Evaluating the Role of PTH in Promotion of Chondrosarcoma Cell Proliferation and Invasion by Inhibiting Primary Cilia Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
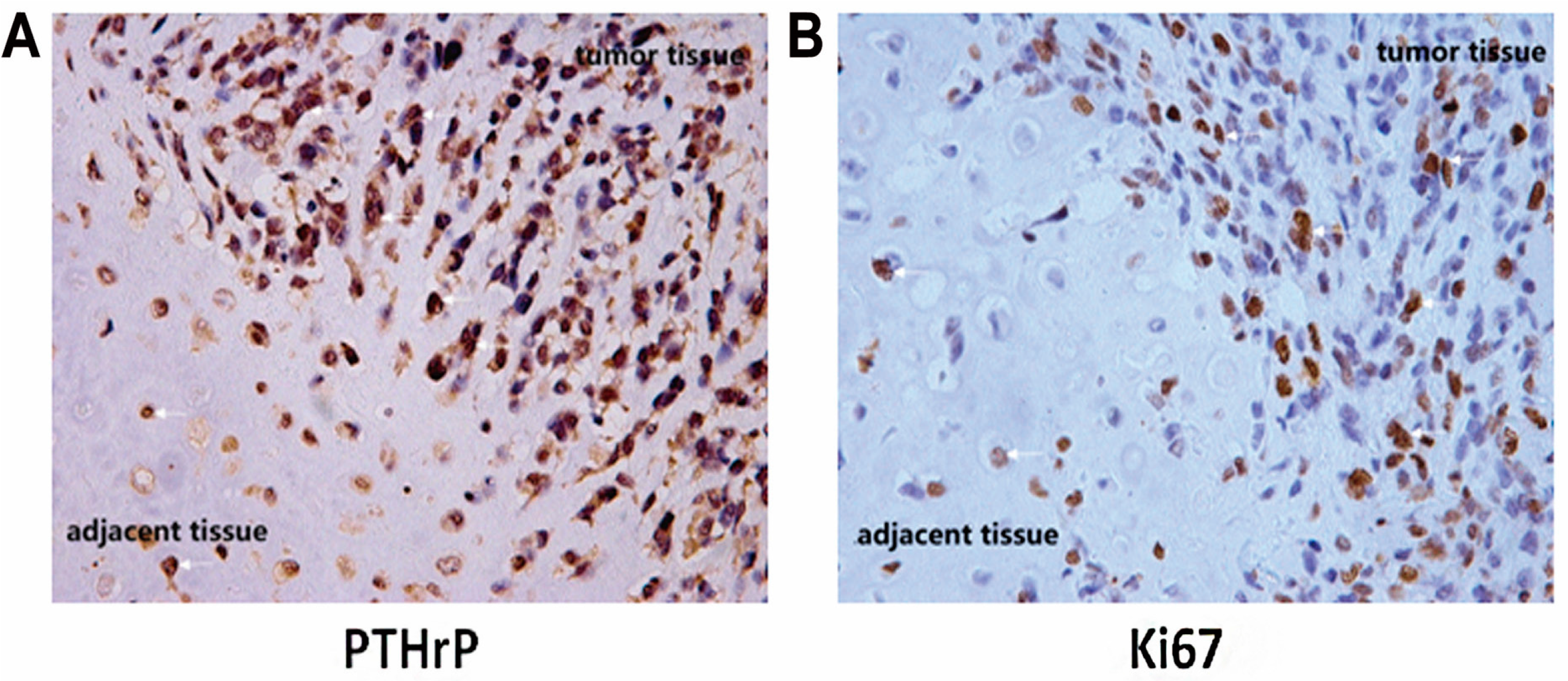
2.1. Human Chondrosarcoma Tissue Expressed an Elevated Level of PTHrP
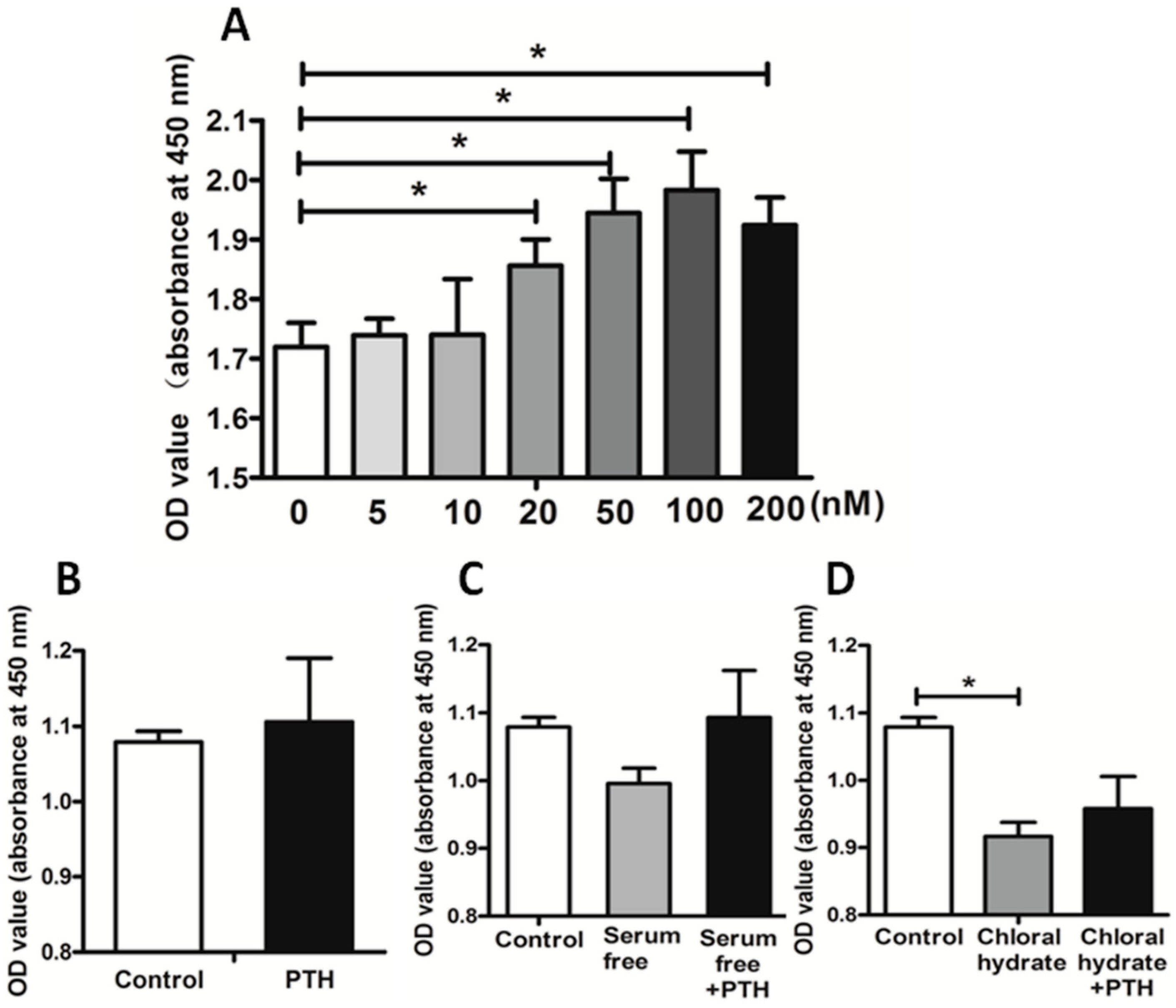
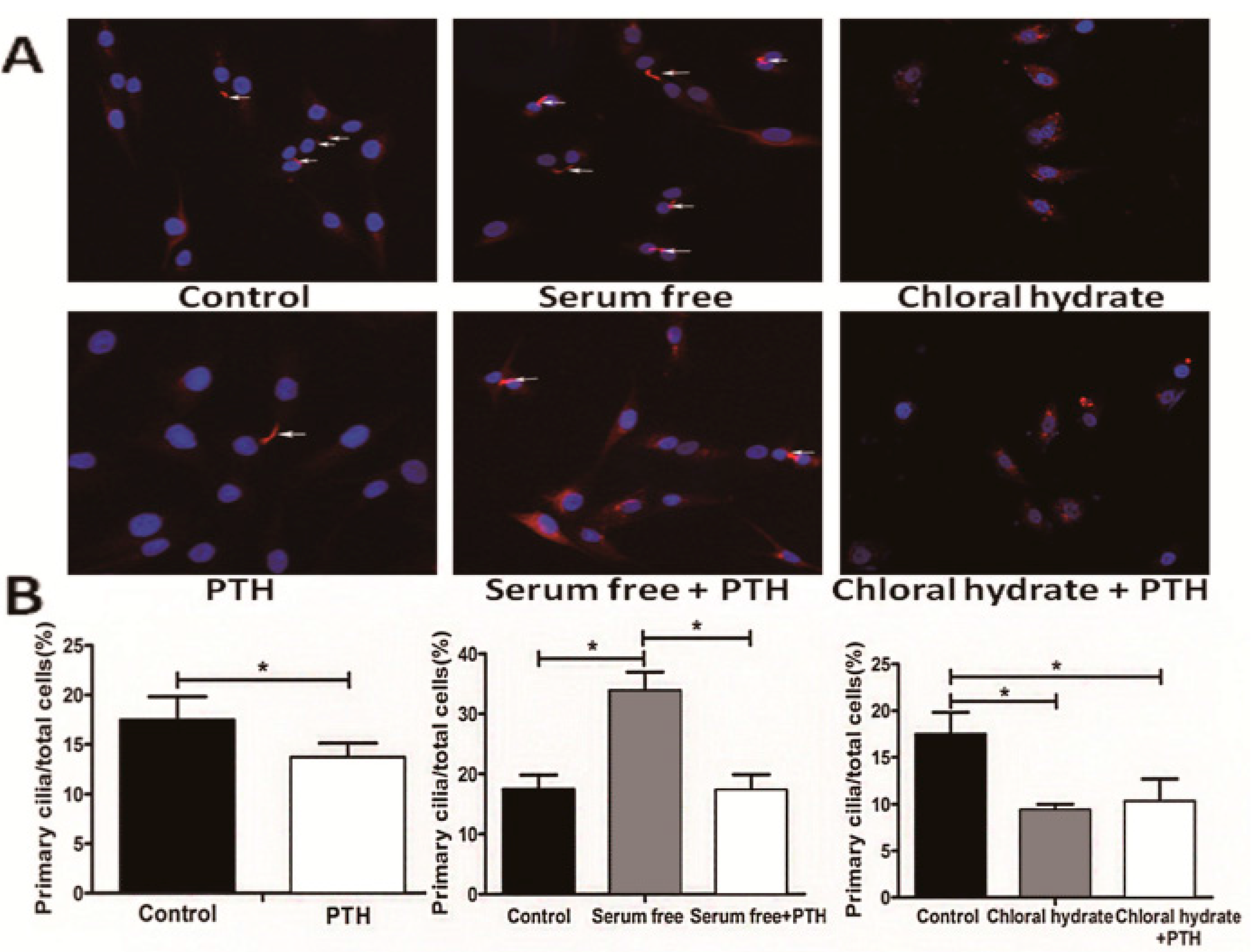
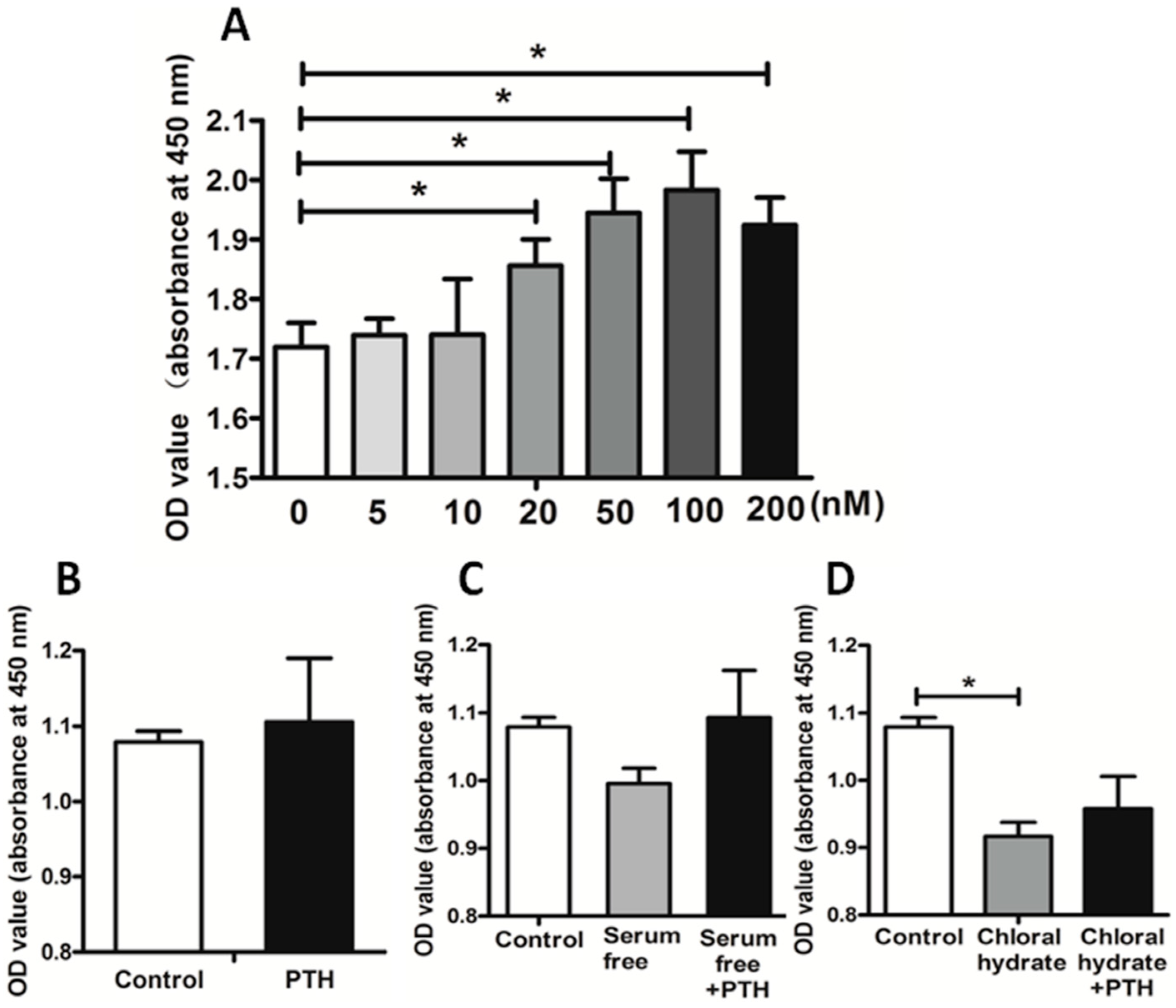
2.2. PTH Promotes Chondrosarcoma Cell Proliferation by Suppressing Primary Cilia Assembly
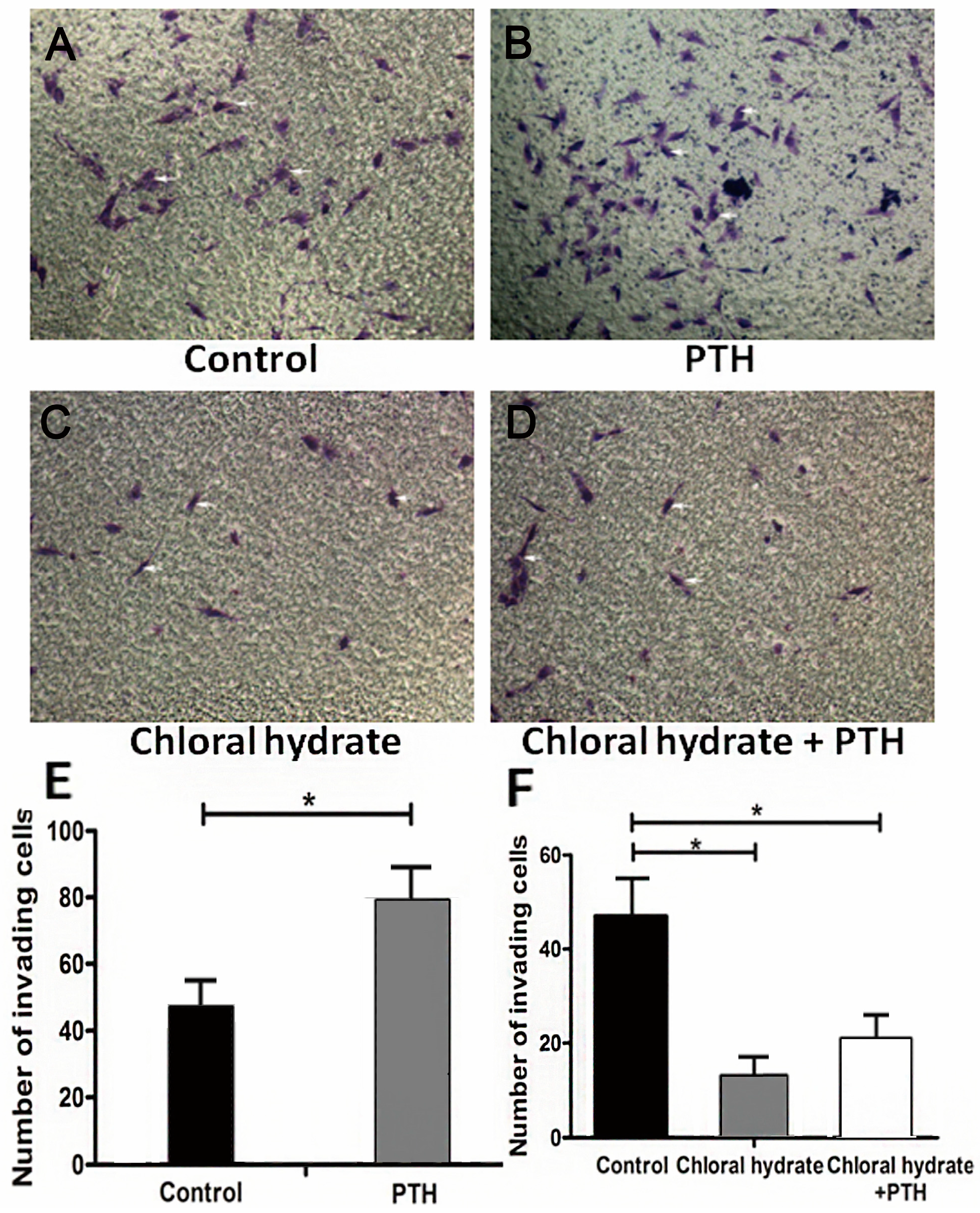
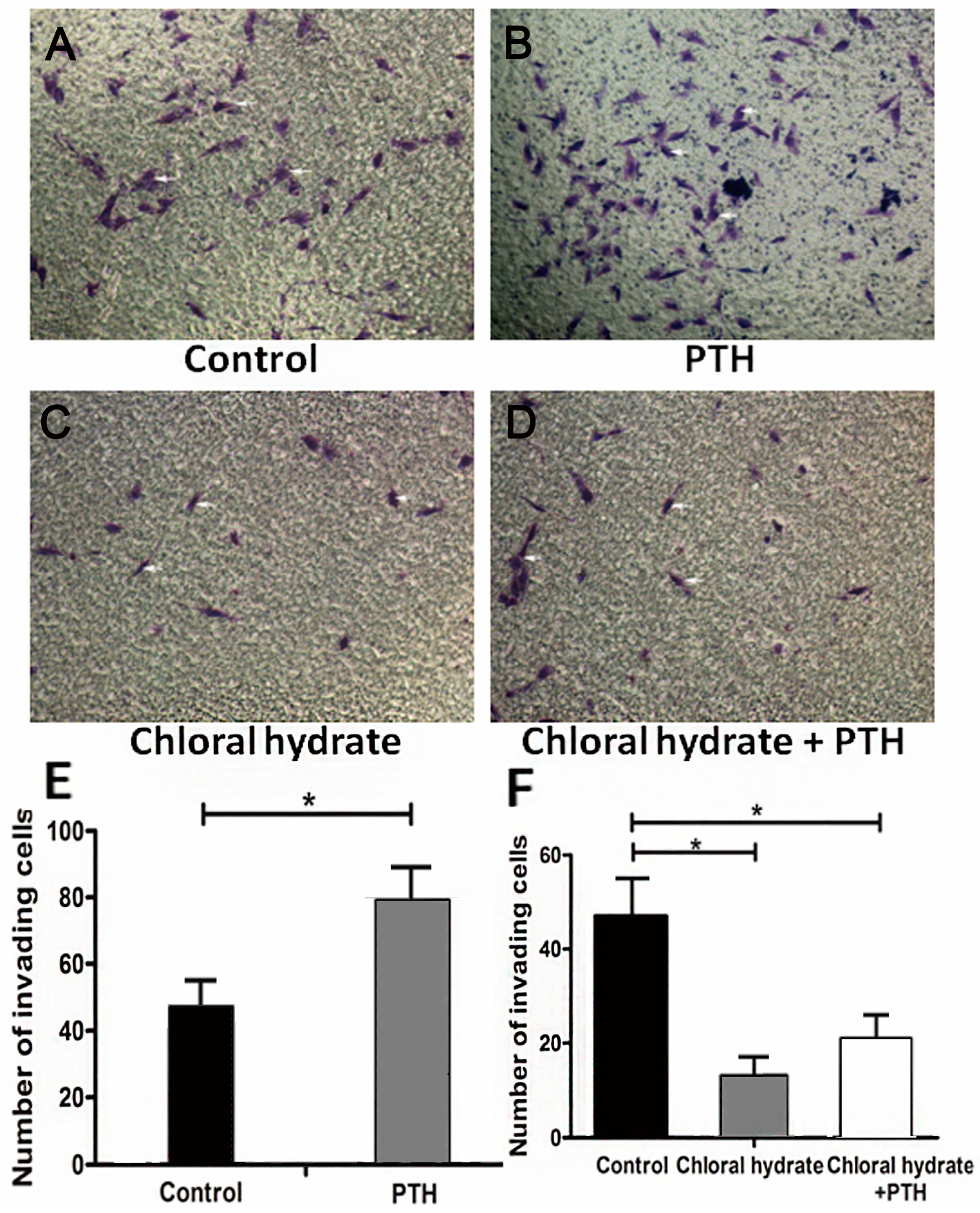
2.3. PTH Enhances Chondrosarcoma Cells Invasion Capacity by Interacting with Primary Cilia

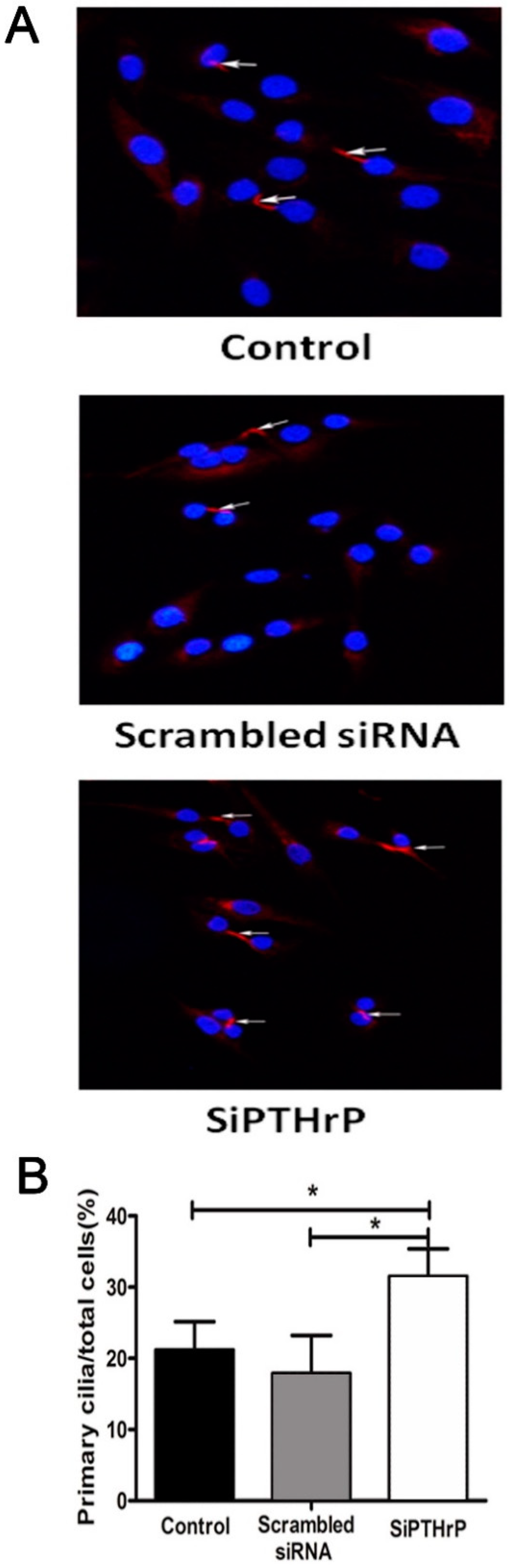
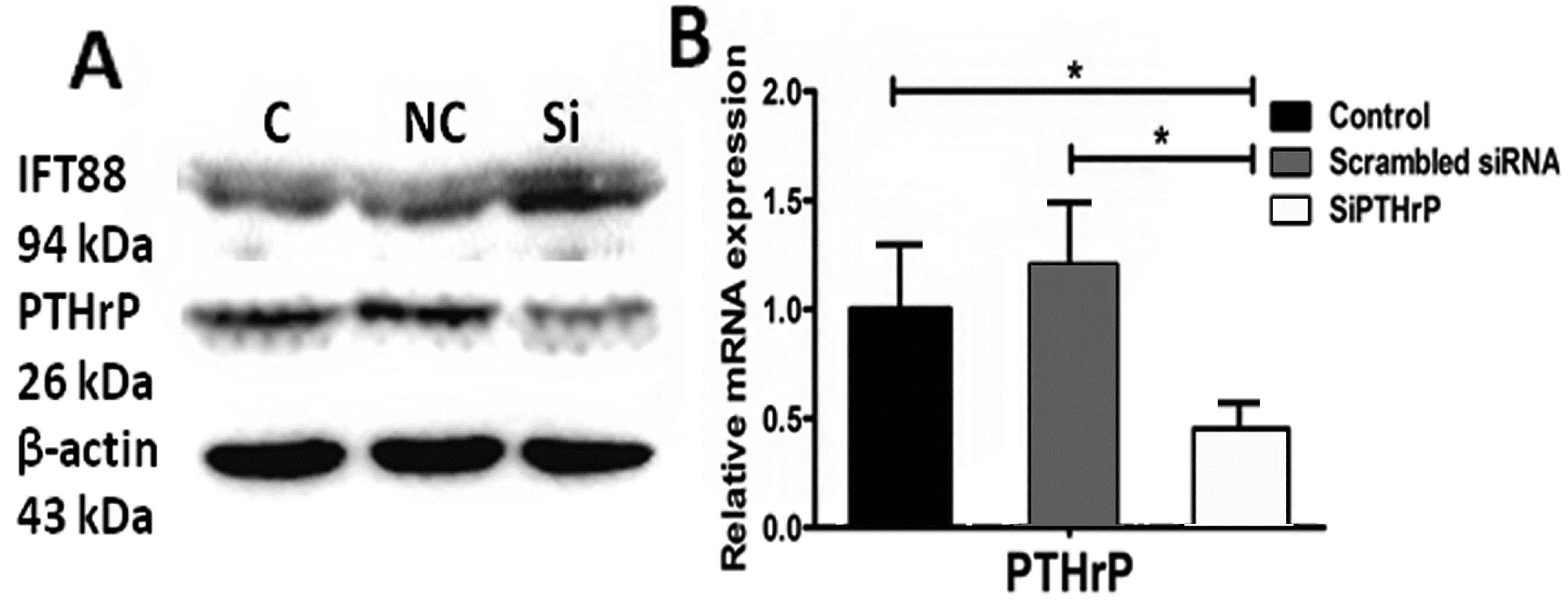
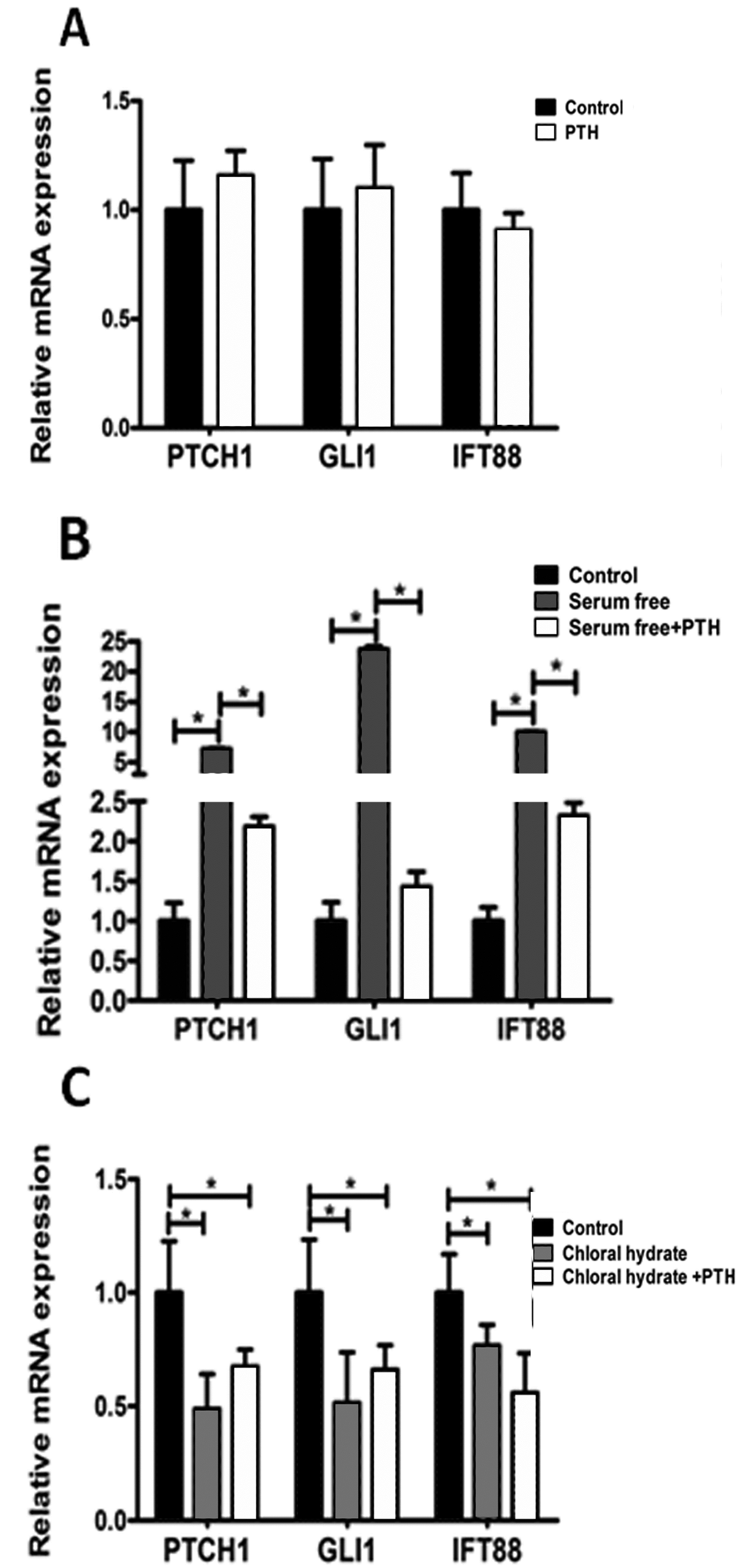
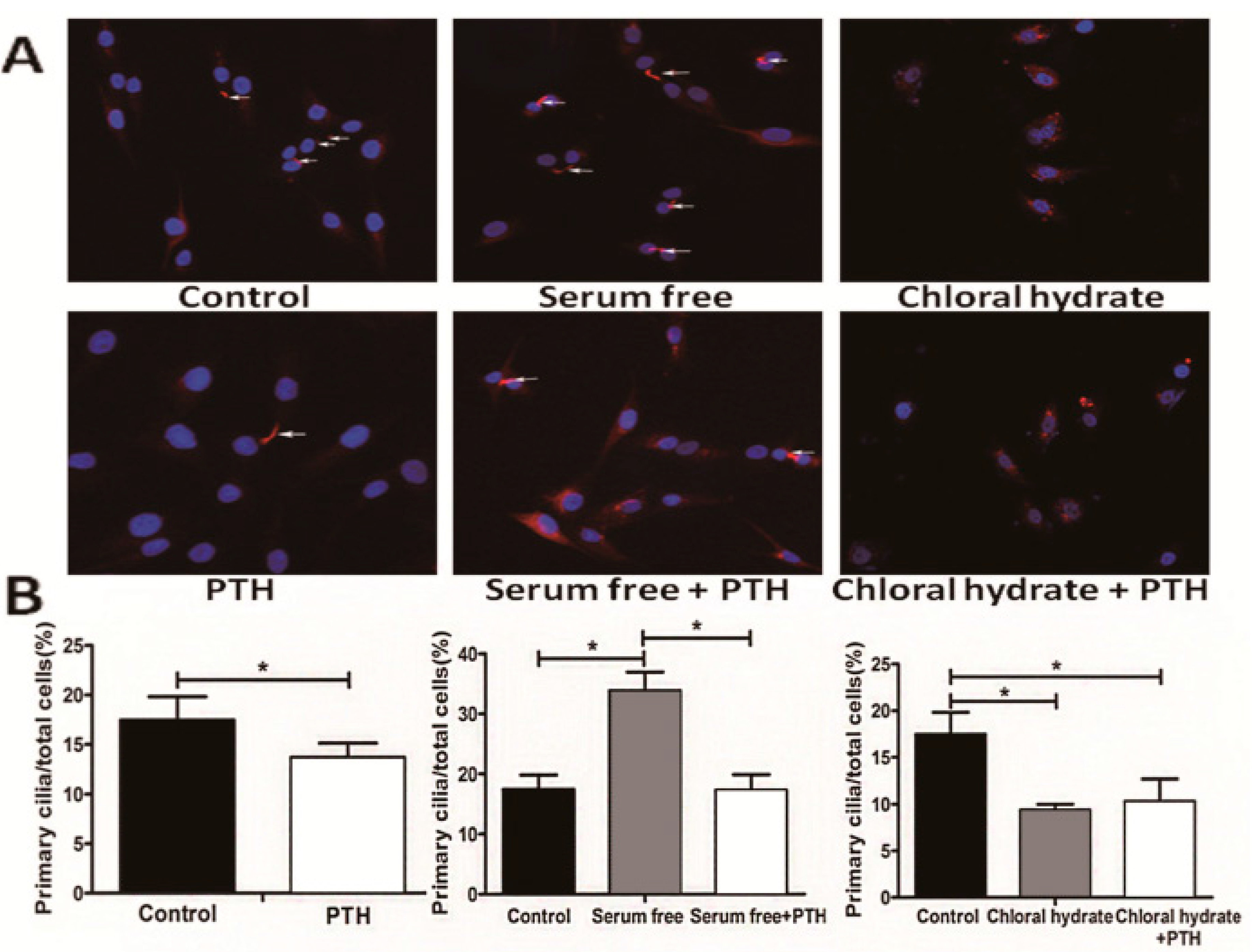
2.4. PTH/PTHrP Can Regulate Primary Cilia Expression
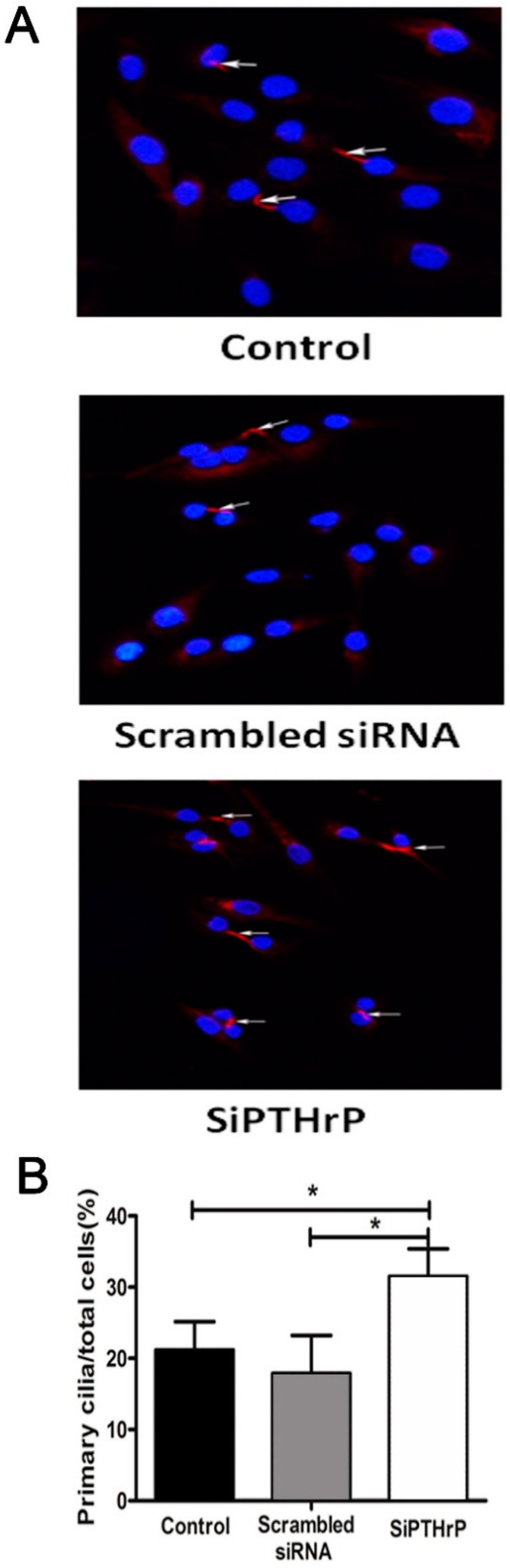
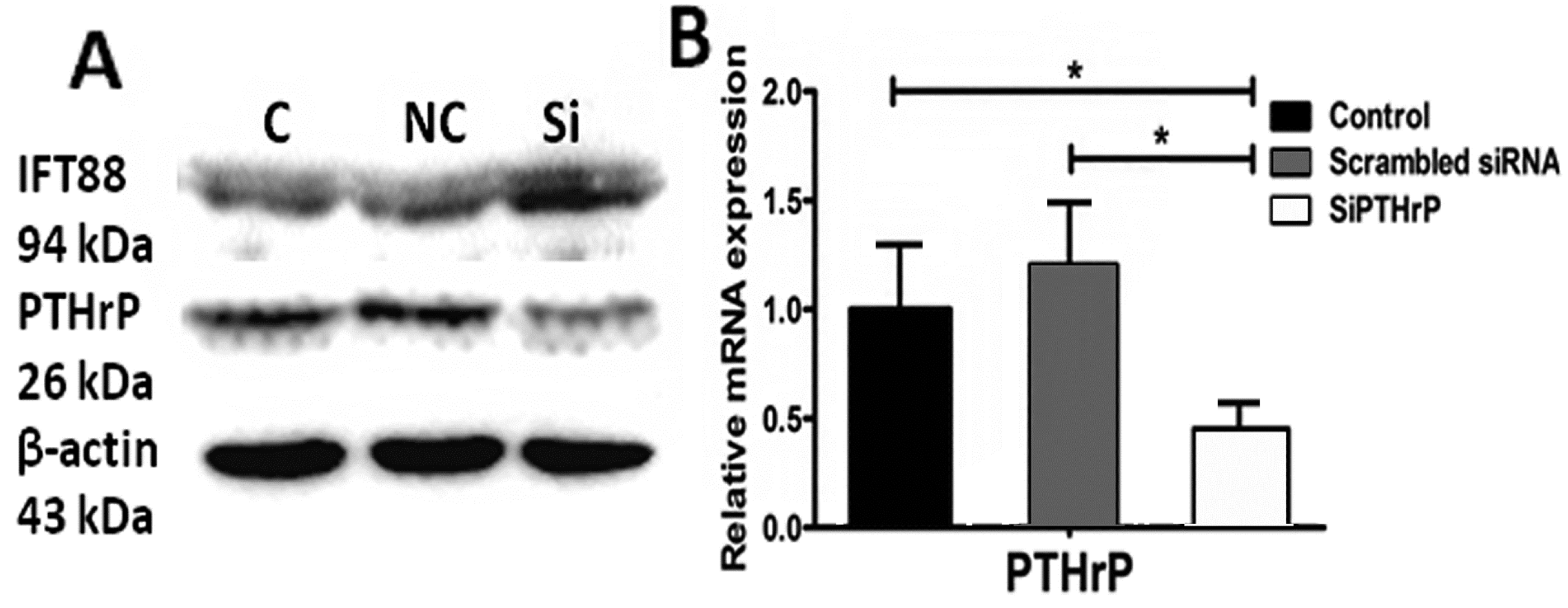
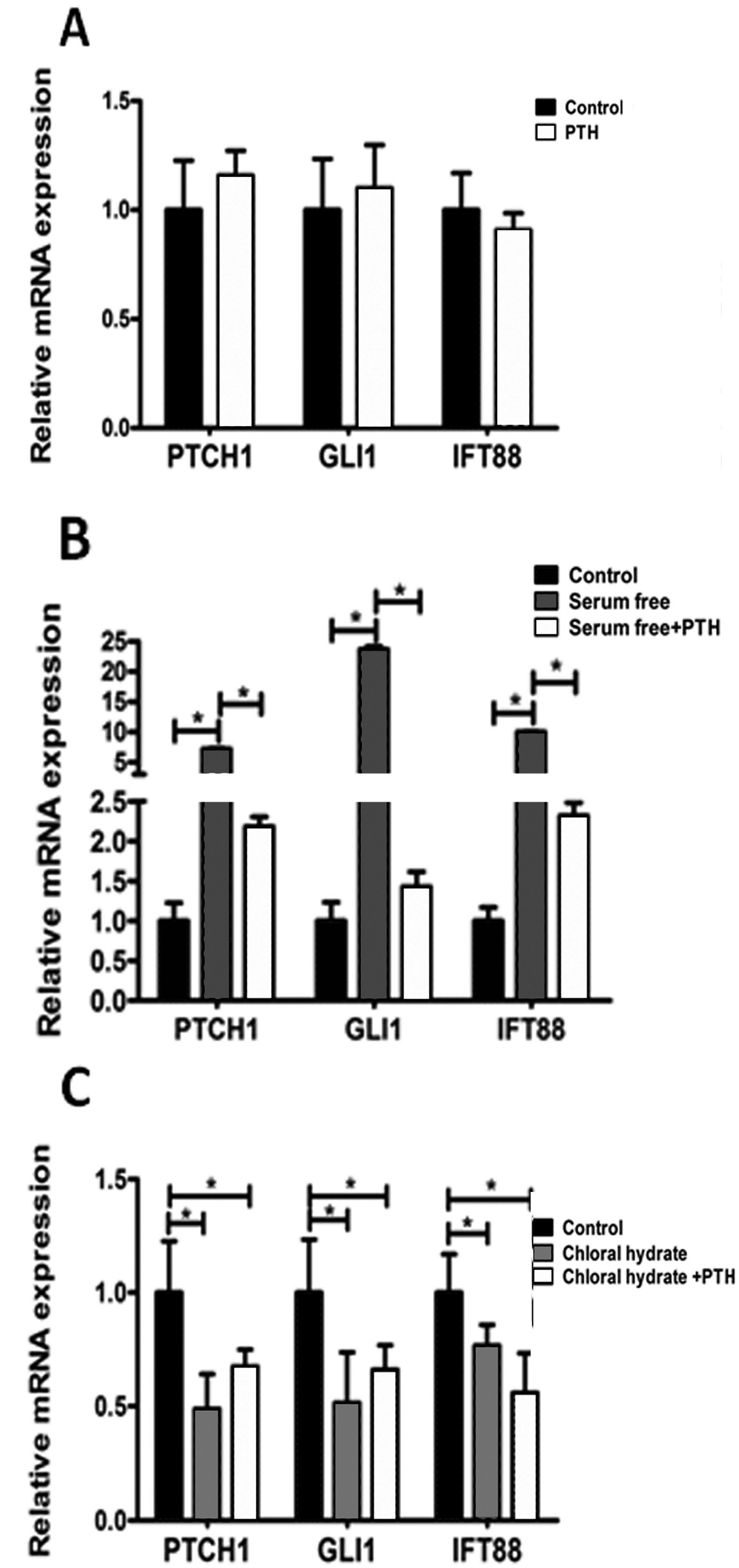
2.5. PTH/PTHrP Can Affect Hh Signaling Pathway Downstream Gene Expression
3. Experimental Section
3.1. Cells and Reagents
3.2. Cell Proliferation Test
3.3. Transwell Invasion Assay
3.4. Immunohistochemical Studies
3.5. Immunofluorescence Assay
3.6. Western Blot Analysis
3.7. Quantitative Real-Time PCR (qRT-PCR)
3.8. Small-Interfering RNA Transfection
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bjornsson, J.; McLeod, R.A.; Unni, K.K.; Ilstrup, D.M.; Pritchard, D.J. Primary chondrosarcoma of long bones and limb girdles. Cancer 1998, 83, 2105–2119. [Google Scholar]
- Schrage, Y.M.; Briaire-de Bruijn, I.H.; de Miranda, N.F.; van Oosterwijk, J.; Taminiau, A.H.; van Wezel, T.; Hogendoorn, P.C.; Bovee, J.V. Kinome profiling of chondrosarcoma reveals src-pathway activity and dasatinib as option for treatment. Cancer Res. 2009, 69, 6216–6222. [Google Scholar]
- Li, X.; Ye, H.; Cai, L.; Yu, F.; Chen, W.; Lin, R.; Zheng, C.; Xu, H.; Ye, J.; Wu, G.; et al. Millimeter wave radiation induces apoptosis via affecting the ratio of bax/bcl-2 in sw1353 human chondrosarcoma cells. Oncol. Rep. 2012, 27, 664–672. [Google Scholar]
- Bovee, J.V.; Cleton-Jansen, A.M.; Taminiau, A.H.; Hogendoorn, P.C. Emerging pathways in the development of chondrosarcoma of bone and implications for targeted treatment. Lancet Oncol. 2005, 6, 599–607. [Google Scholar]
- Gelderblom, H.; Hogendoorn, P.C.; Dijkstra, S.D.; van Rijswijk, C.S.; Krol, A.D.; Taminiau, A.H.; Bovee, J.V. The clinical approach towards chondrosarcoma. Oncologist 2008, 13, 320–329. [Google Scholar]
- Dickey, I.D.; Rose, P.S.; Fuchs, B.; Wold, L.E.; Okuno, S.H.; Sim, F.H.; Scully, S.P. Dedifferentiated chondrosarcoma: The role of chemotherapy with updated outcomes. J. Bone Jt. Surg. 2004, 86, 2412–2418. [Google Scholar]
- Johnson, R.W.; Merkel, A.R.; Danilin, S.; Nguyen, M.P.; Mundy, G.R.; Sterling, J.A. 6-thioguanine inhibition of parathyroid hormone-related protein expression is mediated by GLI2. Anticancer Res. 2011, 31, 2705–2712. [Google Scholar]
- Mak, I.W.; Turcotte, R.E.; Ghert, M. Parathyroid hormone-related protein (pthrp) modulates adhesion, migration and invasion in bone tumor cells. Bone 2013, 55, 198–207. [Google Scholar]
- Mak, I.W.; Cowan, R.W.; Turcotte, R.E.; Singh, G.; Ghert, M. Pthrp induces autocrine/paracrine proliferation of bone tumor cells through inhibition of apoptosis. PLoS One. 2011, 6, e19975. [Google Scholar]
- Johnson, R.W.; Nguyen, M.P.; Padalecki, S.S.; Grubbs, B.G.; Merkel, A.R.; Oyajobi, B.O.; Matrisian, L.M.; Mundy, G.R.; Sterling, J.A. TGF-β promotion of Gli2-induced expression of parathyroid hormone-related protein, an important osteolytic factor in bone metastasis, is independent of canonical hedgehog signaling. Cancer Res. 2011, 71, 822–831. [Google Scholar]
- Soki, F.N.; Park, S.I.; McCauley, L.K. The multifaceted actions of pthrp in skeletal metastasis. Future Oncol. 2012, 8, 803–817. [Google Scholar]
- Xu, K.; Guo, F.; Zhang, S.; Liu, C.; Wang, F.; Zhou, Z.; Chen, A. Blocking Ihh signaling pathway inhibits the proliferation and promotes the apoptosis of PSCs. J. Huazhong Univ. Sci. Technol. Med. Sci. 2009, 29, 39–44. [Google Scholar]
- Steere, N.; Chae, V.; Burke, M.; Li, F.Q.; Takemaru, K.; Kuriyama, R. A Wnt/beta-catenin pathway antagonist chibby binds cenexin at the distal end of mother centrioles and functions in primary cilia formation. PLoS One 2012, 7, e41077. [Google Scholar]
- May-Simera, H.L.; Kelley, M.W. Cilia, wnt signaling, and the cytoskeleton. Cilia 2012, 1. [Google Scholar] [CrossRef]
- Lienkamp, S.; Ganner, A.; Walz, G. Inversin, wnt signaling and primary cilia. Differentiation 2012, 83, S49–S55. [Google Scholar]
- Ho, L.; Ali, S.A.; Al-Jazrawe, M.; Kandel, R.; Wunder, J.S.; Alman, B.A. Primary cilia attenuate hedgehog signalling in neoplastic chondrocytes. Oncogene 2013, 32, 5388–5396. [Google Scholar]
- Hoey, D.A.; Tormey, S.; Ramcharan, S.; O’Brien, F.J.; Jacobs, C.R. Primary cilia-mediated mechanotransduction in human mesenchymal stem cells. Stem Cells 2012, 30, 2561–2570. [Google Scholar]
- Proulx-Bonneau, S.; Annabi, B. The primary cilium as a biomarker in the hypoxic adaptation of bone marrow-derived mesenchymal stromal cells: A role for the secreted frizzled-related proteins. Biomark. Insights 2011, 6, 107–118. [Google Scholar]
- Muhammad, H.; Rais, Y.; Miosge, N.; Ornan, E.M. The primary cilium as a dual sensor of mechanochemical signals in chondrocytes. Cell Mol. Life Sci. 2012, 69, 2101–2107. [Google Scholar]
- Goto, H.; Inoko, A.; Inagaki, M. Cell cycle progression by the repression of primary cilia formation in proliferating cells. Cell Mol. Life Sci. 2013, 70, 3893–3905. [Google Scholar]
- Irigoin, F.; Badano, J.L. Keeping the balance between proliferation and differentiation: The primary cilium. Curr. Genomics 2011, 12, 285–297. [Google Scholar]
- Yang, Y.; Roine, N.; Makela, T.P. Ccrk depletion inhibits glioblastoma cell proliferation in a cilium-dependent manner. EMBO Rep. 2013, 14, 741–747. [Google Scholar]
- Hassounah, N.B.; Bunch, T.A.; McDermott, K.M. Molecular pathways: The role of primary cilia in cancer progression and therapeutics with a focus on hedgehog signaling. Clin. Cancer Res. 2012, 18, 2429–2435. [Google Scholar]
- Basten, S.G.; Willekers, S.; Vermaat, J.S.; Slaats, G.G.; Voest, E.E.; van Diest, P.J.; Giles, R.H. Reduced cilia frequencies in human renal cell carcinomas versus neighboring parenchymal tissue. Cilia 2013, 2. [Google Scholar] [CrossRef]
- Bovee, J.V.; van den Broek, L.J.; Cleton-Jansen, A.M.; Hogendoorn, P.C. Up-regulation of PTHrP and Bcl-2 expression characterizes the progression of osteochondroma towards peripheral chondrosarcoma and is a late event in central chondrosarcoma. Lab. Invesigt. 2000, 80, 1925–1934. [Google Scholar]
- Rich, D.R.; Clark, A.L. Chondrocyte primary cilia shorten in response to osmotic challenge and are sites for endocytosis. Osteoarthr. Cartil. 2012, 20, 923–930. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, W.; Jiang, T.; Guo, F.; Xu, T.; Gong, C.; Cheng, P.; Zhao, L.; Cheng, W.; Xu, K. Evaluating the Role of PTH in Promotion of Chondrosarcoma Cell Proliferation and Invasion by Inhibiting Primary Cilia Expression. Int. J. Mol. Sci. 2014, 15, 19816-19831. https://doi.org/10.3390/ijms151119816
Xiang W, Jiang T, Guo F, Xu T, Gong C, Cheng P, Zhao L, Cheng W, Xu K. Evaluating the Role of PTH in Promotion of Chondrosarcoma Cell Proliferation and Invasion by Inhibiting Primary Cilia Expression. International Journal of Molecular Sciences. 2014; 15(11):19816-19831. https://doi.org/10.3390/ijms151119816
Chicago/Turabian StyleXiang, Wei, Ting Jiang, Fengjing Guo, Tao Xu, Chen Gong, Peng Cheng, Libo Zhao, Weiting Cheng, and Kai Xu. 2014. "Evaluating the Role of PTH in Promotion of Chondrosarcoma Cell Proliferation and Invasion by Inhibiting Primary Cilia Expression" International Journal of Molecular Sciences 15, no. 11: 19816-19831. https://doi.org/10.3390/ijms151119816
APA StyleXiang, W., Jiang, T., Guo, F., Xu, T., Gong, C., Cheng, P., Zhao, L., Cheng, W., & Xu, K. (2014). Evaluating the Role of PTH in Promotion of Chondrosarcoma Cell Proliferation and Invasion by Inhibiting Primary Cilia Expression. International Journal of Molecular Sciences, 15(11), 19816-19831. https://doi.org/10.3390/ijms151119816