
MicroRNA-214 and MicroRNA-126 Are Potential Biomarkers for Malignant Endothelial Proliferative Diseases

 ,
, 
Abstract
:1. Introduction
2. Results
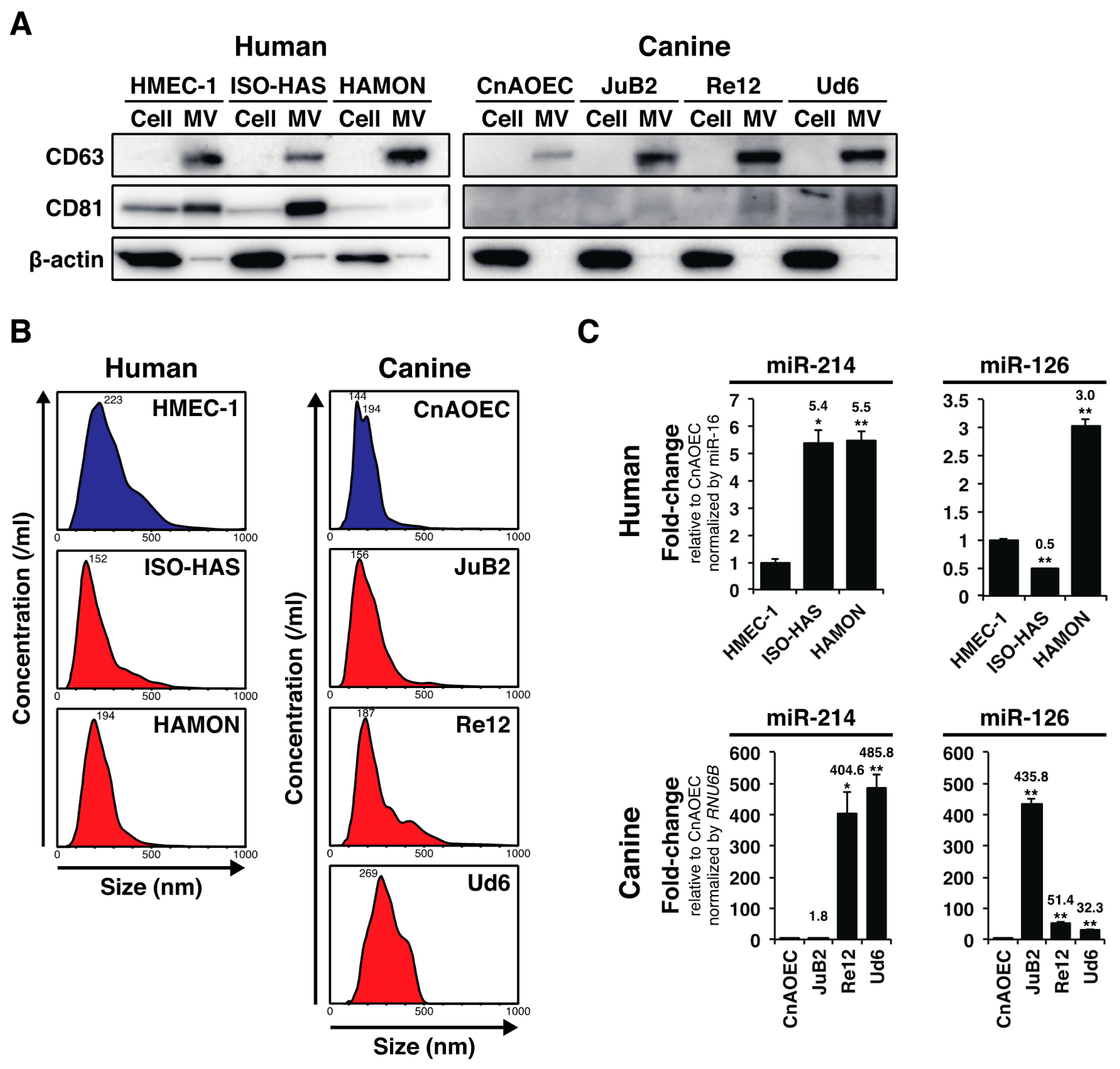
2.1. AS and HSA Cell Lines Over-Secreted miR-214 and miR-126 via MVs
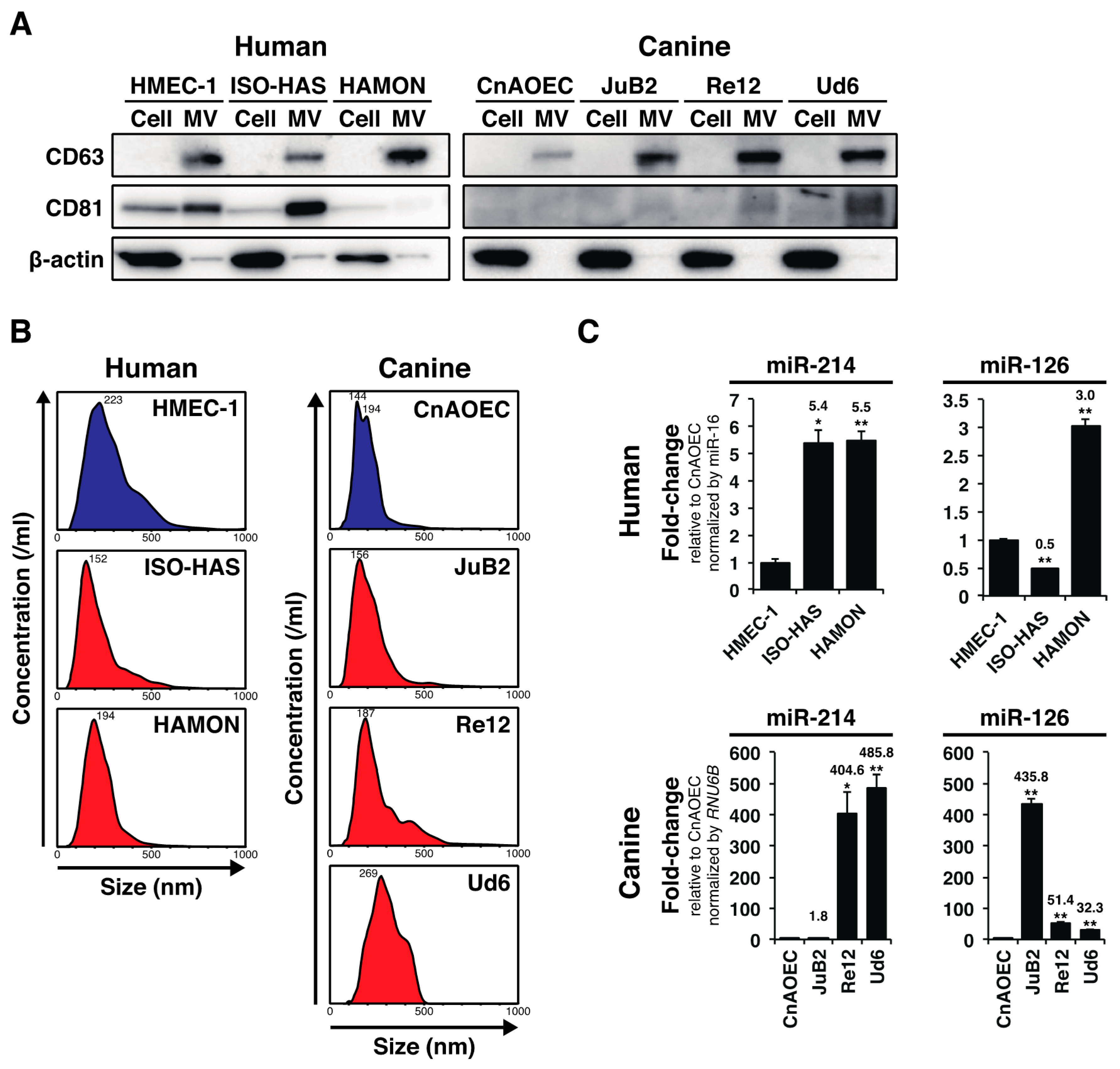
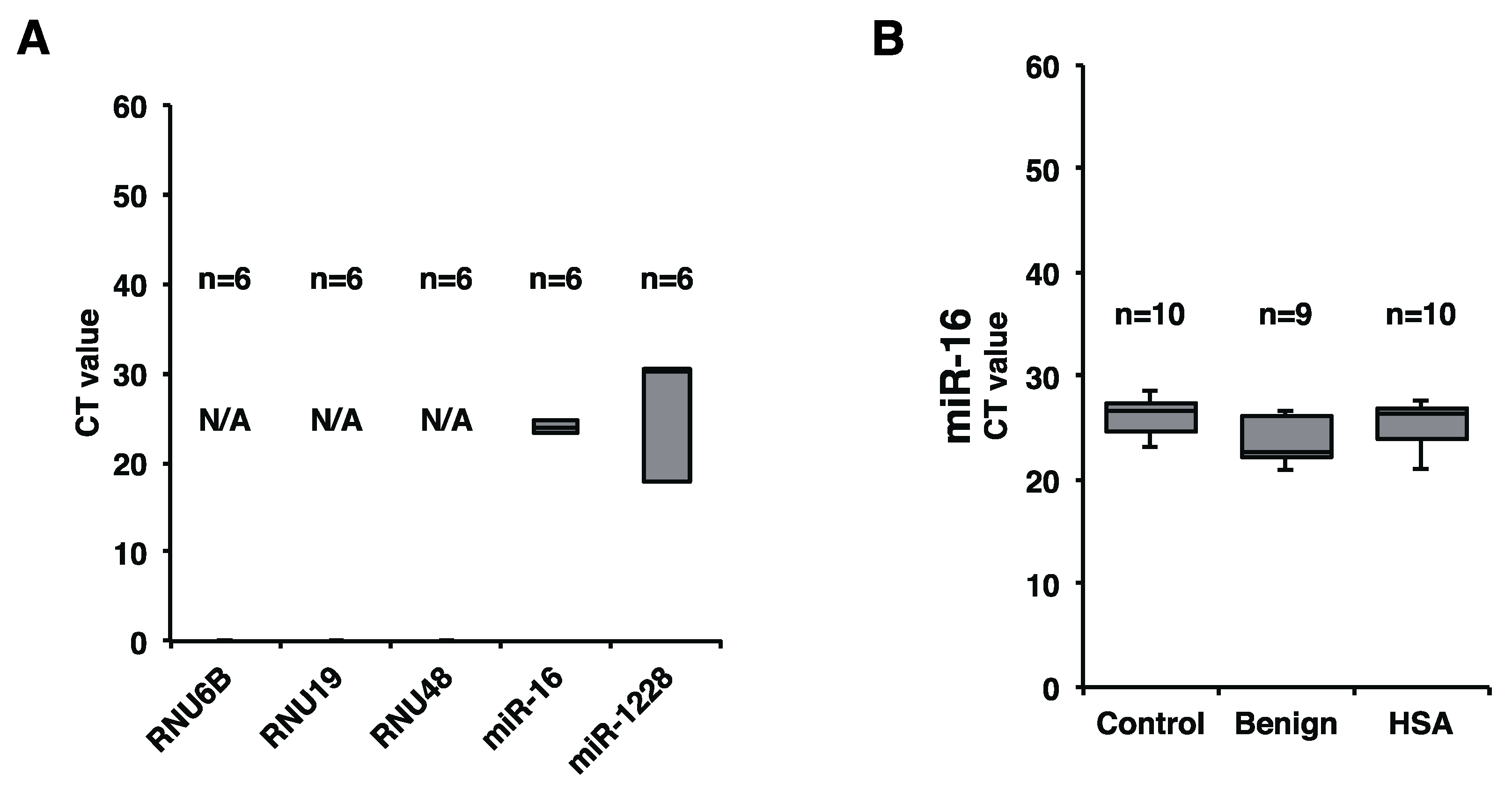
2.2. Plasma miR-16 Was a Suitable Internal Control in this Study
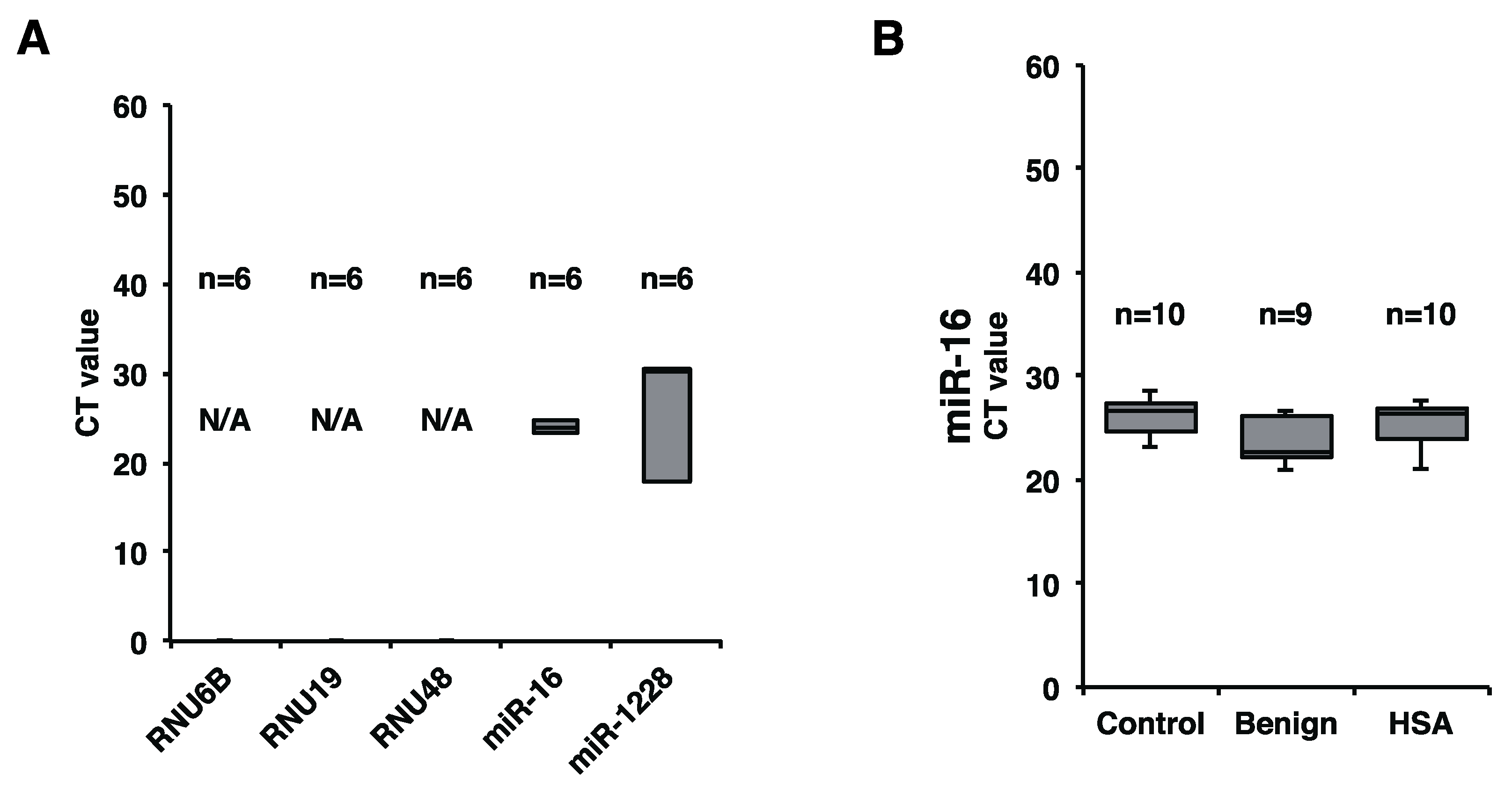
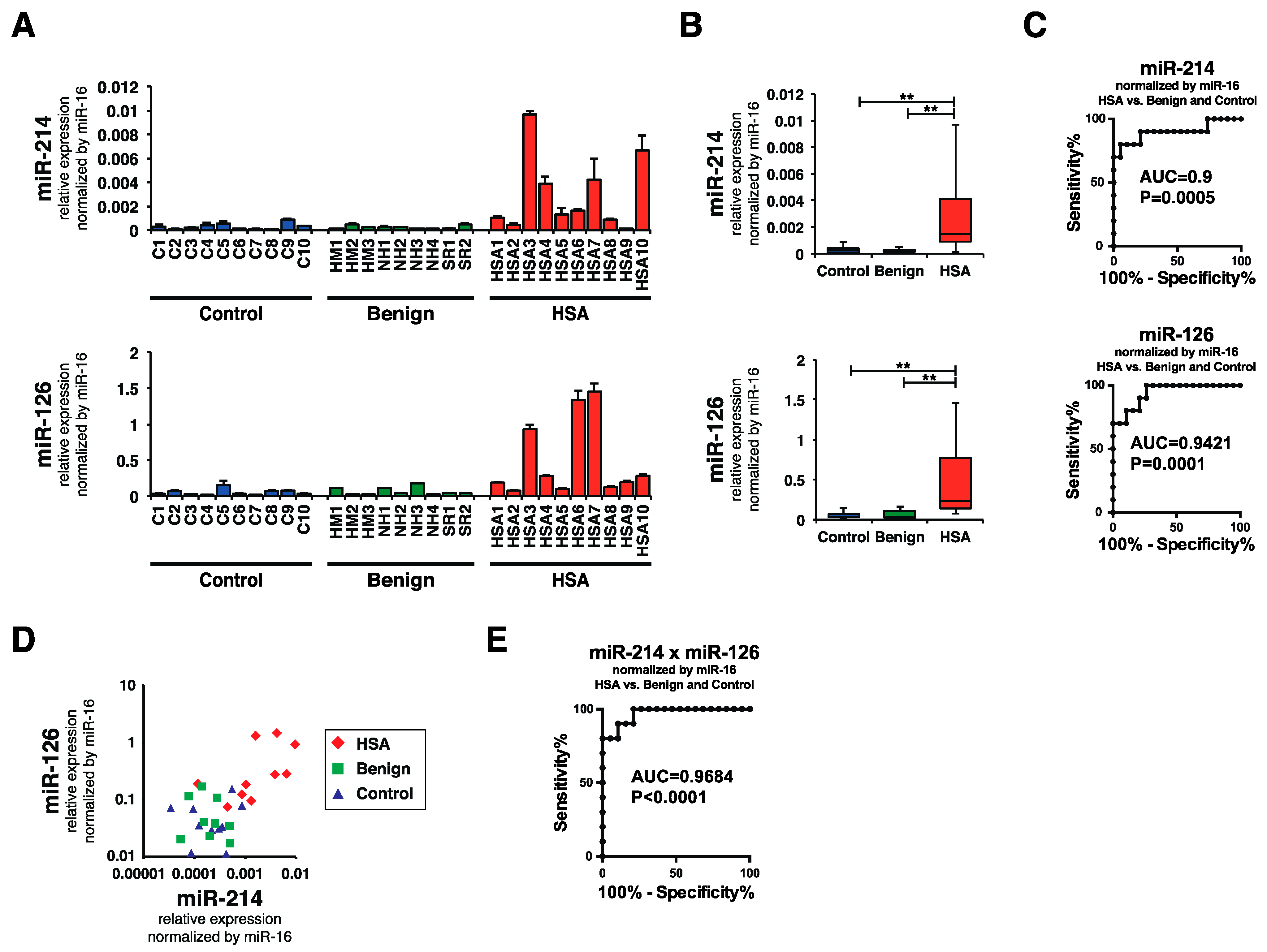
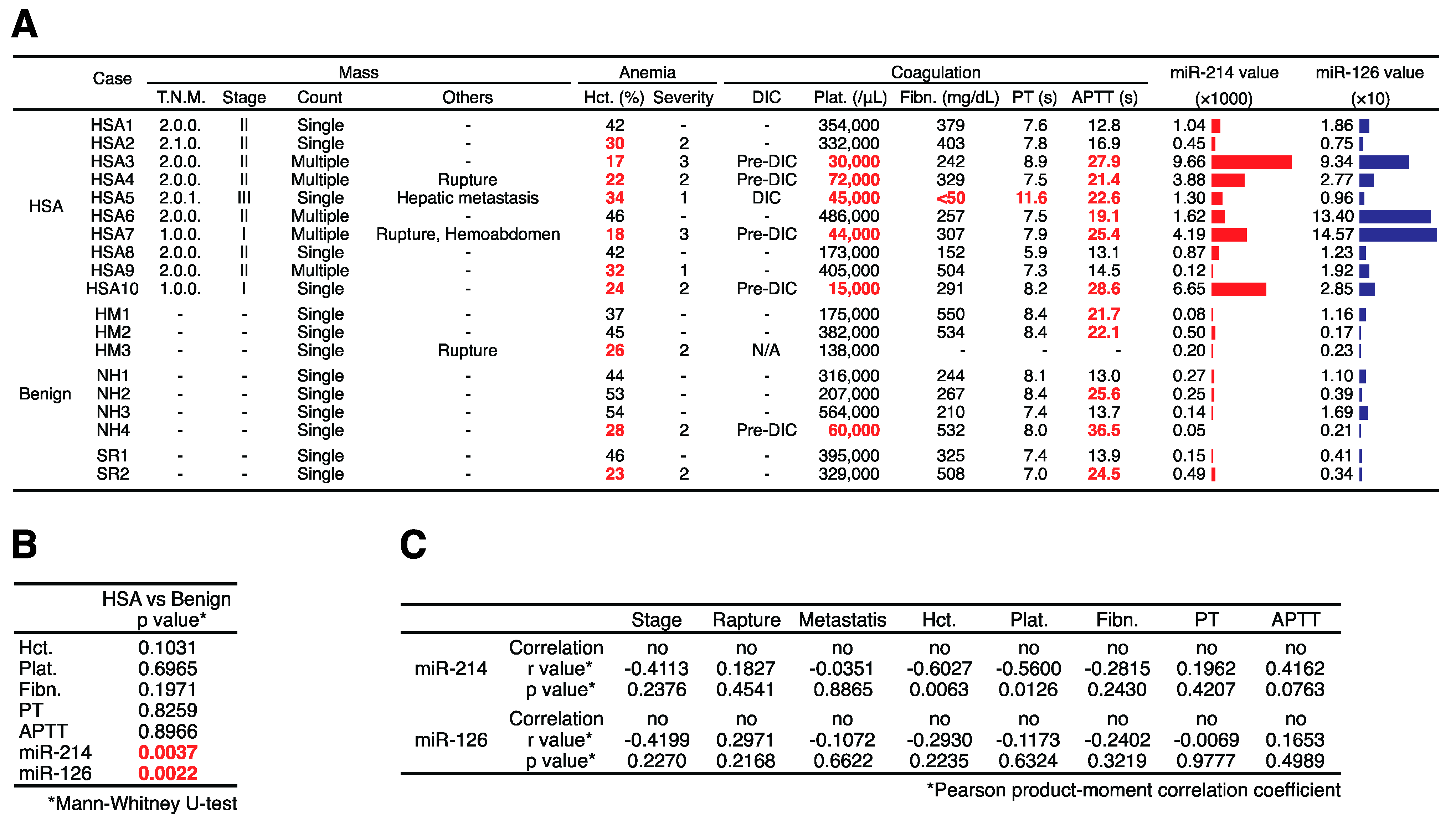
2.3. Plasma miR-214 and miR-126 Levels Were Significantly Increased in the Plasma of Canines with HSA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (y.m.) | Weight (kg) | p Value * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Median | s.d. | Mean | Median | s.d. | Age | Weight | ||||
| HSA | 10.6. | 10.9. | 2.8. | 11.1 | 6.6 | 9.6 | HSA vs. Benign | >0.05 | >0.05 | ||
| Benign | 10.1. | 11.1. | 2.6. | 10.2 | 6.8 | 9.6 | Benign vs. Control | >0.05 | >0.05 | ||
| Control | 10.3. | 10.3. | 2.8. | 6.9 | 5.4 | 3.3 | HSA vs. Control | >0.05 | >0.05 | ||
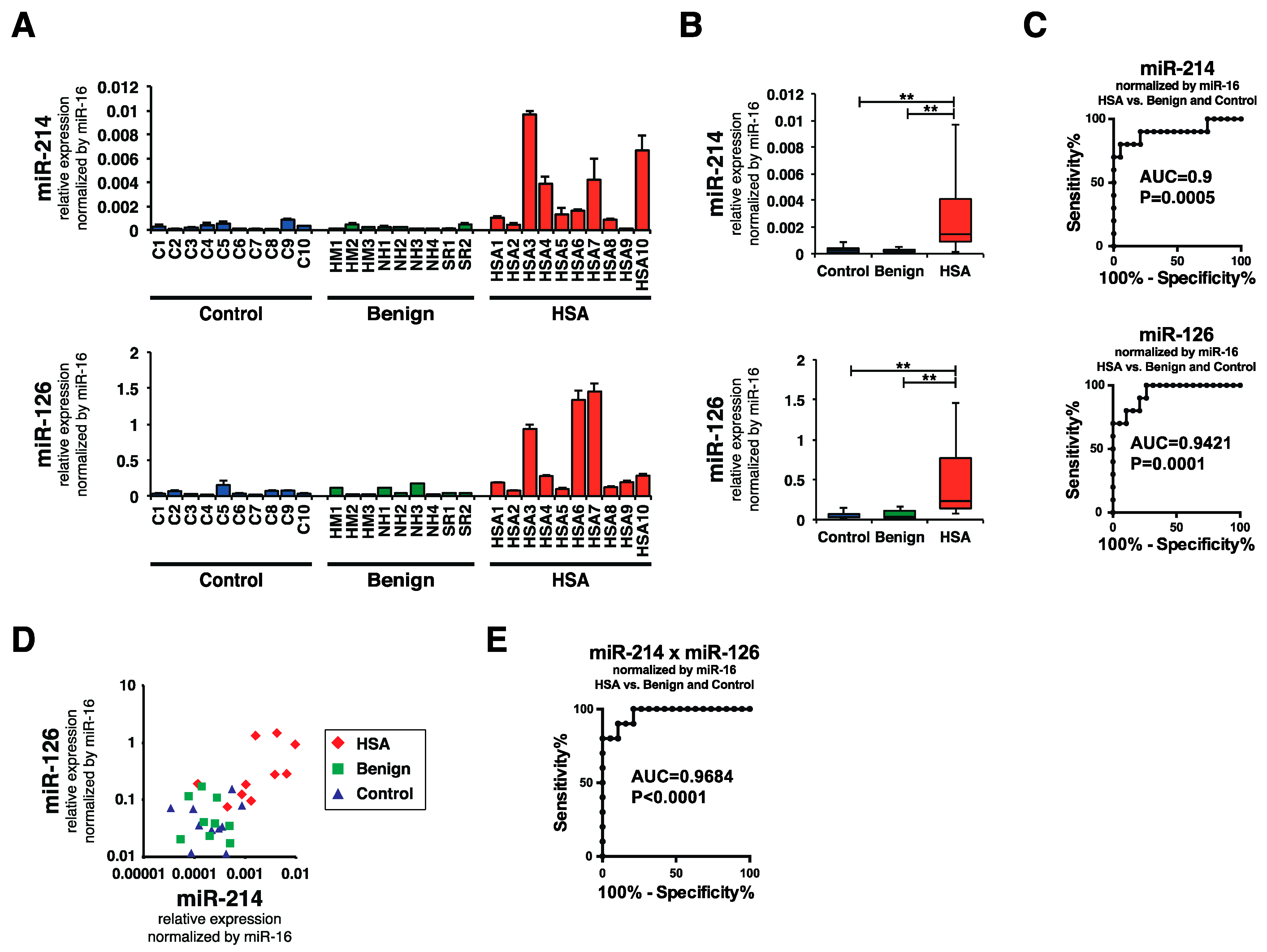
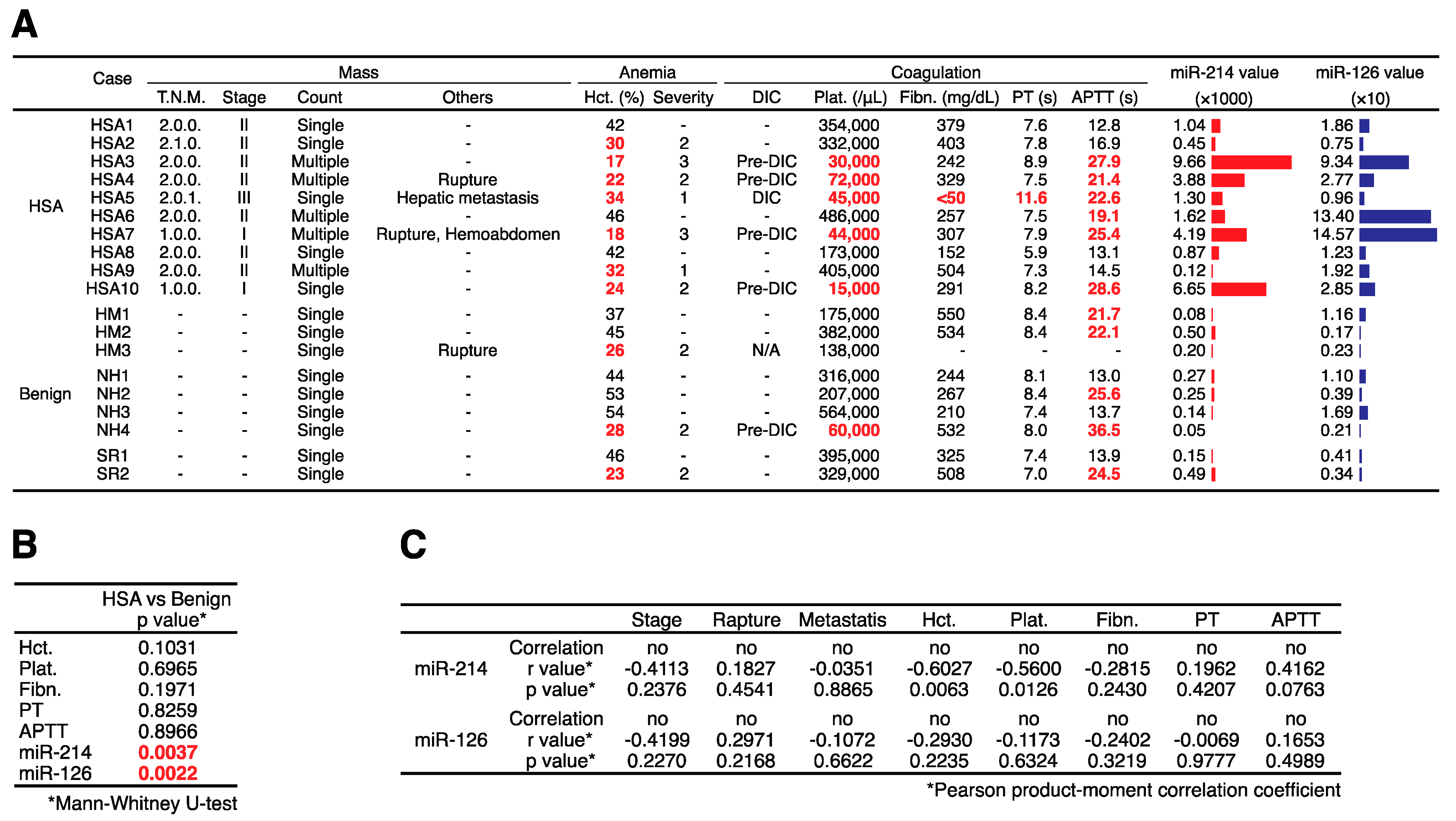
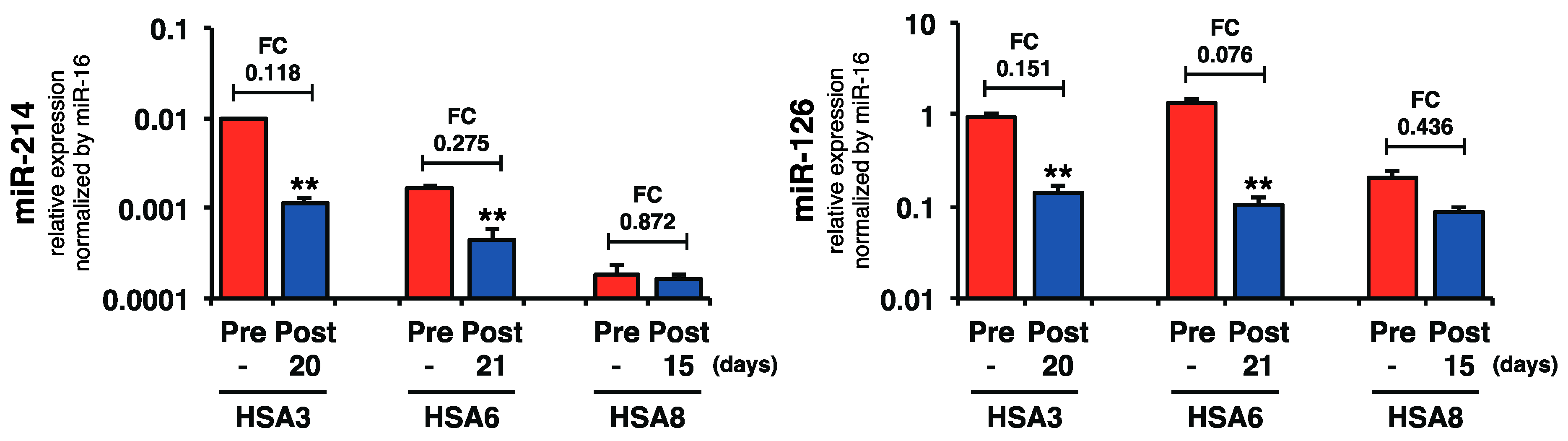
2.4. Plasma miR-214 and miR-126 Levels Were Decreased after the Surgical Resection of Primary Splenic HSA
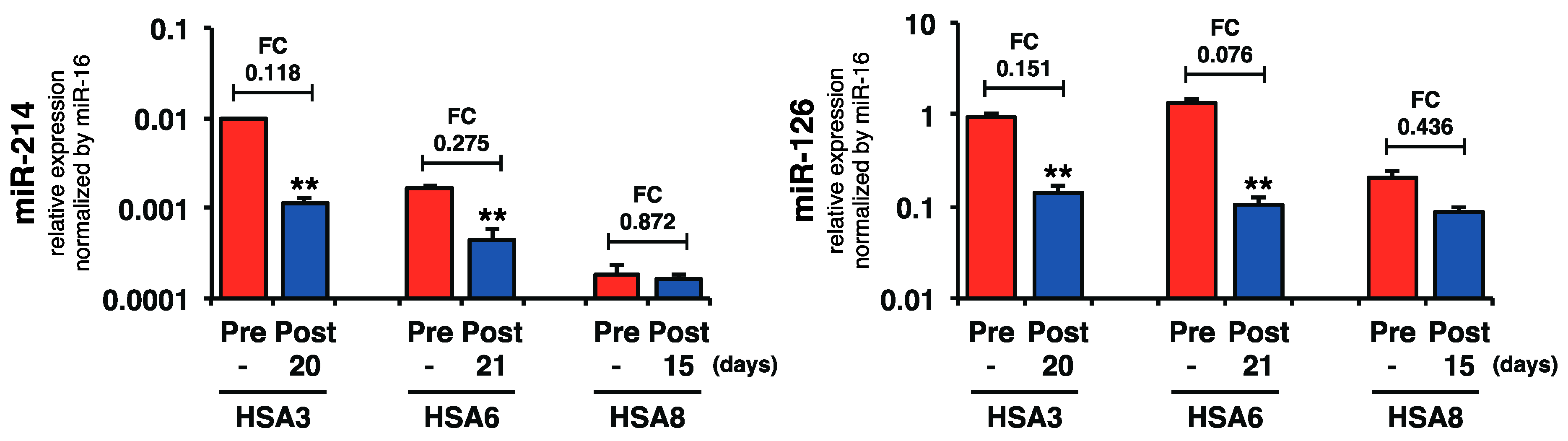
3. Discussion
4. Experimental Section
4.1. Cell Lines Used and Normal Culture Conditions
4.2. Preparation of MV-Free Culture Media
4.3. Isolation of Cell-Derived Nanoparticles
4.4. Immunoblotting
4.5. Nanoparticle Tracking Analysis
4.6. RNA Extraction from the Conditioned Media
4.7. Collection and Handling of Clinical Samples
4.8. RNA Extraction from Plasma
4.9. miRNA Quantitative Real-Time Polymerase Chain Reaction (miRNA qRT-PCR)
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abraham, J.A.; Hornicek, F.J.; Kaufman, A.M.; Harmon, D.C.; Springfield, D.S.; Raskin, K.A.; Mankin, H.J.; Kirsch, D.G.; Rosenberg, A.E.; Nielsen, G.P.; et al. Treatment and outcome of 82 patients with angiosarcoma. Ann. Surg. Oncol. 2007, 14, 1953–1967. [Google Scholar] [CrossRef] [PubMed]
- Coindre, J.M.; Terrier, P.; Guillou, L.; Le Doussal, V.; Collin, F.; Ranchere, D.; Sastre, X.; Vilain, M.O.; Bonichon, F.; N'Guyen Bui, B. Predictive value of grade for metastasis development in the main histologic types of adult soft tissue sarcomas: A study of 1240 patients from the French Federation of Cancer Centers Sarcoma Group. Cancer 2001, 91, 1914–1926. [Google Scholar] [CrossRef]
- Hammer, A.S.; Couto, C.G.; Filppi, J.; Getzy, D.; Shank, K. Efficacy and toxicity of VAC chemotherapy (vincristine, doxorubicin, and cyclophosphamide) in dogs with hemangiosarcoma. J. Vet. Intern. Med. 1991, 5, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Ivancic, M.; Long, F.; Seiler, G.S. Contrast harmonic ultrasonography of splenic masses and associated liver nodules in dogs. J. Am. Vet. Med. Assoc. 2009, 234, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.M.; Miller, N.; Kerin, M.J. MiRNAs as biomarkers and therapeutic targets in cancer. Curr. Opin. Pharmacol. 2010, 10, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Taverna, S.; Amodeo, V.; Saieva, L.; Russo, A.; Giallombardo, M.; De Leo, G.; Alessandro, R. Exosomal shuttling of miR-126 in endothelial cells modulates adhesive and migratory abilities of chronic myelogenous leukemia cells. Mol. Cancer 2014, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Van Balkom, B.W.; de Jong, O.G.; Smits, M.; Brummelman, J.; den Ouden, K.; de Bree, P.M.; van Eijndhoven, M.A.; Pegtel, D.M.; Stoorvogel, W.; Wurdinger, T.; et al. Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells. Blood 2013, 121, 3997–4006. [Google Scholar] [CrossRef] [PubMed]
- Van Mil, A.; Grundmann, S.; Goumans, M.J.; Lei, Z.; Oerlemans, M.I.; Jaksani, S.; Doevendans, P.A.; Sluijter, J.P. MicroRNA-214 inhibits angiogenesis by targeting Quaking and reducing angiogenic growth factor release. Cardiovasc. Res. 2012, 93, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Masuzawa, M.; Fujimura, T.; Hamada, Y.; Fujita, Y.; Hara, H.; Nishiyama, S.; Katsuoka, K.; Tamauchi, H.; Sakurai, Y. Establishment of a human hemangiosarcoma cell line (ISO-HAS). Int. J. Cancer 1999, 81, 305–308. [Google Scholar] [CrossRef]
- Hoshina, D.; Abe, R.; Yoshioka, N.; Saito, N.; Hata, H.; Fujita, Y.; Aoyagi, S.; Shimizu, H. Establishment of a novel experimental model of human angiosarcoma and a VEGF-targeting therapeutic experiment. J. Dermatol. Sci. 2013, 70, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Murai, A.; Asa, S.A.; Kodama, A.; Hirata, A.; Yanai, T.; Sakai, H. Constitutive phosphorylation of the mTORC2/Akt/4E-BP1 pathway in newly derived canine hemangiosarcoma cell lines. BMC Vet. Res. 2012, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Ades, E.W.; Candal, F.J.; Swerlick, R.A.; George, V.G.; Summers, S.; Bosse, D.C.; Lawley, T.J. HMEC-1: Establishment of an Immortalized Human Microvascular Endothelial Cell Line. J. Investig. Dermatol. 1992, 99, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Liao, B.Y.; Yu, L.; Gao, X.; Lu, S.; Wang, S.; Dai, Z.; Zhang, X.; Chen, Q.; et al. Human miR-1228 as a stable endogenous control for the quantification of circulating microRNAs in cancer patients. Int. J. Cancer 2014, 135, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Hamilton, R.; Mandal, C.C. miR-214: A potential biomarker and therapeutic for different cancers. Future Oncol. 2015, 11, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids—The mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Heishima, K.; Mori, T.; Sakai, H.; Sugito, N.; Murakami, M.; Yamada, N.; Akao, Y.; Maruo, K. MicroRNA-214 Promotes Apoptosis in Canine Hemangiosarcoma by Targeting the COP1-p53 Axis. PLoS ONE 2015, 10, 137361. [Google Scholar]
- Yamada, N.; Tsujimura, N.; Kumazaki, M.; Shinohara, H.; Taniguchi, K.; Nakagawa, Y.; Naoe, T.; Akao, Y. Colorectal cancer cell-derived microvesicles containing microRNA-1246 promote angiogenesis by activating Smad 1/5/8 signaling elicited by PML down-regulation in endothelial cells. Biochim. Biophys. Acta 2014, 1839, 1256–1272. [Google Scholar] [CrossRef] [PubMed]
- Chun, R.; Kellihan, H.B.; Henik, R.A.; Stepien, R.L. Comparison of plasma cardiac troponin I concentrations among dogs with cardiac hemangiosarcoma, noncardiac hemangiosarcoma, other neoplasms, and pericardial effusion of nonhemangiosarcoma origin. J. Am. Vet. Med. Assoc. 2010, 237, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Clifford, C.A.; Hughes, D.; Beal, M.W.; Mackin, A.J.; Henry, C.J.; Shofer, F.S.; Sorenmo, K.U. Plasma vascular endothelial growth factor concentrations in healthy dogs and dogs with hemangiosarcoma. J. Vet. Intern. Med. 2001, 15, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Thamm, D.H.; Kamstock, D.A.; Sharp, C.R.; Johnson, S.I.; Mazzaferro, E.; Herold, L.V.; Barnes, S.M.; Winkler, K.; Selting, K.A. Elevated serum thymidine kinase activity in canine splenic hemangiosarcoma*. Vet. Comp. Oncol. 2012, 10, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Miyasho, T.; Hanazono, K.; Saida, K.; Kadosawa, T.; Iwano, H.; Uchide, T. Big endothelin-1 as a tumour marker for canine haemangiosarcoma. Vet. J. 2015, 204, 269–274. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heishima, K.; Mori, T.; Ichikawa, Y.; Sakai, H.; Kuranaga, Y.; Nakagawa, T.; Tanaka, Y.; Okamura, Y.; Masuzawa, M.; Sugito, N.; et al. MicroRNA-214 and MicroRNA-126 Are Potential Biomarkers for Malignant Endothelial Proliferative Diseases. Int. J. Mol. Sci. 2015, 16, 25377-25391. https://doi.org/10.3390/ijms161025377
Heishima K, Mori T, Ichikawa Y, Sakai H, Kuranaga Y, Nakagawa T, Tanaka Y, Okamura Y, Masuzawa M, Sugito N, et al. MicroRNA-214 and MicroRNA-126 Are Potential Biomarkers for Malignant Endothelial Proliferative Diseases. International Journal of Molecular Sciences. 2015; 16(10):25377-25391. https://doi.org/10.3390/ijms161025377
Chicago/Turabian StyleHeishima, Kazuki, Takashi Mori, Yukie Ichikawa, Hiroki Sakai, Yuki Kuranaga, Takayuki Nakagawa, Yuiko Tanaka, Yasuhiko Okamura, Mikio Masuzawa, Nobuhiko Sugito, and et al. 2015. "MicroRNA-214 and MicroRNA-126 Are Potential Biomarkers for Malignant Endothelial Proliferative Diseases" International Journal of Molecular Sciences 16, no. 10: 25377-25391. https://doi.org/10.3390/ijms161025377
APA StyleHeishima, K., Mori, T., Ichikawa, Y., Sakai, H., Kuranaga, Y., Nakagawa, T., Tanaka, Y., Okamura, Y., Masuzawa, M., Sugito, N., Murakami, M., Yamada, N., Akao, Y., & Maruo, K. (2015). MicroRNA-214 and MicroRNA-126 Are Potential Biomarkers for Malignant Endothelial Proliferative Diseases. International Journal of Molecular Sciences, 16(10), 25377-25391. https://doi.org/10.3390/ijms161025377