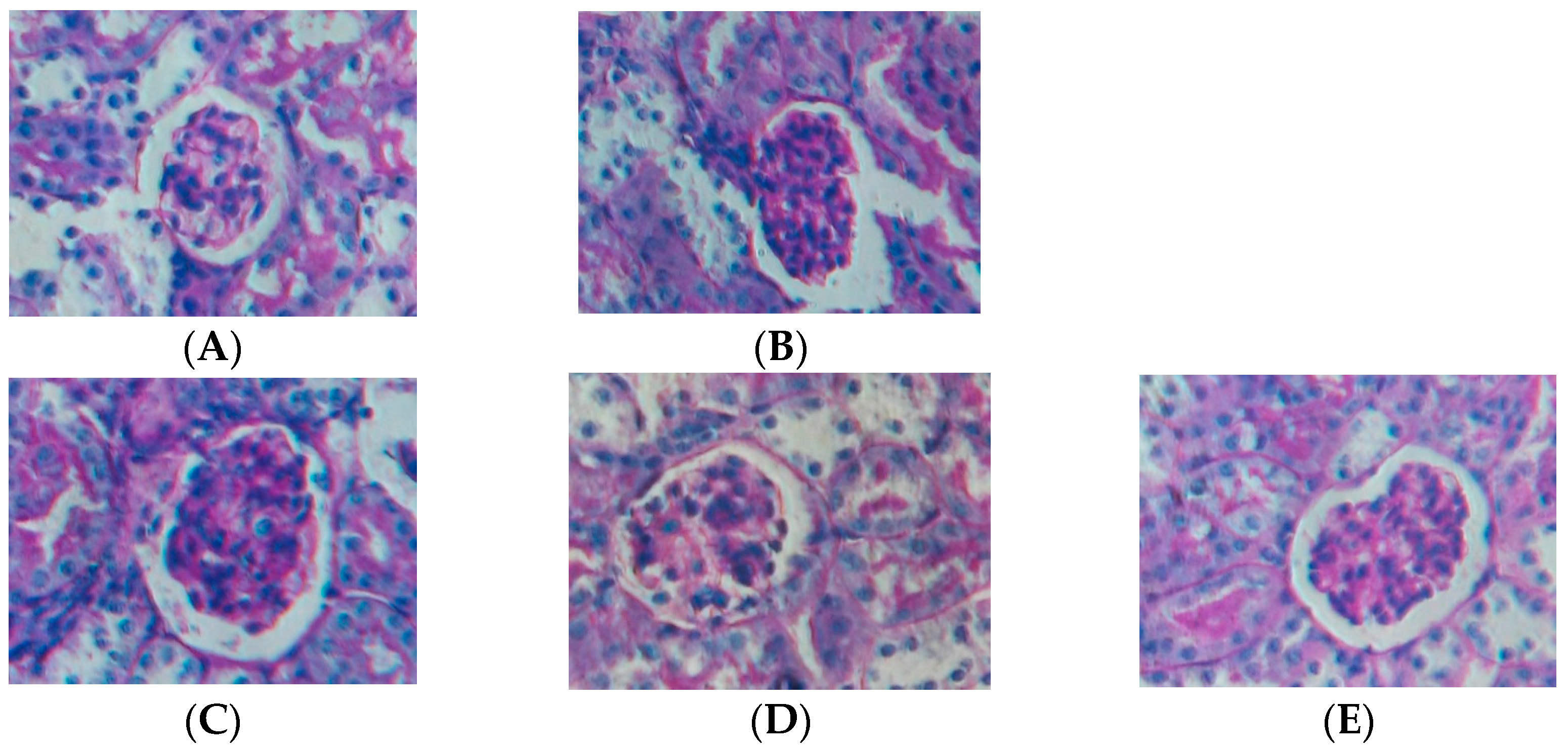
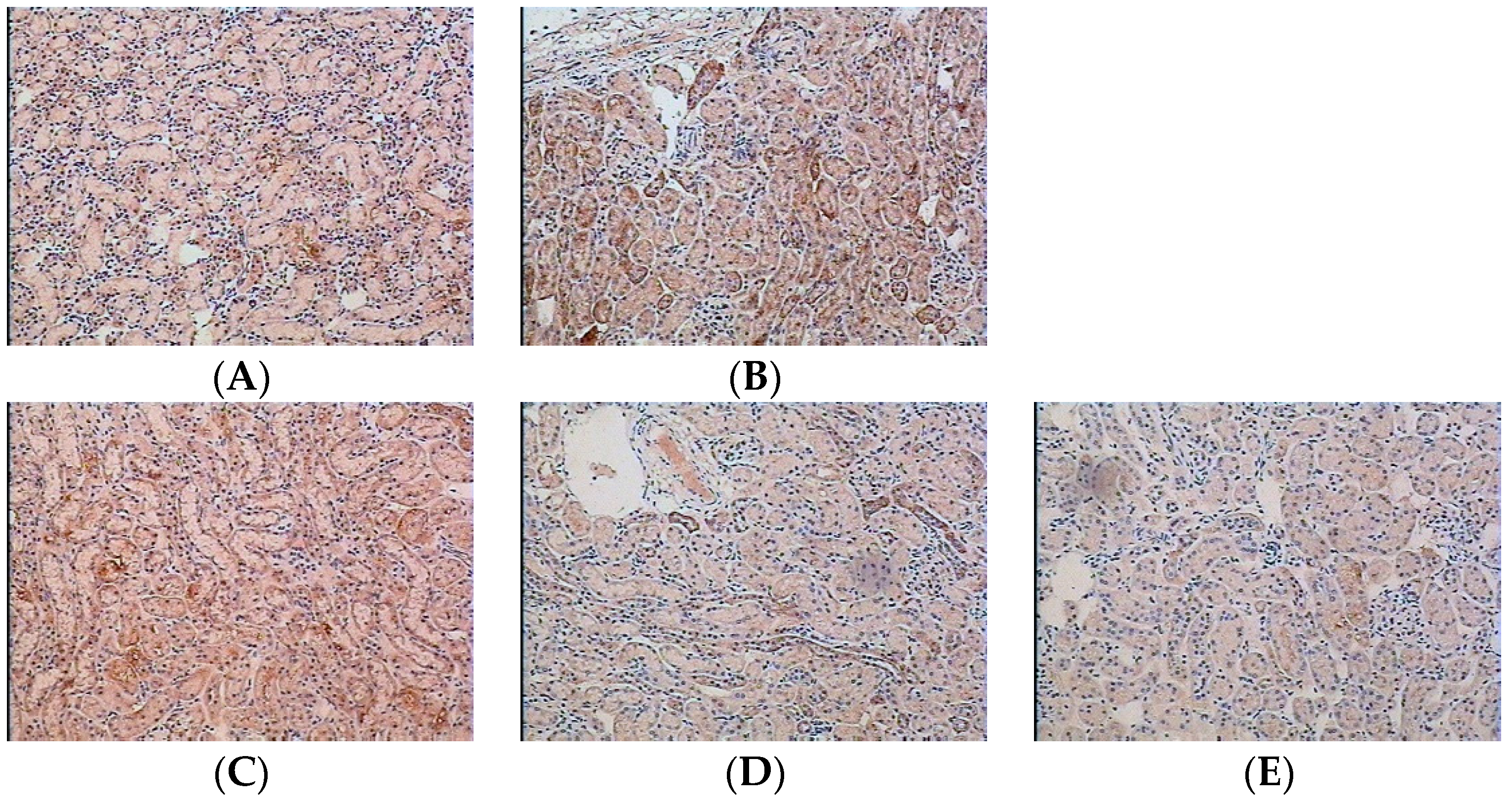
2.1. Effect of Water Extract of H. diffusa on the Histology of LPS-Induced Renal Inflammation Mice
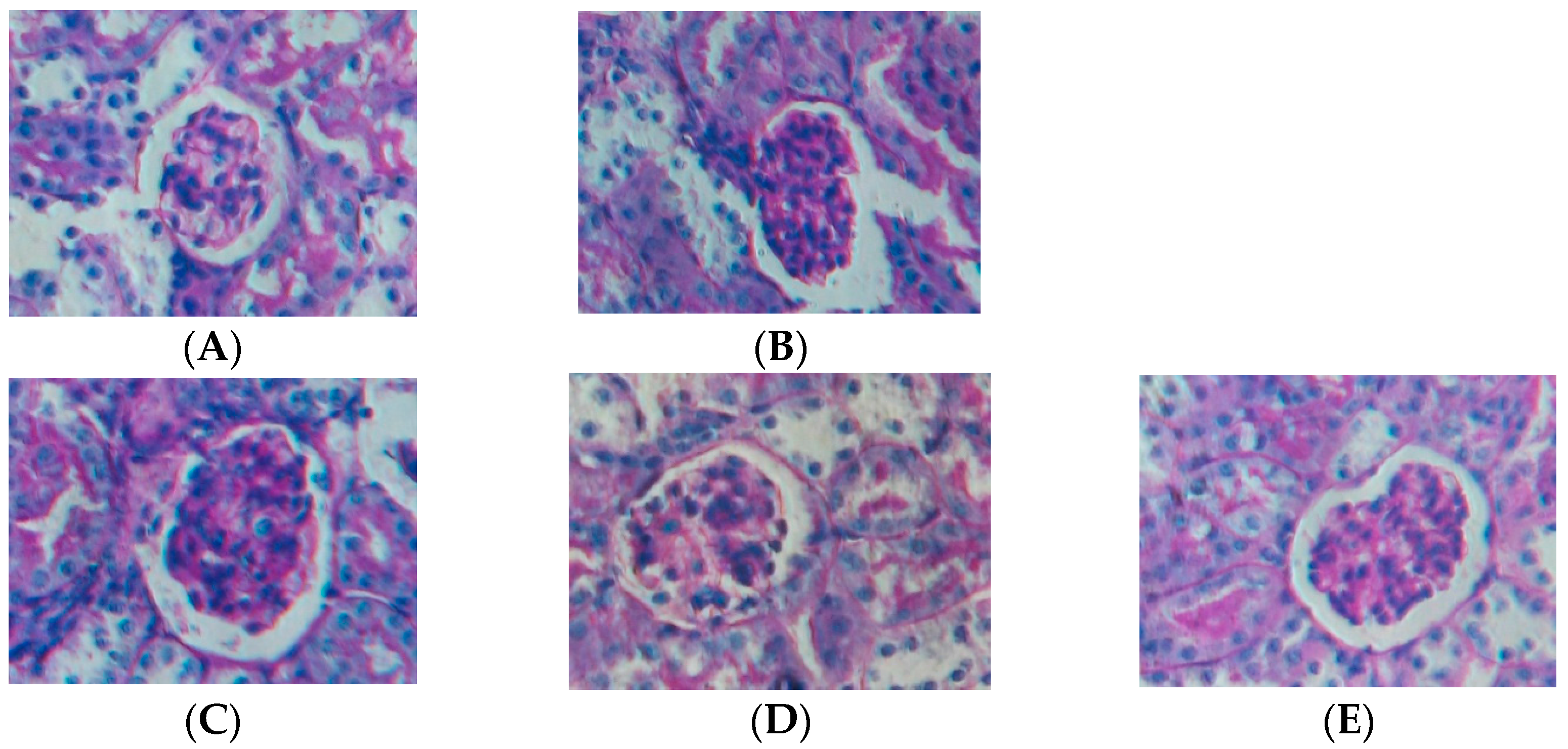
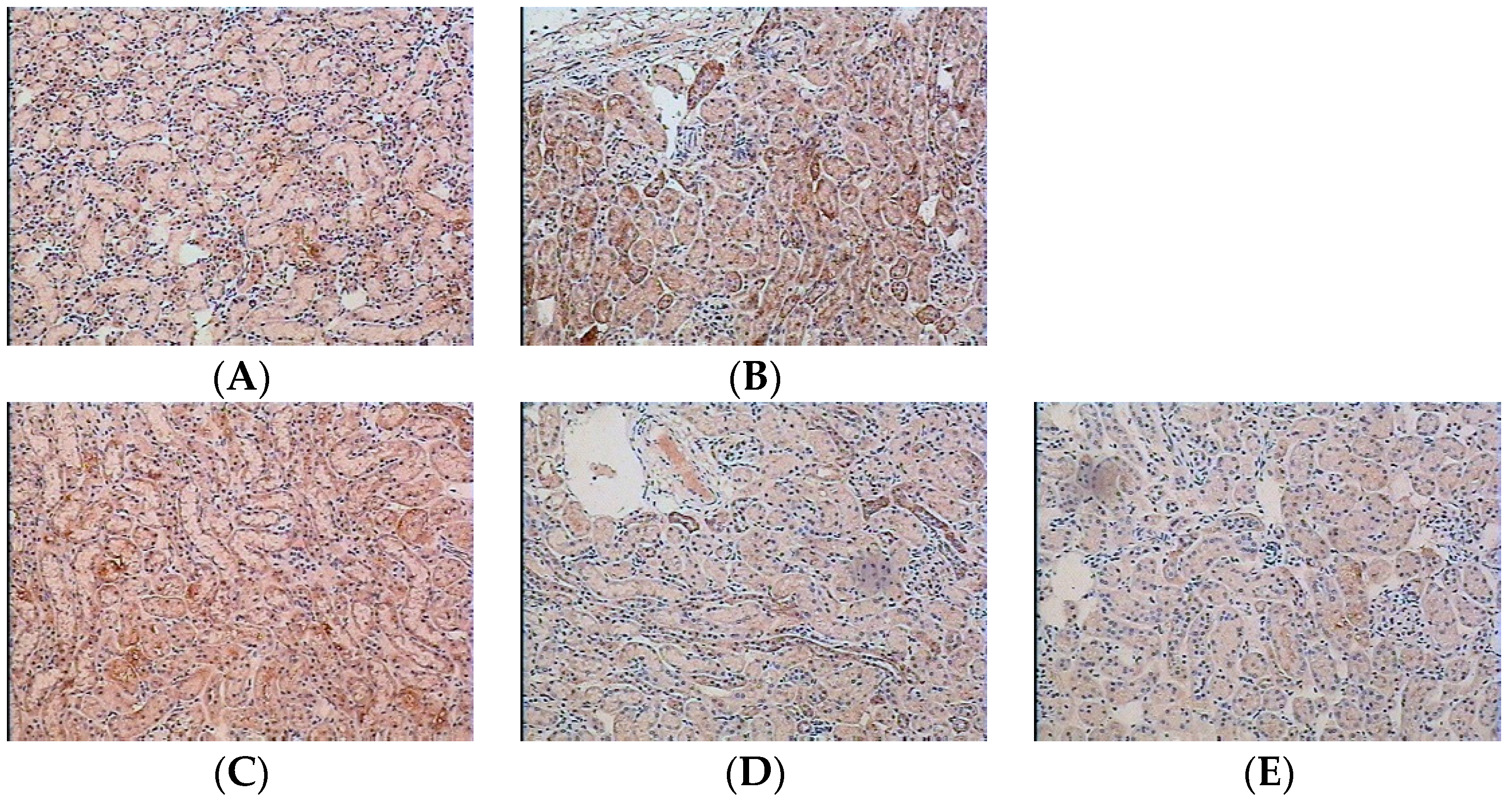
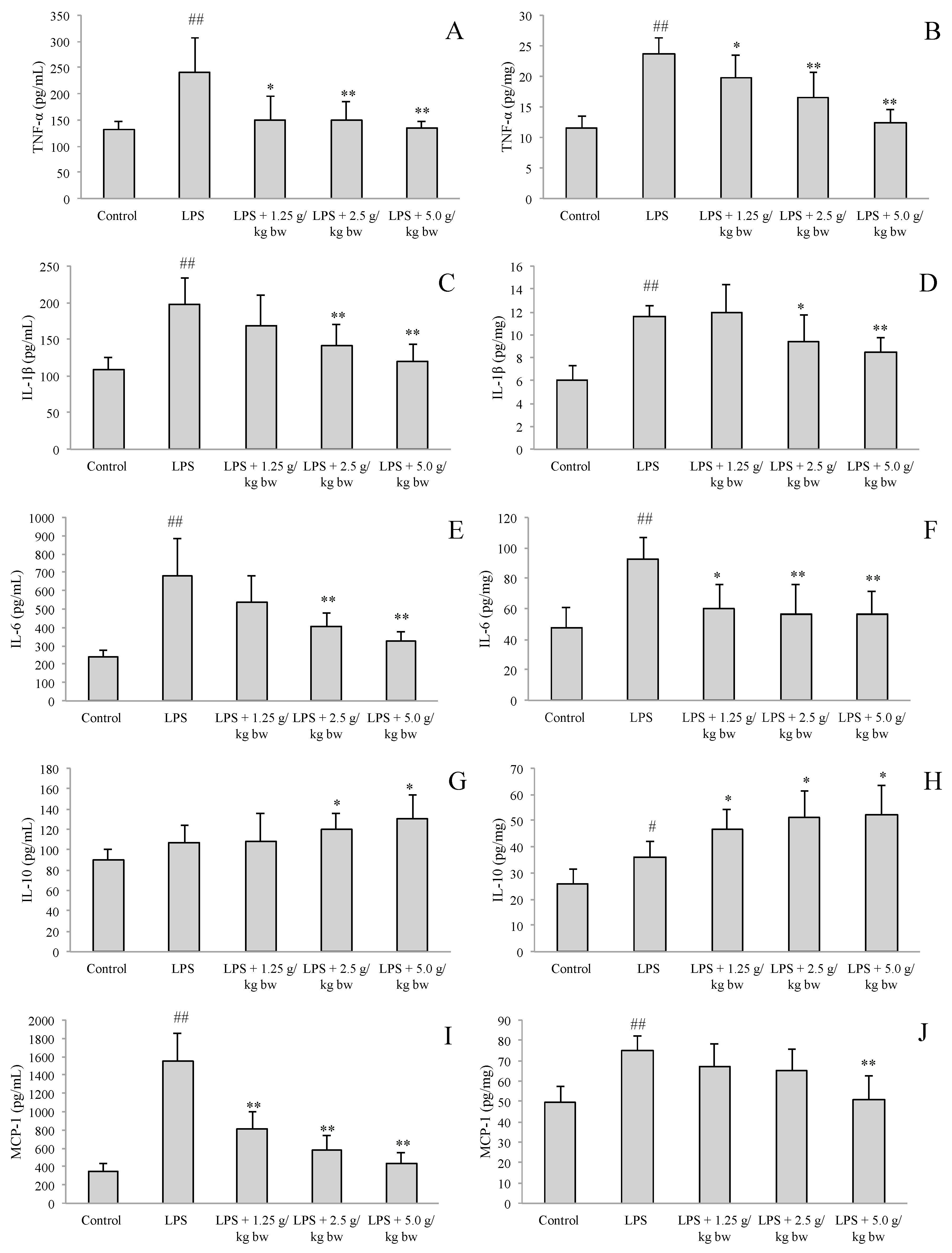
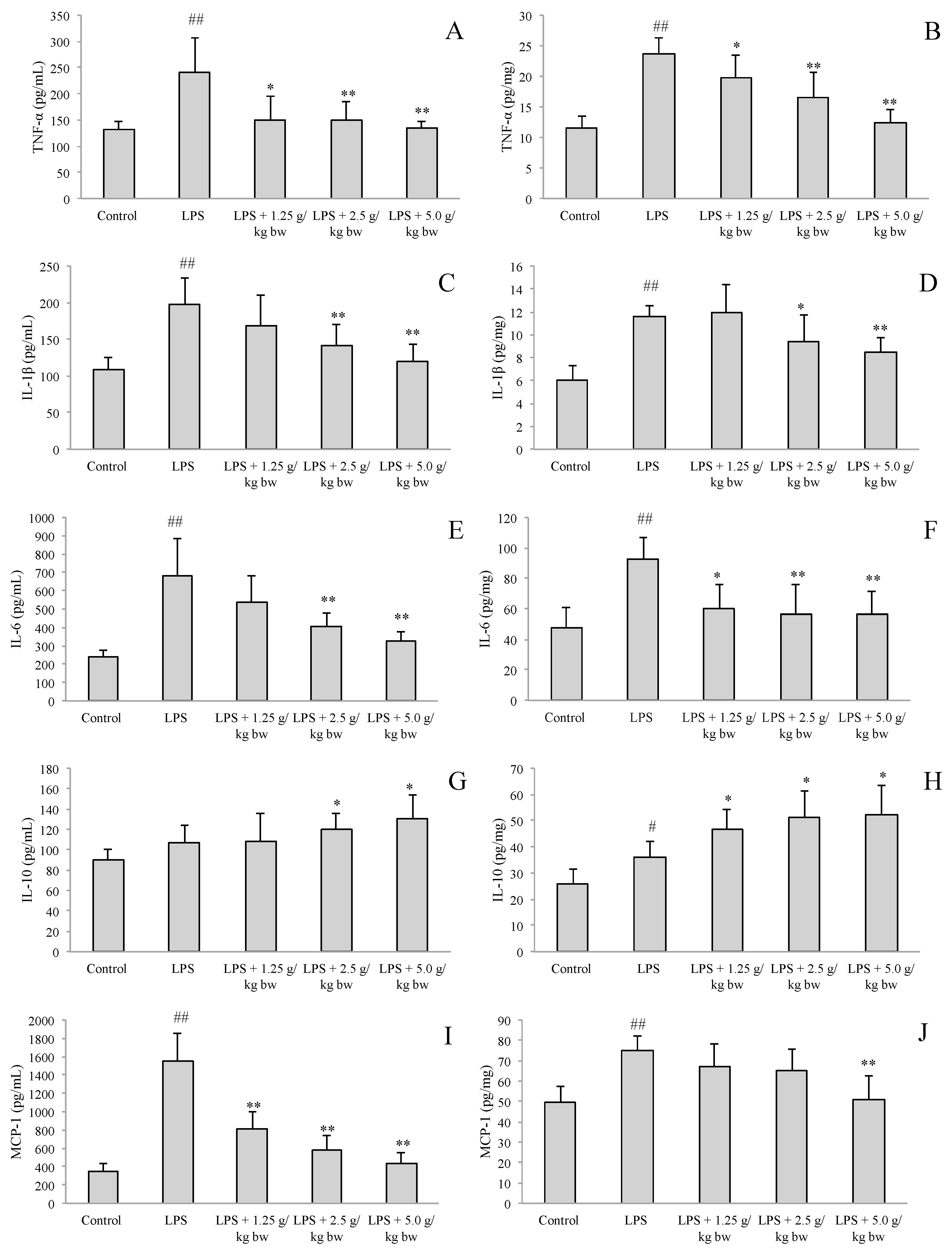
2.2. Effect of Water Extract of H. diffusa on the Productions of Cytokines and Chemokines
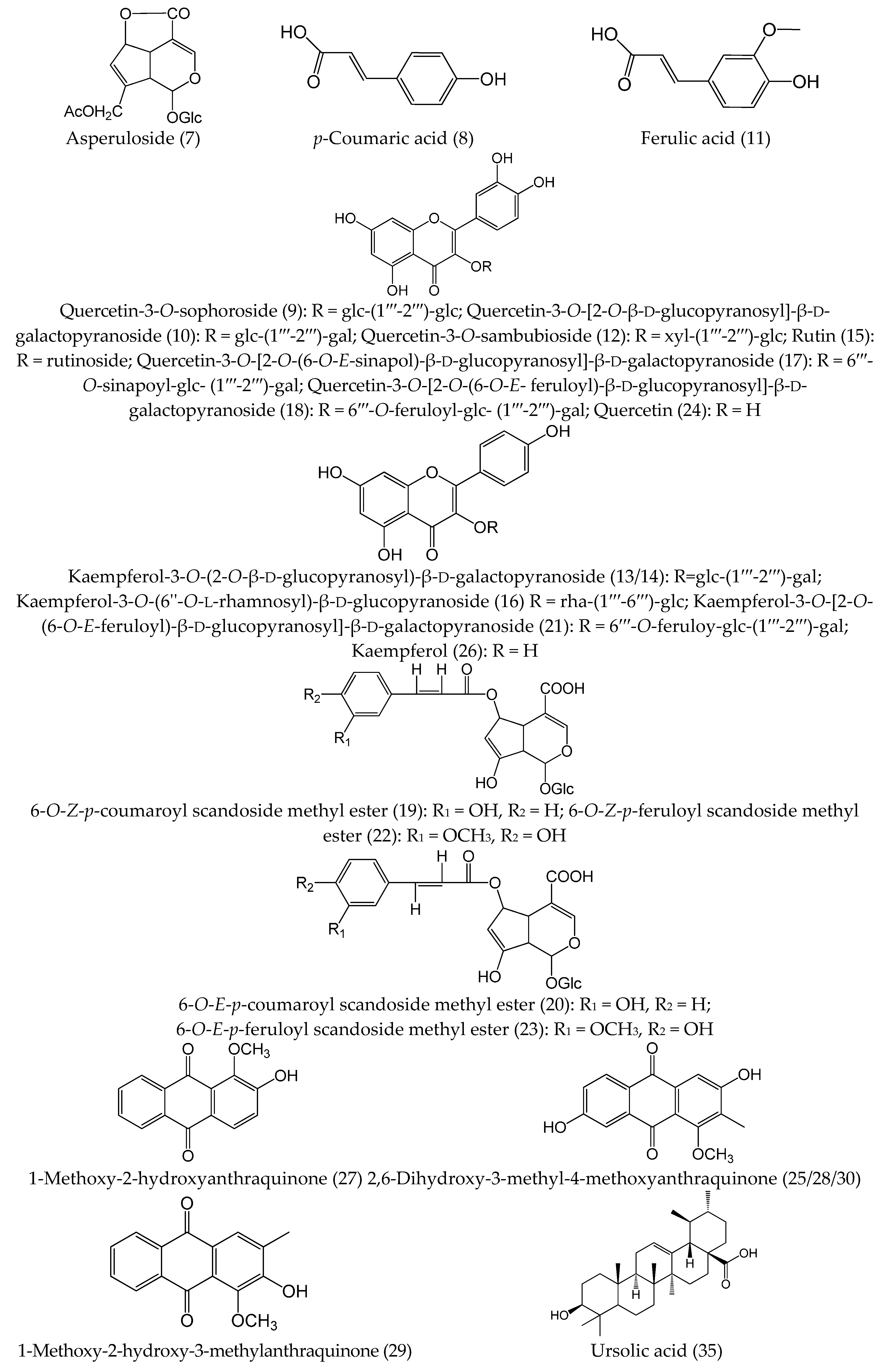
2.3. Chemical Profiles of Water Extract of H. diffusa by UFLC-DAD-Q-TOF-MS/MS
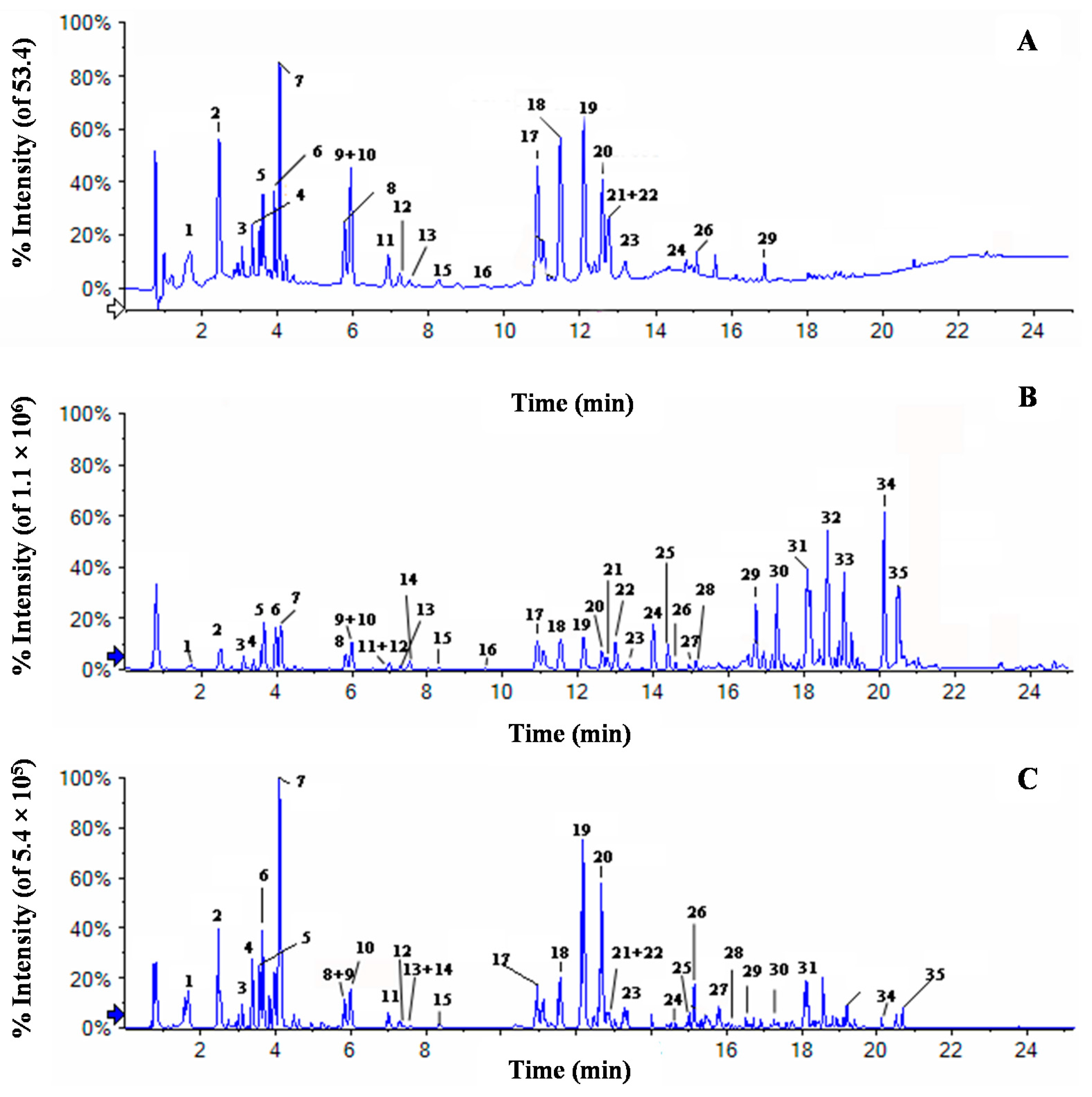
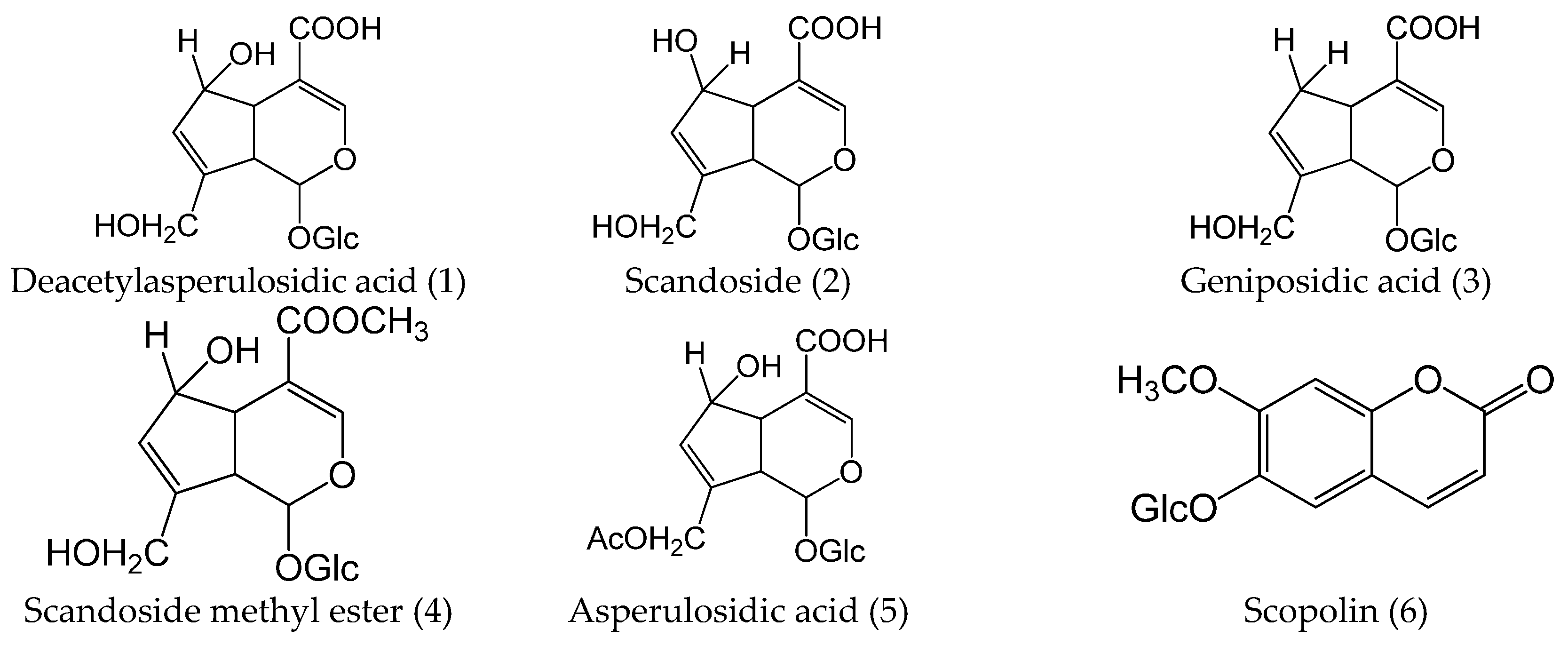

{kind=link}
{kind=link}
{kind=link}
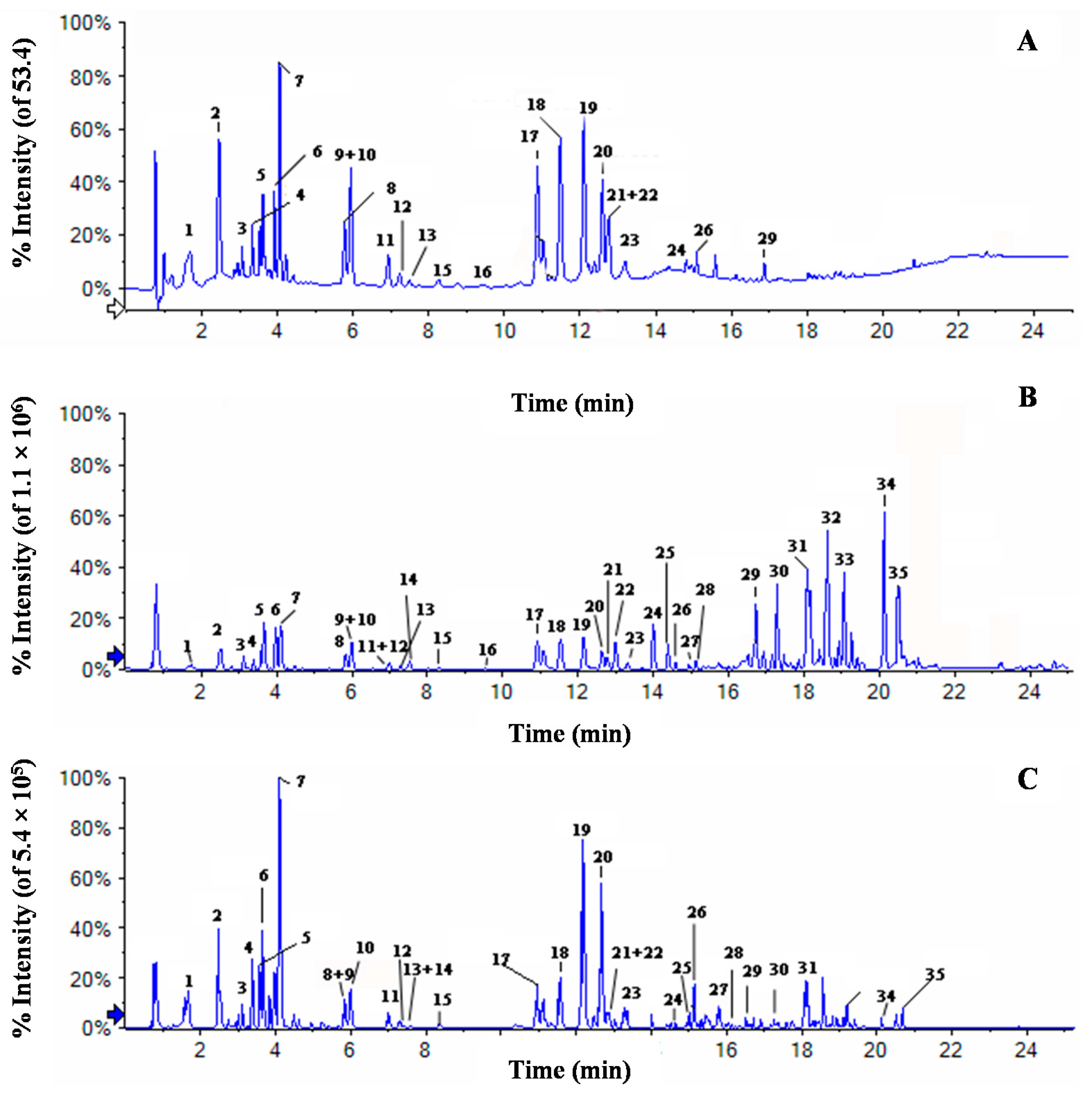{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | Molecular Formula | λmax (nm) | [M + Na]+ | [M + H]+ | [M – H]− | Major Fragmentors in Positive Mode | Major Fragmentors in Negative Mode | Identification | Source |
|---|---|---|---|---|---|---|---|---|---|---|
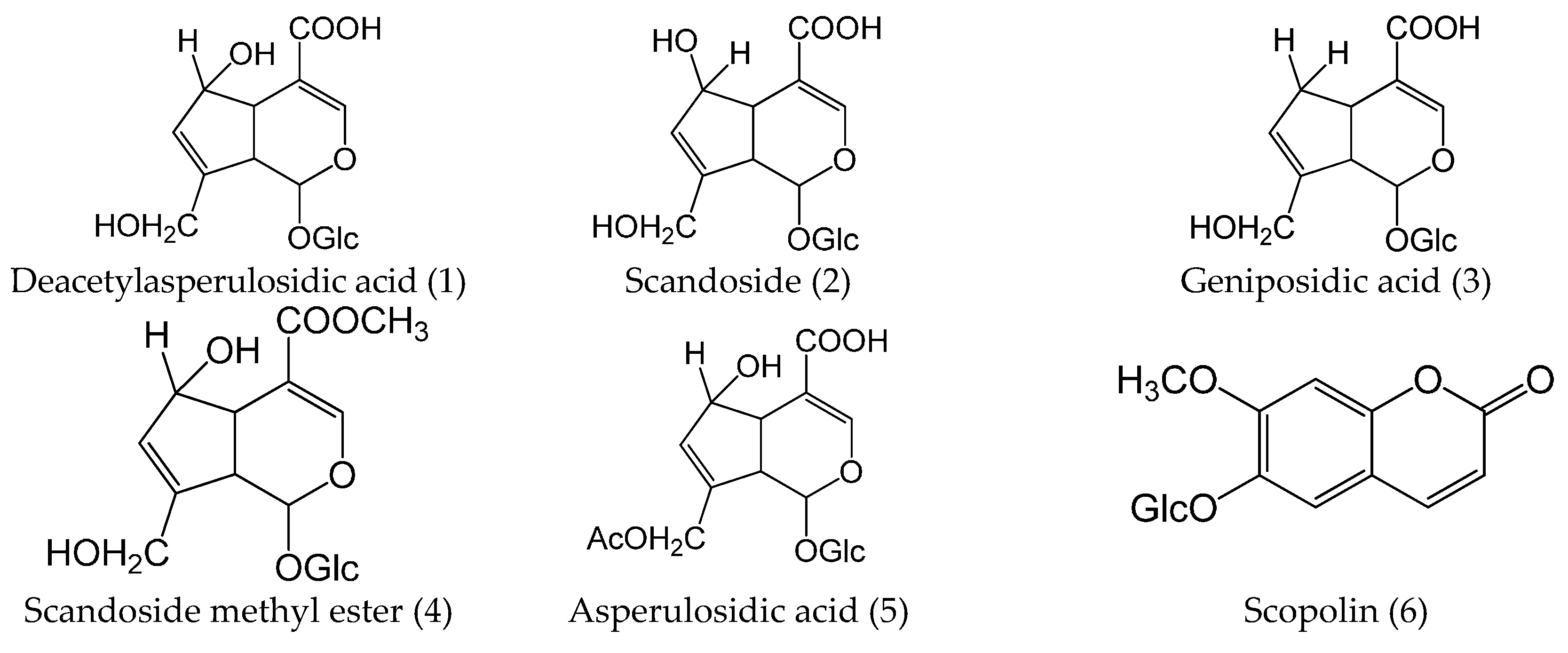
| 1 | 1.62 | C16H22O11 | 239 | 413.10505 (−0.9) | 389.10862(−0.8) | 395.0879 [M+Na–H2O]+, 251.0512 [M+Na–glc]+, 233.0420 [M+H–glc]+, 215.0299 [M+H–glc–H2O]+ | 227.0579 [M–H–glc]−, 209.0468 [M–H–glc–H2O]−, 183.0683 [M–H–glc–CO2]−, 165.0573 [M–H–glc–CO2–H2O]−, 137.0616 [M–H–glc–CO2–H2O–CO]− | Deacetylasperulosidic acid | Herb, Plasma, Kidney | |
| 2 | 2.50 | C16H22O11 | 238 | 413.10505 (−0.9) | 389.10862 (−0.8) | 395.0948 [M+Na–H2O]+, 233.0408 [M+H–glc]+ | 227.0588 [M–H–glc]−, 209.0473 [M–H–glc–H2O]−, 183.0679 [M–H–glc–CO2]−, 165.0573 [M–H–glc–CO2–H2O]−, 147.0464 [M–H–glc–CO2–2H2O]−, 139.0411 [M–H–glc–2CO2]−, 121.0302 [M–H–glc–2CO2–H2O]−, 89.0253, 59.0165 | Scandoside | Herb, Plasma, Kidney | |
| 3 | 3.01 | C16H22O10 | 235 | 397.11053 (+0) | 373.11388 (−0.4) | 235.0561 [M+Na–glc]+, 217.0439, 191.0673 | 211.0620 [M–H–glc]−, 167.0707 [M–H–glc–CO2]−, 149.0606, 123.0453 | Geniposidic acid | Herb, Plasma, Kidney | |
| 4 | 3.38 | C17H24O11 | 238 | 427.12095 (−0.3) | 403.12433 (−0.6) | 265.0669 [M+Na–glc]+ | 241.0710 [M–H–glc]−, 223.0628 [M–H–glc–H2O]−, 209.0444 [M–H–glc–CH3OH]−, 193.0683, 191.0508, 167.0350, 139.0397 | Scandoside methyl ester | Herb, Plasma, Kidney | |
| 5 | 3.60 | C18H24O12 | 229 | 455.11562 (−0.8) | 431.11967 (+0.4) | 437.1027 [M+Na–H2O]+, 293.0621 [M+Na–glc]+, 275.0518 [M+H–glc–H2O]+, 197.0191, 147.0430 | 269.0669 [M–H–glc]−, 251.0588 [M–H–glc–H2O]−, 225.0792 [M–H–glc–CO2]−, 179.0566 [M–H–glc–CO2–HCOOH]−, 165.0573 [M–H–glc–CO2–CH3COOH]−, 121.0302 [M–H–glc–2CO2–CH3COOH]−, 89.0257, 59.0170 | Asperulosidic acid | Herb, Plasma | |
| 6 | 3.98 | C16H18O9 | 221, 320 | 355.10270 (+1.0) | 353.0881 (+0.8) | 163.0384, 145.0238, 135.0442, 117.0343, 89.0398 | 191.0552 [M–H–glc]−, 179.0341, 135.0446 | Scopolin | Herb | |
| 7 | 4.13 | C18H22O11 | 238 | 437.10509 (−0.3) | 413.1085 (−0.9) | 275.0542 [M+Na–glc]+, 187.0372, 147.0458 | 251.0544 [M–H–glc]−, 205.0529 [M–H–glc–HCOOH]−, 191.0364 [M–H–glc–CH3COOH]−, 147.0457 [M–H–glc–CH3COOH–CO2]−, 119.0501 [M–H–glc–CH3COOH–CO–CO2]− | Asperuloside | Herb, Plasma | |
| 8 | 5.79 | C9H8O3 | 219, 296 | 165.05416 (−2.8) | 163.04128 (7.4) | 147.0435 [M+H–H2O]+, 119.049 [M+H–HCOOH]+, 91.0552, 77.0406, 65.0415 | 119.0505 [M–H–CO2]−, 93.0355 | p-Coumaric acid | Herb, Plasma | |
| 9 | 5.85 | C27H30O17 | 256, 355 | 649.1370 (−0.8) | 627.15557 (+) | 625.14120 (+0.3) | 465.1052 [M+H–glc]+, 303.0490 [M+H–2glc]+ | 301.0383 [M–H–2glc]−, 271.0277 [M–H–2glc–CH2O]− | Quercetin-3-O-sophoroside | Herb, Plasma, Kidney |
| 10 | 5.99 | C27H30O17 | 256, 355 | 649.1370 (-0.8) | 627.15525 (−0.5) | 465.1021 [M+H–gal]+, 303.0493 [M+H–gal–glc]+ | 301.0317 [M–H–gal–glc]−, 271.0280 [M–H–gal–glc–HCOH]− | Quercetin-3-O-[2-O-β-d-glucopyranosyl]-β-d-galactopyranoside | Herb, Plasma, Kidney | |
| 11 | 6.92 | C10H10O4 | 219 | 195.06477 (−2.1) | 193.05146 (+4.3) | 177.0549 [M+H–H2O]+, 149.0523 [M+H–HCOOH]+ 145.0283, 89.0396 | 149.0457 [M–H–CO2]− | Ferulic acid | Herb, Plasma | |
| 12 | 7.01 | C26H28O16 | 264, 339 | 619.12649 (-0.8) | 597.14508 (+0.1) | 595.13022 (−0.4) | 465.1011 [M+H–xyl]+, 303.0484 [M+H–xyl–glc]+ | 301.0341 [M–H–xyl–glc]−, 271.0246 [M–H–xyl–glc–CH2O]− | Quercetin-3-O-sambubioside | Herb, Plasma |
| 13 | 7.27 | C27H30O16 | 265, 344 | 633.14211 (−0.8) | 611.16075 (+0.1) | 609.14959 (+5.6) | 449.1062 [M+H–glc]+, 287.0540 [M+H–glc–gal]+, 163.0598 | 285.0348 [M–H–glc–gal]−, 255.0320 [M–H–glc–gal–CH2O]− | Kaempferol-3-O-(2-O-β-d-glucopyranosyl)-β-d-galactopyranoside or isomer | Herb, Plasma, Kidney |
| 14 | 7.57 | C27H30O16 | 264, 340 | 633.14211 (−0.8) | 611.16069 (0) | 609.15134 (+8.6) | 449.1081 [M+H–glc]+, 287.0541 [M+H–glc–gal]+, 145.0482 | 285.0434 [M–H–glc–gal]−, 255.0320 [M–H–glc–gal–CH2O]− | Kaempferol-3-O-(2-O-β-d-glucopyranosyl)-β-d-galactopyranoside or isomer | Herb, Plasma, Kidney |
| 15 | 8.31 | C27H30O16 | 265, 340 | 633.14211 (−0.8) | 611.16069 (0) | 609.15128 (+8.5) | 465.1016 [M+H–rha]+, 303.0489 [M+H–rha–glc]+ | 301.0377 [M–H–rha–glc]− | Rutin | Herb, Plasma |
| 16 | 9.39 | C27H30O15 | 265, 340 | 595.16572 (0) | 593.15110 (-0.2) | 449.1026 [M+H–rha]+, 287.0548 [M+H–rha–glc]+ | 285.0459 [M–H–rha–glc]− | Kaempferol-3-O-(6′′-O-L-rhamnosyl)-β-d-glucopyranoside | Herb, Plasma, Kidney | |
| 17 | 10.92 | C38H40O21 | 265, 338 | 855.19472 (−0.8) | 833.21331 (−0.2) | 831.19906 (+0.2) | 465.1032303.0499 [M+H–sinapol–glc]+, 303.0499 [M+H–sinapol–glc-gal]+, 177.0489 | 625.1414 [M–H–sinapol]−, 301.0344 [M–H–sinapol–glc–gal]−, 271.0250 | Quercetin-3-O-[2-O-(6-O-E-sinapol)-β-d-glucopyranosyl]-β-d-galactopyranoside | Herb, Plasma |
| 18 | 11.54 | C37H38O20 | 254, 336 | 825.18454 (−0.4) | 803.20329 (+0.2) | 801.18894 (+0.7) | 303.0493 [M+H–feruloyl–glc-gal]+, 177.0539 | 625.1477 [M–H–feruloyl]−, 301.0391 [M–H–feruloyl–glc–gal]−, 271.0285 | Quercetin-3-O-[2-O-(6-O-E-feruloyl)-β-d-glucopyranosyl]-β-d-galactopyranoside | Herb, Plasma, Kidney |
| 19 | 12.16 | C26H30O13 | 309 | 573.15785 (+0) | 551.17641 (+0.9) | 549.16274 (+2.5) | 369.1011 [M–H–glc–H2O]−, 223.0634 [M–H–glc–H2O–cou]−, 193.0552, 191.0369, 163.0409, 119.0519 | 6-O-Z-p-coumaroyl scandoside methyl ester | Herb, Plasma | |
| 20 | 12.64 | C26H30O13 | 309 | 573.15785 (+0) | 551.17589 (0) | 549.16396 (+4.7) | 225.0723 [M+H–glc–H2O–cou]+ | 369.0978 [M–H–glc–H2O]−, 223.0616 [M–H–glc–H2O–cou]−, 193.0547, 191.0361, 163.0403, 119.0510 | 6-O-E-p-coumaroyl scandoside methyl ester | Herb, Plasma |
| 21 | 12.78 | C37H38O19 | 268, 328 | 809.18960 (−0.4) | 787.20863 (+0.8) | 785.19407 (+0.8) | 449.1072 [M+H–fer–glc]+, 287.0553 [M+H–fer–glc–gal]+, 177.0542, 145.0275 | 609.1492 [M–H–fer]−, 429.0849 [M–H–fer–glc–H2O]−, 285.0431 [M–H–fer–glc–gal]−, 255.0303, 227.0367 | Kaempferol-3-O-[2-O-(6-O-E-feruloyl)-β-d-glucopyranosyl]-β-d-galactopyranoside | Herb, Plasma |
| 22 | 12.87 | C27H32O14 | 299 | 603.16862 (+0.3) | 581.1864 (−0.2) | 579.17408 (3.7) | 399.1111 [M–H–glc–H2O]−, 223.0617 [M–H–fer–glc–H2O]−, 193.0532, 191.0377, 134.0391, 119.0350 | 6-O-Z-p-feruloyl scandoside methyl ester | Herb | |
| 23 | 13.26 | C27H32O14 | 299 | 603.16862 (+0.3) | 581.18627 (−0.4) | 579.17283 (+1.5) | 399.1083 [M–H–glc–H2O]−, 223.0622 [M–H–fer–glc–H2O]−, 193.0518, 191.0375, 134.0382, 119.0348 | 6-O-E-p-feruloyl scandoside methyl ester | Herb | |
| 24 | 14.64 | C15H10O7 | 254, 370 | 303.04973 (−0.7) | 301.03565 (+0.9) | 285.0385 [M+H–H2O]+, 229.0479 [M+H–H2O–2CO]+, 177.0543, 153.0176 | 286.0583 [M–H–CH3]−, 179.0001, 151.0058 | Quercetin | Herb, Plasma, Kidney | |
| 25 | 14.78 | C16H12O5 | 285.07565 (−0.3) | 283.06143 (+0.8) | 267.0651 [M+H–H2O]+, 239.0722 [M+H–H2O–CO]+, 209.0580 [M+H–H2O–CO–HCOH]+, 181.0650 [M+H–H2O–2CO–HCOH]+, 153.0697 | 268.0399 [M–H–CH3]− , 239.0364 [M–H–CO2]−, 211.0432 [M–H–CO2–CO]−, 195.0471 [M–H–2CO2]− | 2,6-Dihydroxy-3-methyl-4-methoxyanthraquinone or isomer | Herb, Plasma | ||
| 26 | 15.01 | C15H10O6 | 260, 370 | 287.0548 | 285.04049 (+0.1) | 165.0170, 153.0170, 121.0274 | 179.0001 [M–H–2CO2]−, 151.0025, 107.0129, 121.0284 | Kaempferol | Herb, Plasma, Kidney | |
| 27 | 15.98 | C15H10O4 | 255.06504 (−0.6) | 253.05088 (+1.0) | 227.0675 [M+H–CO]+, 209.0570 [M+H–CO–H2O]+, 199.0738 [M+H–2CO]+, 171.0792 [M+H–3CO]+, 153.0678 [M+H–3CO–H2O]+ | 224.0497 [M+H–CHO]−, 183.0670 [M+H–CHO–CH3CO]− | 1-Methoxy-2-hydroxyanthraquinone | Herb, Plasma, Kidney | ||
| 28 | 16.17 | C16H12O5 | 285.07548 (−1.0) | 283.05952 (−5.9) | 270.0544 [M+H–CH3]+, 252.0400 [M+H–CH3–H2O]+, 224.0463 [M+H–CH3–H2O–CO]+ 196.0500 [M+H–CH3–H2O–2CO]+, 168.0568, 139.0521 | 268.0376 [M–H–CH3]−, 253.0146 [M+H–HCOH]−, 240.0432 [M–H–CH3–CO]−, 225.0202 [M+H–HCOH–CO]−, 212.0451 [M–H–CH3–2CO]−, 197.0311 [M+H–HCOH–2CO]−, 184.0539, 169.0316 | 2,6-Dihydroxy-3-methyl-4-methoxyanthraquinone or isomer | Herb, Plasma | ||
| 29 | 16.93 | C16H12O4 | 269.08036 (−1.8) | 267.06684 (+2.1) | 254.0562 [M+H–CH3]+, 226.0610 [M+H–CH3–CO]+, 197.0582 [M+H–CH3–CO–CHO]+, 181.0636, 152.0609 | 252.0534 [M+H–CH3]−, 224.0485 [M+H–CH3–CO]− | 1-Methoxy-2-hydroxy-3-methylanthraquinone | Herb, Plasma | ||
| 30 | 17.53 | C16H12O5 | 285.0754 | 283.06143 (+0.8) | 270.0521 [M+H–CH3]+, 242.0551 [M+H–CH3–CO]+, 214.0610 [M+H–CH3–2CO]+ 187.0594, 169.0646 | 268.0376 [M–H–CH3]−, 240.0432 [M–H–CH3–CO]−, 212.0451 [M–H–CH3–2CO]− | 2,6-Dihydroxy-3-methyl-4-methoxyanthraquinone or isomer | Herb, Plasma | ||
| 31 | 18.21 | C18H28O2 | 277.21599 (−0.8) | 275.20167 (+0.1) | 259.2041, 235.1693, 171.1151, 149.1311, 135.1159, 121.1088, 107.0859, 93.0709, 79.0561 | 231.2161 [M–H–CO2]− | Unidentified | Herb, Plasma, Kidney | ||
| 32 | 18.56 | C18H30O2 | 297.23200 (+0.5) | 277.21874 (+5.2) | 261.2217, 243.2109, 223.1691, 187.1478, 173.1321, 151.1482, 137.1328, 123.1172, 109.1021, 95.0807, 81.0720 | Unidentified | Herb, Plasma, Kidney | |||
| 33 | 19.07 | C19H26O5 | 357.16079 (−0.4) | 335.18563 (+1.0) | 163.0751, 145.0643, 115.0543, 91.0555, 71.0515 | Unidentified | Herb | |||
| 34 | 20.13 | C18H30O2 | 279.23202 (+0.6) | 277.21874 (+5.2) | 261.2214, 243.2105, 223.1696, 187.1479, 173.1322, 151.1470, 137.1321, 123.1173, 109.1020, 95.0867, 81.0716, 67.0572 | 259.2064 [M–H–H2O]−, 233.2282 [M–H–CO2]− | Unidentified | Herb, Plasma, Kidney | ||
| 35 | 20.68 | C30H48O3 | 457.36734 (−0.6) | 455.35279 (−0.6) | 439.3564, 411.3615, 393.3508, 341.2846, 315.2694, 297.2575, 249.1844, 231.2106, 203.1791, 163.1476, 149.1316, 121.1009, 95.0863, 81.0713 | 407.3300 [M–H–HCOOH]− | Ursolic acid | Herb |
2.4. Discussion
3. Experimental Section
3.1. Plant Material
3.2. Chemicals and Reagents
3.3. Sample Preparation
3.4. Animals and Experiment Design
3.5. Preparation of Bioanalytical Samples
3.6. Bioanalytical Method
3.7. Histological Examination
3.8. Immunohistochemical Examination
3.9. Determination of Cytokines
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Editorial Board of Flora of China. The Chinese Academy of Sciences. In Flora of China; Science Press: Beijing, China, 1999; Volume 71, p. 75. [Google Scholar]
- Nanjing University of Chinese Medicine. Dictionary of Chinese Traditional Medicine (Zhong. Yao Da Ci Dian), 2nd ed.; Shanghai Scientific & Technical Publishers: Shanghai, China, 2006; pp. 1039–1041. [Google Scholar]
- Xu, G.H.; Kim, Y.H.; Chi, S.W.; Choo, S.J.; Ryoo, I.J.; Ahn, J.S.; Yoo, I.D. Evaluation of human neutrophil elastase inhibitory effect of iridoid glycosides from Hedyotis diffusa. Bioorg. Med. Chem. Lett. 2010, 20, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Ng, L.T.; Yang, J.J.; Hsu, Y.F. Anti-inflammatory and hepatoprotective activity of Peh-Hue-Juwa-Chi-Cao in male rats. Am. J. Chin. Med. 2002, 30, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Li, D.Y.; Niu, H.Y.; Zhang, Y.; He, P.; Wang, J.H. 2-Hydroxy-3-methylanthraquinone from Hedyotis diffusa Willd induces apoptosis in human leukemic U937 cells through modulation of MAPK pathways. Arch. Pharm. Res. 2013, 36, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, M.; Liu, M.; Li, J.C. Methylanthraquinone from Hedyotis diffusa WILLD induces Ca2+-mediated apoptosis in human breast cancer cells. Toxicol. Vitro 2010, 24, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Meng, Q.X. Chemical and preclinical studies on Hedyotis diffusa with anticancer potential. J. Asian Nat. Prod. Res. 2013, 15, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.H.; Zhou, T.; Li, G.B.; Li, J.; Huang, X.N.; Pan, F.; Gao, N. Characterization and identification of iridoid glucosides, flavonoids and anthraquinones in Hedyotis diffusa by high-performance liquid chromatography/electrospray ionization tandem mass spectrometry. J. Sep. Sci. 2012, 35, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Li, C.M.; Zhao, Y.Y.; Guo, Z.M.; Zhang, X.L.; Xue, X.Y.; Liang, X.M. Effective 2D-RPLC/RPLC enrichment and separation of micro-components from Hedyotis diffusa Willd and characterization by using ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry. J. Pharm. Biomed. Anal. 2014, 99, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Gil-Izquierdo, A.; Andrade, P.B.; Valentão, P.; Tomás-Barberán, F.A. Characterization of C-glycosyl flavones O-glycosylated by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2007, 1161, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Vit, P.; Ferreres, F.; Tomas-Barberan, F. Liquid chromatography-tandem mass spectrometry analysis allows the simultaneous characterization of C-glycosyl and O-glycosyl flavonoids in stingless bee honeys. J. Chromatogr. A 2011, 1218, 7601–7607. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, E.J.; Kim, J.; Kim, Y.; Kim, S.R.; Kim, Y.Y. Neuroprotective constituents from Hedyotis diffusa. J. Nat. Prod. 2001, 64, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.M.; Yang, J.J.; Wang, P.Y.; Lin, C.C. A new acylated flavonol glycoside and antioxidant effects of Hedyotis diffusa. Planta Med. 2000, 66, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Chen, Y.G.; Huang, R. Study on flavonoids from Hedyotis diffusa willia. J. Chin. Med. Mater. 2005, 28, 385–387. [Google Scholar]
- Hamzah, A.S.; Aimi, N.; Lajis, N.H.J. Constituents of Hedyotis herbacea (Rubiaceae). Biochem. Syst. Ecol. 1996, 24, 273. [Google Scholar] [CrossRef]
- Permana, D.; Lajis, N.H.; Othman, A.G.; Ali, A.M.; Aimi, N.; Kitajima, M.; Takayama, H. Anthraquinones from hedyotis herbacea. J. Nat. Prod. 1999, 62, 1430–1431. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.J.; Sung, C.K. Chemical constituents and pharmacological activities of Hedyotis diffusa. Nat. Prod. Sci. 2005, 11, 1–9. [Google Scholar]
- Shi, Y.; Wang, C.H.; Gong, X.G. Apoptosis-inducing effects of two anthraquinones from Hedyotis diffusa WILLD. Biol. Pharm. Bull. 2008, 31, 1075–1078. [Google Scholar] [CrossRef] [PubMed]
- Bascands, J.L.; Bachvarova, M.; Neau, E.; Schanstra, J.P.; Bachvarov, D. Molecular determinants of LPS-induced acute renal inflammation: Implication of the kinin B1 receptor. Biochem. Biophys. Res. Commun. 2009, 386, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.; Kang, K.P.; Moon, S.O.; Sung, M.J.; Kim, D.H.; Kim, H.J.; Park, S.K. Agonist of peroxisome proliferator-activated receptor-γ, rosiglitazone, reduces renal injury and dysfunction in a murine sepsis model. Nephrol. Dial. Transplant. 2005, 20, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Hsiang, C.Y.; Cheng, H.M.; Lo, H.Y.; Li, C.C.; Chou, P.C.; Lee, Y.C.; Ho, T.Y. Ginger and zingerone ameliorate lipopolysaccharide-induced acute systemic inflammation in mice, assessed by nuclear factor-κB bioluminescent imaging. J. Agric. Food Chem. 2015, 63, 6051–6058. [Google Scholar] [CrossRef] [PubMed]
- Granholm, N.A.; Cavallo, T. Long-lasting effects of bacterial lipopolysaccharide promote progression of lupus nephritis in NZB/W mice. Lupus 1994, 3, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.N.; Cui, Y.L.; Wang, Q.S.; Wang, S.X. Amelioration of Danhong iniection on the lipopolysaccharide-stimulated systemic acute inflammatory reaction via multi-target strategy. J. Ethnopharmacol. 2013, 149, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.R.M.; Silva, M.D.; Cordova, M.M.; Schambach, T.R.; Pizzolatti, M.G.; Santos, A.R.S. Anti-inflammatory action of hydroalcoholic extract, dichloromethane fraction and steroid α-spinasterol from Polygala sabulosa in LPS-induced peritonitis in mice. J. Ethnopharmacol. 2014, 151, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Segerer, S.; Nelson, P.J.; Schlondorff, D. Chemokines, chemokine receptors, and renal disease: From basic science to pathophysiologic and therapeutic studies. J. Am. Soc. Nephrol. 2000, 11, 152–176. [Google Scholar] [PubMed]
- Abraham, A.P.; Ma, F.Y.; Mulley, W.R.; Ozols, E.; Nikolic-Paterson, D.J. Macrophage infiltration and renal damage are independent of matrix metalloproteinase 12 in the obstructed kidney. Nephrology 2012, 17, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheng, Z.A.; Wang, Y.M.; Dai, X.Q.; Zhang, J.; Xue, D.Y. Paeoniflorin exerts a nephroprotective effect on concanavalin A-induced damage through inhibition of macrophage infiltration. Diagn. Pathol. 2015, 10, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.H.; Dudics, S.; Acharya, B.; Moudgil, K.D. Cytokine-Modulating strategies and newer cytokine targets for arthritis therapy. Int. J. Mol. Sci. 2015, 16, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, P.N.; Dyanov, H.M.; Park, P.; Wang, J.; Newell, K.A.; Quigg, R.J. Acute renal failure in endotoxemia is caused by TNF acting directly on TNF receptor-1 in kidney. J. Immunol. 2002, 168, 5817–5823. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, G.; Reeves, W.B. TNF-α mediates chemokine and cytokine expression and renal injury in cisplatin nephrotoxicity. J. Clin. Investig. 2002, 110, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Hanada, T. Regulation of cytokine signaling and inflammation. Cytokine Growth Factor Rev. 2002, 13, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.S.; Chatterjee, P.K.; Di Paola, R.; Mazzon, E.; de Sarro, A.; Cuzzpcrea, S.; Thiemermann, C. Endogenous interleukin-6 enhances the renal injury, dysfunction, and inflammation caused by ischemia/reperfusion. J. Pharmacol. Exp. Ther. 2005, 312, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Rovin, B.H.; Phan, L.T. Chemotactic factors and renal inflammation. Am. J. Kidney Dis. 1998, 31, 1065–1084. [Google Scholar] [CrossRef] [PubMed]
- Banas, B.; Luckow, B.; Moller, M.; Klier, C.; Nelson, P.J.; Schadde, E.; Brigl, M.; Halevy, D.; Holthofer, H.; Reinhart, B.; et al. Chemokine and chemokine receptor expression in a novel human mesangial cell line. J. Am. Soc. Nephrol. 1999, 10, 2314–2322. [Google Scholar] [PubMed]
- Moore, K.W.; De Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K.; Ishii, R.; Kobiyama, K.; Kitanaka, S. New benzophenone and quercetin galloyl glycosides from Psidium guajava L. J. Nat. Med. 2010, 64, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Flamini, R.; Mattivi, F.; de Rosso, M.; Arapitsas, P.; Bavaresco, L. Advanced knowledge of three important classes of grape phenolics: Anthocyanins, stilbenes and flavonols. Int. J. Mol. Sci. 2013, 14, 19651–19669. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.Q.; Yin, G.J.; Fan, Y.T.; Qiu, L.; Cang, X.F.; Yu, G.; Hu, Y.L.; Xing, M.; Wu, D.Q.; Wang, X.P.; et al. Catalpol ameliorates sodium taurocholate-induced acute pancreatitis in rats via inhibiting activation of nuclear factor κB. Int. J. Mol. Sci. 2014, 15, 11957–11972. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Zhu, T.; Zhang, W.; Wang, T.; Shen, Y.C.; Wan, Q.F.; Wen, F.Q. Emodin ameliorates LPS-induced acute lung injury, involving the inactivation of NF-κB in mice. Int. J. Mol. Sci. 2014, 15, 19355–19368. [Google Scholar] [CrossRef] [PubMed]
- He, Y.M.; Zhu, S.; Ge, Y.W.; Kazuma, K.; Zou, K.; Cai, S.Q.; Komatsu, K. The anti-inflammatory secoiridoid glycosides from Gentianae Scabrae Radix: The root and rhizome of Gentiana. scabra. J. Nat. Med. 2015, 69, 303–312. [Google Scholar] [CrossRef] [PubMed]
- He, Y.M.; Zhu, S.; Ge, Y.W.; Cai, S.Q.; Komatsu, K. Secoiridoid glycosides from the root of Gentiana crassicaulis with inhibitory effects against LPS-induced NO and IL-6 production in RAW264 macrophages. J. Nat. Med. 2015, 69, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Lee, E.K.; Heo, H.S.; Sung, B.; Kim, M.K.; Lee, J.; Kim, N.D.; Anton, S.; Choi, J.S.; Yu, B.P.; et al. The anti-inflammatory effect of kaempferol in aged kidney tissues: The involvement of nuclear factor-κB via nuclear factor-inducing kinase/IκB kinase and mitogen-activated protein kinase pathways. J. Med. Food 2009, 12, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Sugiyama, H.; Stylianou, E.; Kitamura, M. Bioflavonoid quercetin inhibits interleukin-1-induced transcriptional expression of monocyte chemoattractant protein-1 in glomerular cells via suppression of nuclear factor-κB. J. Am. Soc. Nephorl. 1999, 10, 2290–2296. [Google Scholar]
- Arjumand, W.; Seth, A.; Sultana, S. Rutin attenuates cisplatin induced renal inflammation and apoptosis by reducing NF-κB, TNF-α and caspase-3 expression in wistar rats. Food Chem. Toxicol. 2011, 49, 2013–2021. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.Y.; Chung, K.S.; Jeon, E.; Nugroho, A.; Park, H.J.; An, H.J. Anti-inflammatory activity of saxifragin via inhibition of NF-κB involves caspase-1 activation. J. Nat. Prod. 2015, 78, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.T.; Zhou, E.S.; Wei, D.; Li, D.P.; Wei, Z.K.; Zhang, W.; Zhang, X.C. Emodin inhibits LPS-induced inflammatory response by activating PPAR-γ in mouse mammary epithelial cells. Int. Immunopharmacol. 2014, 21, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiong, W.J.; Yang, J.; Zhong, J.; Zhang, L.; Zheng, J.; Liu, H.; Zhang, Q.; Ouyang, X.Q.; Lei, L.; Yu, X. Attenuation of inflammation by emodin in lipopolysaccharide-induced acute kidney injury via inhibition of toll-like receptor 2 signal pathway. Iran. J. Kidney Dis. 2015, 9, 202–208. [Google Scholar] [PubMed]
- Zhang, F.; Wu, Z.J.; Sun, L.N.; Wang, J.; Tao, X.; Chen, W.S. Iridoid glucosides and a C13-norisoprenoid from Lamiophlomis. rotata and their effects on NF-κB activation. Bioorg. Med. Chem. Lett. 2012, 22, 4447–4452. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Pang, M.; Dong, L.; Huang, X.; Wang, S.; Zhou, L. Anti-inflammatory and immunomodulatory effects of iridoid glycosides from Paederia scandens (LOUR.) MERRILL (Rubiaceae) on uric acid nephropathy rats. Life Sci. 2012, 91, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.X.; Zhu, W.J.; Pang, M.Q.; Jeffry, J.; Zhou, L.L. Protective effect of iridoid glycosides from Paederia scandens (LOUR.) MERRILL (Rubiaceae) on uric acid nephropathy rats induced by yeast and potassium oxonate. Food Chem. Toxicol. 2014, 64, 57–64. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, J.-H.; Liu, M.-H.; Zhang, X.-L.; He, J.-Y. Chemical Profiles and Protective Effect of Hedyotis diffusa Willd in Lipopolysaccharide-Induced Renal Inflammation Mice. Int. J. Mol. Sci. 2015, 16, 27252-27269. https://doi.org/10.3390/ijms161126021
Ye J-H, Liu M-H, Zhang X-L, He J-Y. Chemical Profiles and Protective Effect of Hedyotis diffusa Willd in Lipopolysaccharide-Induced Renal Inflammation Mice. International Journal of Molecular Sciences. 2015; 16(11):27252-27269. https://doi.org/10.3390/ijms161126021
Chicago/Turabian StyleYe, Jian-Hong, Meng-Hua Liu, Xu-Lin Zhang, and Jing-Yu He. 2015. "Chemical Profiles and Protective Effect of Hedyotis diffusa Willd in Lipopolysaccharide-Induced Renal Inflammation Mice" International Journal of Molecular Sciences 16, no. 11: 27252-27269. https://doi.org/10.3390/ijms161126021
APA StyleYe, J.-H., Liu, M.-H., Zhang, X.-L., & He, J.-Y. (2015). Chemical Profiles and Protective Effect of Hedyotis diffusa Willd in Lipopolysaccharide-Induced Renal Inflammation Mice. International Journal of Molecular Sciences, 16(11), 27252-27269. https://doi.org/10.3390/ijms161126021