Posttranscriptional Regulation of Insulin Family Ligands and Receptors

Abstract
:1. Introduction
2. Posttranscriptional Regulation of Insulin mRNA
2.1. Insulin Alternative Splicing
2.2. Regulation of Insulin Expression TTR-RBPs
2.2.1. PABP
2.2.2. HuD
2.2.3. PTBP
3. Posttranscriptional Regulation of Insulin Receptor
3.1. RBPs in Alternative Splicing of Insulin Receptor
3.2. CELF
3.3. hnRNPs
3.4. SR Proteins
3.5. MBNL
3.6. Staufen 1
3.7. RBM4
3.8. Regulation of Insulin Receptor Expression by TTR-RBPs and miRNAs
4. Posttranscriptional Regulation of IGF and IGFR
4.1. IGF and IGFR Alternative Splicing
4.2. Regulation of IGF and IGF-1R by TTR-RBPs
4.2.1. Nocturnin
4.2.2. HuR
4.2.3. hnRNP C
4.2.4. PTBP
4.2.5. Lin-28
4.2.6. IMPs
4.3. Non-Coding RNAs in IGF and IGFR Regulation
5. Concluding Remarks and Perspectives
Acknowledgments
Conflicts of Interest
Abbreviations
| CR | coding region |
| CELF | CUG-binding protein and Elav-like family |
| ELAV | embryonic lethal abnormal vision |
| hnRNP | heterogeneous nuclear ribonucleoprotein |
| IGF | insulin-like growth factor |
| IGFBP | IGF binding protein |
| IGF-1R | IGF-1 receptor |
| IIS | insulin and IGF signaling |
| IMP | IGF-2 mRNA binding proteins |
| IR | insulin receptor |
| lncRNA | Long non-coding RNA |
| MBNL | muscle blind-like |
| miR | microRNA |
| PABP | poly(A)-binding protein |
| PTBP | polypyrimidine tract-binding protein |
| RBM4 | RNA-Binding Motif Protein 4 |
| RBP | RNA-binding protein |
| SRSF1 | serine/arginine-rich splicing factor 1 |
| TTR-RBP | turnover and translation regulator RNA-binding proteins |
| UTR | untranslated region |
References
- National Diabetes Statistics. National Diabetes Information Clearinghouse, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health. Available online: http://diabetes.niddk.nih.gov/dm/pubs/statistics/ (accessed on 1 June 2013).
- Gallagher, E.J.; LeRoith, D. Minireview: IGF, insulin, and cancer. Endocrinology 2011, 152, 2546–2551. [Google Scholar]
- Arcidiacono, B.; Iiritano, S.; Nocera, A.; Possidente, K.; Nevolo, M.T.; Ventura, V.; Foti, D.; Chiefari, E.; Brunetti, A. Insulin resistance and cancer risk: An overview of the pathogenetic mechanisms. Exp. Diabetes Res 2012, 2012, 789174, :1–789174:12.. [Google Scholar]
- Itoh, N.; Sei, T.; Nose, K.; Okamoto, H. Glucose stimulation of the proinsulin synthesis in isolated pancreatic islets without increasing amount of proinsulin mRNA. FEBS Lett 1978, 93, 343–347. [Google Scholar]
- Clayton, P.E.; Banerjee, I.; Murray, P.G.; Renehan, A.G. Growth hormone, the insulin-like growth factor axis, insulin and cancer risk. Nat. Rev. Endocrinol 2011, 7, 11–24. [Google Scholar]
- Will, C.L.; Luhrmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol 2011, 3, a003707, :1–a003707:24.. [Google Scholar]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet 2008, 40, 1413–1415. [Google Scholar]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar]
- Ho, T.H.; Charlet, B.N.; Poulos, M.G.; Singh, G.; Swanson, M.S.; Cooper, T.A. Muscleblind proteins regulate alternative splicing. EMBO J 2004, 23, 3103–3112. [Google Scholar]
- Philips, A.V.; Timchenko, L.T.; Cooper, T.A. Disruption of splicing regulated by a CUG-binding protein in myotonic dystrophy. Science 1998, 280, 737–741. [Google Scholar]
- Tillmar, L.; Welsh, N. Hypoxia may increase rat insulin mRNA levels by promoting binding of the polypyrimidine tract-binding protein (PTB) to the pyrimidine-rich insulin mRNA 3′-untranslated region. Mol. Med 2002, 8, 263–272. [Google Scholar]
- Lee, E.K.; Gorospe, M. Minireview: Posttranscriptional regulation of the insulin and insulin-like growth factor systems. Endocrinology 2010, 151, 1403–1408. [Google Scholar]
- Meng, Z.; King, P.H.; Nabors, L.B.; Jackson, N.L.; Chen, C.Y.; Emanuel, P.D.; Blume, S.W. The ELAV RNA-stability factor HuR binds the 5′-untranslated region of the human IGF-IR transcript and differentially represses cap-dependent and IRES-mediated translation. Nucleic Acids Res 2005, 33, 2962–2979. [Google Scholar]
- Polesskaya, A.; Cuvellier, S.; Naguibneva, I.; Duquet, A.; Moss, E.G.; Harel-Bellan, A. Lin-28 binds IGF-2 mRNA and participates in skeletal myogenesis by increasing translation efficiency. Genes Dev 2007, 21, 1125–1138. [Google Scholar]
- Dai, N.; Rapley, J.; Angel, M.; Yanik, M.F.; Blower, M.D.; Avruch, J. mTOR phosphorylates IMP2 to promote IGF2 mRNA translation by internal ribosomal entry. Genes Dev 2011, 25, 1159–1172. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar]
- Mathonnet, G.; Fabian, M.R.; Svitkin, Y.V.; Parsyan, A.; Huck, L.; Murata, T.; Biffo, S.; Merrick, W.C.; Darzynkiewicz, E.; Pillai, R.S.; et al. MicroRNA inhibition of translation initiation in vitro by targeting the cap-binding complex eIF4F. Science 2007, 317, 1764–1767. [Google Scholar]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol 2012, 425, 3723–3730. [Google Scholar]
- Frost, R.J.; Olson, E.N. Control of glucose homeostasis and insulin sensitivity by the Let-7 family of microRNAs. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar]
- Elia, L.; Contu, R.; Quintavalle, M.; Varrone, F.; Chimenti, C.; Russo, M.A.; Cimino, V.; de Marinis, L.; Frustaci, A.; Catalucci, D.; et al. Reciprocal regulation of microRNA-1 and insulin-like growth factor-1 signal transduction cascade in cardiac and skeletal muscle in physiological and pathological conditions. Circulation 2009, 120, 2377–2385. [Google Scholar]
- Ge, Y.; Sun, Y.; Chen, J. IGF-II is regulated by microRNA-125b in skeletal myogenesis. J. Cell Biol 2011, 192, 69–81. [Google Scholar]
- Doghman, M.; El Wakil, A.; Cardinaud, B.; Thomas, E.; Wang, J.; Zhao, W.; Peralta-Del Valle, M.H.; Figueiredo, B.C.; Zambetti, G.P.; Lalli, E. Regulation of insulin-like growth factor-mammalian target of rapamycin signaling by microRNA in childhood adrenocortical tumors. Cancer Res 2010, 70, 4666–4675. [Google Scholar]
- Moran, I.; Akerman, I.; van de Bunt, M.; Xie, R.; Benazra, M.; Nammo, T.; Arnes, L.; Nakic, N.; Garcia-Hurtado, J.; Rodriguez-Segui, S.; et al. Human beta cell transcriptome analysis uncovers lncRNAs that are tissue-specific, dynamically regulated, and abnormally expressed in type 2 diabetes. Cell Metab 2012, 16, 435–448. [Google Scholar]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol 2012, 14, 659–665. [Google Scholar]
- Santoro, F.; Mayer, D.; Klement, R.M.; Warczok, K.E.; Stukalov, A.; Barlow, D.P.; Pauler, F.M. Imprinted Igf2r silencing depends on continuous Airn lncRNA expression and is not restricted to a developmental window. Development 2013, 140, 1184–1195. [Google Scholar]
- Yoon, J.H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. LincRNA-p21 suppresses target mRNA translation. Mol. Cell 2012, 47, 648–655. [Google Scholar]
- Shalev, A.; Blair, P.J.; Hoffmann, S.C.; Hirshberg, B.; Peculis, B.A.; Harlan, D.M. A proinsulin gene splice variant with increased translation efficiency is expressed in human pancreatic islets. Endocrinology 2002, 143, 2541–2547. [Google Scholar]
- Kralovicova, J.; Gaunt, T.R.; Rodriguez, S.; Wood, P.J.; Day, I.N.; Vorechovsky, I. Variants in the human insulin gene that affect pre-mRNA splicing: Is -23HphI a functional single nucleotide polymorphism at IDDM2? Diabetes 2006, 55, 260–264. [Google Scholar]
- Minn, A.H.; Lan, H.; Rabaglia, M.E.; Harlan, D.M.; Peculis, B.A.; Attie, A.D.; Shalev, A. Increased insulin translation from an insulin splice-variant overexpressed in diabetes, obesity, and insulin resistance. Mol. Endocrinol 2005, 19, 794–803. [Google Scholar]
- Panda, A.C.; Kulkarni, S.D.; Muralidharan, B.; Bakthavachalu, B.; Seshadri, V. Novel splice variant of mouse insulin2 mRNA: Implications for insulin expression. FEBS Lett 2010, 584, 1169–1173. [Google Scholar]
- Steiner, D.F.; Chan, S.J.; Welsh, J.M.; Kwok, S.C. Structure and evolution of the insulin gene. Annu. Rev. Genet 1985, 19, 463–484. [Google Scholar]
- Knight, S.W.; Docherty, K. RNA-protein interactions in the 5′ untranslated region of preproinsulin mRNA. J. Mol. Endocrinol 1992, 8, 225–234. [Google Scholar]
- Khanam, T.; Muddashetty, R.S.; Kahvejian, A.; Sonenberg, N.; Brosius, J. Poly(A)-binding protein binds to A-rich sequences via RNA-binding domains 1 + 2 and 3 + 4. RNA Biol 2006, 3, 170–177. [Google Scholar]
- Munroe, D.; Jacobson, A. mRNA poly(A) tail, a 3′ enhancer of translational initiation. Mol. Cell. Biol 1990, 10, 3441–3455. [Google Scholar]
- De Melo Neto, O.P.; Standart, N.; Martins de Sa, C. Autoregulation of poly(A)-binding protein synthesis in vitro. Nucleic Acids Res 1995, 23, 2198–2205. [Google Scholar]
- Oberer, M.; Marintchev, A.; Wagner, G. Structural basis for the enhancement of eIF4A helicase activity by eIF4G. Genes Dev 2005, 19, 2212–2223. [Google Scholar]
- Kulkarni, S.D.; Muralidharan, B.; Panda, A.C.; Bakthavachalu, B.; Vindu, A.; Seshadri, V. Glucose-stimulated translation regulation of insulin by the 5′ UTR-binding proteins. J. Biol. Chem 2011, 286, 14146–14156. [Google Scholar]
- Hinman, M.N.; Lou, H. Diverse molecular functions of Hu proteins. Cell. Mol. Life Sci 2008, 65, 3168–3181. [Google Scholar]
- Pascale, A.; Amadio, M.; Quattrone, A. Defining a neuron: Neuronal ELAV proteins. Cell. Mol. Life Sci 2008, 65, 128–140. [Google Scholar]
- Abdelmohsen, K.; Hutchison, E.R.; Lee, E.K.; Kuwano, Y.; Kim, M.M.; Masuda, K.; Srikantan, S.; Subaran, S.S.; Marasa, B.S.; Mattson, M.P.; et al. miR-375 inhibits differentiation of neurites by lowering HuD levels. Mol. Cell. Biol 2010, 30, 4197–4210. [Google Scholar]
- Lee, E.K.; Kim, W.; Tominaga, K.; Martindale, J.L.; Yang, X.; Subaran, S.S.; Carlson, O.D.; Mercken, E.M.; Kulkarni, R.N.; Akamatsu, W.; et al. RNA-binding protein HuD controls insulin translation. Mol. Cell 2012, 45, 826–835. [Google Scholar]
- Garcia-Blanco, M.A.; Jamison, S.F.; Sharp, P.A. Identification and purification of a 62,000-dalton protein that binds specifically to the polypyrimidine tract of introns. Genes Dev 1989, 3, 1874–1886. [Google Scholar]
- Sawicka, K.; Bushell, M.; Spriggs, K.A.; Willis, A.E. Polypyrimidine-tract-binding protein: A multifunctional RNA-binding protein. Biochem. Soc. Trans 2008, 36, 641–647. [Google Scholar]
- Tillmar, L.; Carlsson, C.; Welsh, N. Control of insulin mRNA stability in rat pancreatic islets. Regulatory role of a 3′-untranslated region pyrimidine-rich sequence. J. Biol. Chem 2002, 277, 1099–1106. [Google Scholar]
- Fred, R.G.; Sandberg, M.; Pelletier, J.; Welsh, N. The human insulin mRNA is partly translated via a cap- and eIF4A-independent mechanism. Biochem. Biophys. Res. Commun 2011, 412, 693–698. [Google Scholar]
- Kulkarni, R.N.; Bruning, J.C.; Winnay, J.N.; Postic, C.; Magnuson, M.A.; Kahn, C.R. Tissue-specific knockout of the insulin receptor in pancreatic beta cells creates an insulin secretory defect similar to that in type 2 diabetes. Cell 1999, 96, 329–339. [Google Scholar]
- Kahn, C.R.; White, M.F. The insulin receptor and the molecular mechanism of insulin action. J. Clin. Invest 1988, 82, 1151–1156. [Google Scholar]
- Frattali, A.L.; Treadway, J.L.; Pessin, J.E. Transmembrane signaling by the human insulin receptor kinase. Relationship between intramolecular beta subunit trans- and cis-autophosphorylation and substrate kinase activation. J. Biol. Chem 1992, 267, 19521–19528. [Google Scholar]
- Chiefari, E.; Nevolo, M.T.; Arcidiacono, B.; Maurizio, E.; Nocera, A.; Iiritano, S.; Sgarra, R.; Possidente, K.; Palmieri, C.; Paonessa, F.; et al. HMGA1 is a novel downstream nuclear target of the insulin receptor signaling pathway. Sci. Rep 2012, 2, 251, :1–251:10.. [Google Scholar]
- Brunetti, A. Insulin Receptor. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1537–1540. [Google Scholar]
- Bouzakri, K.; Roques, M.; Gual, P.; Espinosa, S.; Guebre-Egziabher, F.; Riou, J.P.; Laville, M.; Le Marchand-Brustel, Y.; Tanti, J.F.; Vidal, H. Reduced activation of phosphatidylinositol-3 kinase and increased serine 636 phosphorylation of insulin receptor substrate-1 in primary culture of skeletal muscle cells from patients with type 2 diabetes. Diabetes 2003, 52, 1319–1325. [Google Scholar]
- Seino, S.; Bell, G.I. Alternative splicing of human insulin receptor messenger RNA. Biochem. Biophys. Res. Commun 1989, 159, 312–316. [Google Scholar]
- Mosthaf, L.; Grako, K.; Dull, T.J.; Coussens, L.; Ullrich, A.; McClain, D.A. Functionally distinct insulin receptors generated by tissue-specific alternative splicing. EMBO J 1990, 9, 2409–2413. [Google Scholar]
- Vogt, B.; Carrascosa, J.M.; Ermel, B.; Ullrich, A.; Haring, H.U. The two isotypes of the human insulin receptor (HIR-A and HIR-B) follow different internalization kinetics. Biochem. Biophys. Res. Commun 1991, 177, 1013–1018. [Google Scholar]
- McClain, D.A. Different ligand affinities of the two human insulin receptor splice variants are reflected in parallel changes in sensitivity for insulin action. Mol. Endocrinol 1991, 5, 734–739. [Google Scholar]
- Moller, D.E.; Yokota, A.; Caro, J.F.; Flier, J.S. Tissue-specific expression of two alternatively spliced insulin receptor mRNAs in man. Mol. Endocrinol 1989, 3, 1263–1269. [Google Scholar]
- Savkur, R.S.; Philips, A.V.; Cooper, T.A. Aberrant regulation of insulin receptor alternative splicing is associated with insulin resistance in myotonic dystrophy. Nat. Genet 2001, 29, 40–47. [Google Scholar]
- Vidal, H.; Auboeuf, D.; Beylot, M.; Riou, J.P. Regulation of insulin receptor mRNA splicing in rat tissues. Effect of fasting, aging, and diabetes. Diabetes 1995, 44, 1196–1201. [Google Scholar]
- Wiersma, M.M.; Auboeuf, D.; Nieuwenhuizen-Bakker, I.M.; Radder, J.K.; Riou, J.P.; Vidal, H. Insulin receptor mRNA splicing and altered metabolic control in aged and mildly insulin-deficient rats. Am. J. Physiol 1997, 272, E607–E615. [Google Scholar]
- Chang, E.T.; Donahue, J.M.; Xiao, L.; Cui, Y.; Rao, J.N.; Turner, D.J.; Twaddell, W.S.; Wang, J.Y.; Battafarano, R.J. The RNA-binding protein CUG-BP1 increases survivin expression in oesophageal cancer cells through enhanced mRNA stability. Biochem. J 2012, 446, 113–123. [Google Scholar]
- Sen, S.; Talukdar, I.; Webster, N.J. SRp20 and CUG-BP1 modulate insulin receptor exon 11 alternative splicing. Mol. Cell. Biol 2009, 29, 871–880. [Google Scholar]
- Shetty, S. Regulation of urokinase receptor mRNA stability by hnRNP C in lung epithelial cells. Mol. Cell. Biochem 2005, 272, 107–118. [Google Scholar]
- Lee, E.K.; Kim, H.H.; Kuwano, Y.; Abdelmohsen, K.; Srikantan, S.; Subaran, S.S.; Gleichmann, M.; Mughal, M.R.; Martindale, J.L.; Yang, X.; et al. hnRNP C promotes APP translation by competing with FMRP for APP mRNA recruitment to P bodies. Nat. Struct. Mol. Biol 2010, 17, 732–739. [Google Scholar]
- Paul, S.; Dansithong, W.; Kim, D.; Rossi, J.; Webster, N.J.; Comai, L.; Reddy, S. Interaction of muscleblind, CUG-BP1 and hnRNP H proteins in DM1-associated aberrant IR splicing. EMBO J 2006, 25, 4271–4283. [Google Scholar]
- Paul, S.; Dansithong, W.; Jog, S.P.; Holt, I.; Mittal, S.; Brook, J.D.; Morris, G.E.; Comai, L.; Reddy, S. Expanded CUG repeats Dysregulate RNA splicing by altering the stoichiometry of the muscleblind 1 complex. J. Biol. Chem 2011, 286, 38427–38438. [Google Scholar]
- Talukdar, I.; Sen, S.; Urbano, R.; Thompson, J.; Yates, J.R., III; Webster, N.J. hnRNP A1 and hnRNP F modulate the alternative splicing of exon 11 of the insulin receptor gene. PLoS One 2011, 6, e27869. [Google Scholar]
- Shepard, P.J.; Hertel, K.J. The SR protein family. Genome Biol 2009, 10, 242. [Google Scholar]
- Grammatikakis, I.; Goo, Y.H.; Echeverria, G.V.; Cooper, T.A. Identification of MBNL1 and MBNL3 domains required for splicing activation and repression. Nucleic Acids Res 2011, 39, 2769–2780. [Google Scholar]
- Sen, S.; Talukdar, I.; Liu, Y.; Tam, J.; Reddy, S.; Webster, N.J. Muscleblind-like 1 (Mbnl1) promotes insulin receptor exon 11 inclusion via binding to a downstream evolutionarily conserved intronic enhancer. J. Biol. Chem 2010, 285, 25426–25437. [Google Scholar]
- Lin, X.; Miller, J.W.; Mankodi, A.; Kanadia, R.N.; Yuan, Y.; Moxley, R.T.; Swanson, M.S.; Thornton, C.A. Failure of MBNL1-dependent post-natal splicing transitions in myotonic dystrophy. Hum. Mol. Genet 2006, 15, 2087–2097. [Google Scholar]
- Ravel-Chapuis, A.; Belanger, G.; Yadava, R.S.; Mahadevan, M.S.; DesGroseillers, L.; Cote, J.; Jasmin, B.J. The RNA-binding protein Staufen1 is increased in DM1 skeletal muscle and promotes alternative pre-mRNA splicing. J. Cell Biol 2012, 196, 699–712. [Google Scholar]
- Kim, Y.K.; Furic, L.; Parisien, M.; Major, F.; DesGroseillers, L.; Maquat, L.E. Staufen1 regulates diverse classes of mammalian transcripts. EMBO J 2007, 26, 2670–2681. [Google Scholar]
- Cho, H.; Kim, K.M.; Han, S.; Choe, J.; Park, S.G.; Choi, S.S.; Kim, Y.K. Staufen1-mediated mRNA decay functions in adipogenesis. Mol. Cell 2012, 46, 495–506. [Google Scholar]
- Lin, J.C.; Yan, Y.T.; Hsieh, W.K.; Peng, P.J.; Su, C.H.; Tarn, W.Y. RBM4 promotes pancreas cell differentiation and insulin expression. Mol. Cell. Biol 2013, 33, 319–327. [Google Scholar]
- Lu, C.C.; Chen, T.H.; Wu, J.R.; Chen, H.H.; Yu, H.Y.; Tarn, W.Y. Phylogenetic and molecular characterization of the splicing factor RBM4. PLoS One 2013, 8, e59092. [Google Scholar]
- Hellen, C.U.; Sarnow, P. Internal ribosome entry sites in eukaryotic mRNA molecules. Genes Dev 2001, 15, 1593–1612. [Google Scholar]
- Spriggs, K.A.; Cobbold, L.C.; Ridley, S.H.; Coldwell, M.; Bottley, A.; Bushell, M.; Willis, A.E.; Siddle, K. The human insulin receptor mRNA contains a functional internal ribosome entry segment. Nucleic Acids Res 2009, 37, 5881–5893. [Google Scholar]
- Chen, B.; Li, H.; Zeng, X.; Yang, P.; Liu, X.; Zhao, X.; Liang, S. Roles of microRNA on cancer cell metabolism. J. Transl. Med 2012, 10, 228. [Google Scholar]
- Fan, Q.; He, M.; Deng, X.; Wu, W.K.; Zhao, L.; Tang, J.; Wen, G.; Sun, X.; Liu, Y. Derepression of c-Fos caused by MicroRNA-139 down-regulation contributes to the metastasis of human hepatocellular carcinoma. Cell Biochem. Funct 2012, 31, 319–324. [Google Scholar]
- Fernandez-Valverde, S.L.; Taft, R.J.; Mattick, J.S. MicroRNAs in beta-cell biology, insulin resistance, diabetes and its complications. Diabetes 2011, 60, 1825–1831. [Google Scholar]
- Zhu, H.; Shyh-Chang, N.; Segre, A.V.; Shinoda, G.; Shah, S.P.; Einhorn, W.S.; Takeuchi, A.; Engreitz, J.M.; Hagan, J.P.; Kharas, M.G.; et al. The Lin28/let-7 axis regulates glucose metabolism. Cell 2011, 147, 81–94. [Google Scholar]
- Scharf, J.G.; Braulke, T. The role of the IGF axis in hepatocarcinogenesis. Horm. Metab. Res 2003, 35, 685–693. [Google Scholar]
- Grimberg, A. Mechanisms by which IGF-I may promote cancer. Cancer Biol. Ther 2003, 2, 630–635. [Google Scholar]
- Cohen, D.H.; LeRoith, D. Obesity, type 2 diabetes, and cancer: The insulin and IGF connection. Endocr. Relat. Cancer 2012, 19, F27–F45. [Google Scholar]
- Laron, Z. Effects of growth hormone and insulin-like growth factor 1 deficiency on ageing and longevity. Novartis Found. Symp 2002, 242, 125–137, ; discussion 137–142.. [Google Scholar]
- Philippou, A.; Maridaki, M.; Halapas, A.; Koutsilieris, M. The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology. In Vivo 2007, 21, 45–54. [Google Scholar]
- Velloso, C.P.; Harridge, S.D. Insulin-like growth factor-I E peptides: Implications for aging skeletal muscle. Scand. J. Med. Sci. Sports 2010, 20, 20–27. [Google Scholar]
- Goldspink, G.; Yang, S.Y. The splicing of the IGF-I gene to yield different muscle growth factors. Adv. Genet 2004, 52, 23–49. [Google Scholar]
- Matheny, R.W., Jr; Nindl, B.C. Loss of IGF-IEa or IGF-IEb impairs myogenic differentiation. Endocrinology 2011, 152, 1923–1934. [Google Scholar]
- Coleman, M.E.; DeMayo, F.; Yin, K.C.; Lee, H.M.; Geske, R.; Montgomery, C.; Schwartz, R.J. Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J. Biol. Chem 1995, 270, 12109–12116. [Google Scholar]
- Barton, E.R.; Morris, L.; Musaro, A.; Rosenthal, N.; Sweeney, H.L. Muscle-specific expression of insulin-like growth factor I counters muscle decline in mdx mice. J. Cell Biol 2002, 157, 137–148. [Google Scholar]
- Dai, Z.; Wu, F.; Yeung, E.W.; Li, Y. IGF-IEc expression, regulation and biological function in different tissues. Growth Horm. IGF Res 2010, 20, 275–281. [Google Scholar]
- Ates, K.; Yang, S.Y.; Orrell, R.W.; Sinanan, A.C.; Simons, P.; Solomon, A.; Beech, S.; Goldspink, G.; Lewis, M.P. The IGF-I splice variant MGF increases progenitor cells in ALS, dystrophic, and normal muscle. FEBS Lett 2007, 581, 2727–2732. [Google Scholar]
- Kandalla, P.K.; Goldspink, G.; Butler-Browne, G.; Mouly, V. Mechano Growth Factor E peptide (MGF-E), derived from an isoform of IGF-1, activates human muscle progenitor cells and induces an increase in their fusion potential at different ages. Mech. Ageing Dev 2011, 132, 154–162. [Google Scholar]
- Weller, P.A.; Dickson, M.C.; Huskisson, N.S.; Dauncey, M.J.; Buttery, P.J.; Gilmour, R.S. The porcine insulin-like growth factor-I gene: Characterization and expression of alternate transcription sites. J. Mol. Endocrinol 1993, 11, 201–211. [Google Scholar]
- Yang, H.; Adamo, M.L.; Koval, A.P.; McGuinness, M.C.; Ben-Hur, H.; Yang, Y.; LeRoith, D.; Roberts, C.T., Jr. Alternative leader sequences in insulin-like growth factor I mRNAs modulate translational efficiency and encode multiple signal peptides. Mol. Endocrinol. 1995, 9, 1380–1395. [Google Scholar]
- Fuqua, J.S.; Derr, M.; Rosenfeld, R.G.; Hwa, V. Identification of a novel heterozygous IGF1 splicing mutation in a large kindred with familial short stature. Horm. Res. Paediatr 2012, 78, 59–66. [Google Scholar]
- Ekstrom, T.J.; Cui, H.; Li, X.; Ohlsson, R. Promoter-specific IGF2 imprinting status and its plasticity during human liver development. Development 1995, 121, 309–316. [Google Scholar]
- Monk, D.; Sanches, R.; Arnaud, P.; Apostolidou, S.; Hills, F.A.; Abu-Amero, S.; Murrell, A.; Friess, H.; Reik, W.; Stanier, P.; et al. Imprinting of IGF2 P0 transcript and novel alternatively spliced INS-IGF2 isoforms show differences between mouse and human. Hum. Mol. Genet 2006, 15, 1259–1269. [Google Scholar]
- Ohlsson, R.; Hedborg, F.; Holmgren, L.; Walsh, C.; Ekstrom, T.J. Overlapping patterns of IGF2 and H19 expression during human development: Biallelic IGF2 expression correlates with a lack of H19 expression. Development 1994, 120, 361–368. [Google Scholar]
- Smith, P.J.; Spurrell, E.L.; Coakley, J.; Hinds, C.J.; Ross, R.J.; Krainer, A.R.; Chew, S.L. An exonic splicing enhancer in human IGF-I pre-mRNA mediates recognition of alternative exon 5 by the serine-arginine protein splicing factor-2/alternative splicing factor. Endocrinology 2002, 143, 146–154. [Google Scholar]
- Goldspink, G. Mechanical signals, IGF-I gene splicing, and muscle adaptation. Physiology 2005, 20, 232–238. [Google Scholar]
- Abbott, A.M.; Bueno, R.; Pedrini, M.T.; Murray, J.M.; Smith, R.J. Insulin-like growth factor I receptor gene structure. J. Biol. Chem 1992, 267, 10759–10763. [Google Scholar]
- Yee, D.; Lebovic, G.S.; Marcus, R.R.; Rosen, N. Identification of an alternate type I insulin-like growth factor receptor beta subunit mRNA transcript. J. Biol. Chem 1989, 264, 21439–21441. [Google Scholar]
- Condorelli, G.; Bueno, R.; Smith, R.J. Two alternatively spliced forms of the human insulin-like growth factor I receptor have distinct biological activities and internalization kinetics. J. Biol. Chem 1994, 269, 8510–8516. [Google Scholar]
- Kawai, M.; Delany, A.M.; Green, C.B.; Adamo, M.L.; Rosen, C.J. Nocturnin suppresses igf1 expression in bone by targeting the 3′ untranslated region of igf1 mRNA. Endocrinology 2010, 151, 4861–4870. [Google Scholar]
- Wilusz, C.J.; Wilusz, J. Bringing the role of mRNA decay in the control of gene expression into focus. Trends Genet 2004, 20, 491–497. [Google Scholar]
- Green, C.B.; Douris, N.; Kojima, S.; Strayer, C.A.; Fogerty, J.; Lourim, D.; Keller, S.R.; Besharse, J.C. Loss of Nocturnin, a circadian deadenylase, confers resistance to hepatic steatosis and diet-induced obesity. Proc. Natl. Acad. Sci. USA 2007, 104, 9888–9893. [Google Scholar]
- Tominaga, K.; Srikantan, S.; Lee, E.K.; Subaran, S.S.; Martindale, J.L.; Abdelmohsen, K.; Gorospe, M. Competitive regulation of nucleolin expression by HuR and miR-494. Mol. Cell. Biol 2011, 31, 4219–4231. [Google Scholar]
- Mazan-Mamczarz, K.; Galban, S.; Lopez de Silanes, I.; Martindale, J.L.; Atasoy, U.; Keene, J.D.; Gorospe, M. RNA-binding protein HuR enhances p53 translation in response to ultraviolet light irradiation. Proc. Natl. Acad. Sci. USA 2003, 100, 8354–8359. [Google Scholar]
- Srikantan, S.; Gorospe, M. HuR function in disease. Front. Biosci 2012, 17, 189–205. [Google Scholar]
- Yeh, C.H.; Hung, L.Y.; Hsu, C.; Le, S.Y.; Lee, P.T.; Liao, W.L.; Lin, Y.T.; Chang, W.C.; Tseng, J.T. RNA-binding protein HuR interacts with thrombomodulin 5′ untranslated region and represses internal ribosome entry site-mediated translation under IL-1 beta treatment. Mol. Biol. Cell 2008, 19, 3812–3822. [Google Scholar]
- Meng, Z.; Jackson, N.L.; Choi, H.; King, P.H.; Emanuel, P.D.; Blume, S.W. Alterations in RNA-binding activities of IRES-regulatory proteins as a mechanism for physiological variability and pathological dysregulation of IGF-IR translational control in human breast tumor cells. J. Cell. Physiol 2008, 217, 172–183. [Google Scholar]
- Abdelmohsen, K.; Pullmann, R., Jr; Lal, A.; Kim, H.H.; Galban, S.; Yang, X.; Blethrow, J.D.; Walker, M.; Shubert, J.; Gillespie, D.A.; et al. Phosphorylation of HuR by Chk2 regulates SIRT1 expression. Mol. Cell 2007, 25, 543–557. [Google Scholar]
- Yaman, I.; Fernandez, J.; Sarkar, B.; Schneider, R.J.; Snider, M.D.; Nagy, L.E.; Hatzoglou, M. Nutritional control of mRNA stability is mediated by a conserved AU-rich element that binds the cytoplasmic shuttling protein HuR. J. Biol. Chem 2002, 277, 41539–41546. [Google Scholar]
- Mitchell, S.A.; Spriggs, K.A.; Coldwell, M.J.; Jackson, R.J.; Willis, A.E. The Apaf-1 internal ribosome entry segment attains the correct structural conformation for function via interactions with PTB and unr. Mol. Cell 2003, 11, 757–771. [Google Scholar]
- Giraud, S.; Greco, A.; Brink, M.; Diaz, J.J.; Delafontaine, P. Translation initiation of the insulin-like growth factor I receptor mRNA is mediated by an internal ribosome entry site. J. Biol. Chem 2001, 276, 5668–5675. [Google Scholar]
- Thornton, J.E.; Chang, H.M.; Piskounova, E.; Gregory, R.I. Lin28-mediated control of let-7 microRNA expression by alternative TUTases Zcchc11 (TUT4) and Zcchc6 (TUT7). RNA 2012, 18, 1875–1885. [Google Scholar]
- Nielsen, J.; Christiansen, J.; Lykke-Andersen, J.; Johnsen, A.H.; Wewer, U.M.; Nielsen, F.C. A family of insulin-like growth factor II mRNA-binding proteins represses translation in late development. Mol. Cell. Biol 1999, 19, 1262–1270. [Google Scholar]
- Liao, B.; Hu, Y.; Herrick, D.J.; Brewer, G. The RNA-binding protein IMP-3 is a translational activator of insulin-like growth factor II leader-3 mRNA during proliferation of human K562 leukemia cells. J. Biol. Chem 2005, 280, 18517–18524. [Google Scholar]
- Pedersen, S.K.; Christiansen, J.; Hansen, T.; Larsen, M.R.; Nielsen, F.C. Human insulin-like growth factor II leader 2 mediates internal initiation of translation. Biochem. J 2002, 363, 37–44. [Google Scholar]
- Nielsen, F.C.; Ostergaard, L.; Nielsen, J.; Christiansen, J. Growth-dependent translation of IGF-II mRNA by a rapamycin-sensitive pathway. Nature 1995, 377, 358–362. [Google Scholar]
- Suvasini, R.; Shruti, B.; Thota, B.; Shinde, S.V.; Friedmann-Morvinski, D.; Nawaz, Z.; Prasanna, K.V.; Thennarasu, K.; Hegde, A.S.; Arivazhagan, A.; et al. Insulin growth factor-2 binding protein 3 (IGF2BP3) is a glioblastoma-specific marker that activates phosphatidylinositol 3-kinase/mitogen-activated protein kinase (PI3K/MAPK) pathways by modulating IGF-2. J. Biol. Chem 2011, 286, 25882–25890. [Google Scholar]
- Schonrock, N.; Harvey, R.P.; Mattick, J.S. Long noncoding RNAs in cardiac development and pathophysiology. Circ. Res 2012, 111, 1349–1362. [Google Scholar]
- Pullmann, R., Jr; Kim, H.H.; Abdelmohsen, K.; Lal, A.; Martindale, J.L.; Yang, X.; Gorospe, M. Analysis of turnover and translation regulatory RNA-binding protein expression through binding to cognate mRNAs. Mol. Cell. Biol. 2007, 27, 6265–6278. [Google Scholar]
- Kwiecinski, M.; Elfimova, N.; Noetel, A.; Tox, U.; Steffen, H.M.; Hacker, U.; Nischt, R.; Dienes, H.P.; Odenthal, M. Expression of platelet-derived growth factor-C and insulin-like growth factor I in hepatic stellate cells is inhibited by miR-29. Lab. Invest 2012, 92, 978–987. [Google Scholar]
- Yu, X.Y.; Song, Y.H.; Geng, Y.J.; Lin, Q.X.; Shan, Z.X.; Lin, S.G.; Li, Y. Glucose induces apoptosis of cardiomyocytes via microRNA-1 and IGF-1. Biochem. Biophys. Res. Commun 2008, 376, 548–552. [Google Scholar]
- Shan, Z.X.; Lin, Q.X.; Fu, Y.H.; Deng, C.Y.; Zhou, Z.L.; Zhu, J.N.; Liu, X.Y.; Zhang, Y.Y.; Li, Y.; Lin, S.G.; et al. Upregulated expression of miR-1/miR-206 in a rat model of myocardial infarction. Biochem. Biophys. Res. Commun 2009, 381, 597–601. [Google Scholar]
- Wang, X.H.; Qian, R.Z.; Zhang, W.; Chen, S.F.; Jin, H.M.; Hu, R.M. MicroRNA-320 expression in myocardial microvascular endothelial cells and its relationship with insulin-like growth factor-1 in type 2 diabetic rats. Clin. Exp. Pharmacol. Physiol 2009, 36, 181–188. [Google Scholar]
- Jiang, L.; Liu, X.; Chen, Z.; Jin, Y.; Heidbreder, C.E.; Kolokythas, A.; Wang, A.; Dai, Y.; Zhou, X. MicroRNA-7 targets IGF1R (insulin-like growth factor 1 receptor) in tongue squamous cell carcinoma cells. Biochem. J 2010, 432, 199–205. [Google Scholar]
- Zhao, X.; Dou, W.; He, L.; Liang, S.; Tie, J.; Liu, C.; Li, T.; Lu, Y.; Mo, P.; Shi, Y.; et al. MicroRNA-7 functions as an anti-metastatic microRNA in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene 2013, 32, 1363–1372. [Google Scholar]
- Huang, M.B.; Xu, H.; Xie, S.J.; Zhou, H.; Qu, L.H. Insulin-like growth factor-1 receptor is regulated by microRNA-133 during skeletal myogenesis. PLoS One 2011, 6, e29173. [Google Scholar]
- Song, L.; Duan, P.; Guo, P.; Li, D.; Li, S.; Xu, Y.; Zhou, Q. Downregulation of miR-223 and miR-153 mediates mechanical stretch-stimulated proliferation of venous smooth muscle cells via activation of the insulin-like growth factor-1 receptor. Arch. Biochem. Biophys 2012, 528, 204–211. [Google Scholar]
- Knezevic, I.; Patel, A.; Sundaresan, N.R.; Gupta, M.P.; Solaro, R.J.; Nagalingam, R.S.; Gupta, M. A novel cardiomyocyte-enriched microRNA, miR-378, targets insulin-like growth factor 1 receptor: Implications in postnatal cardiac remodeling and cell survival. J. Biol. Chem 2012, 287, 12913–12926. [Google Scholar]
- Shen, K.; Liang, Q.; Xu, K.; Cui, D.; Jiang, L.; Yin, P.; Lu, Y.; Li, Q.; Liu, J. MiR-139 inhibits invasion and metastasis of colorectal cancer by targeting the type I insulin-like growth factor receptor. Biochem. Pharmacol 2012, 84, 320–330. [Google Scholar]
- La Rocca, G.; Badin, M.; Shi, B.; Xu, S.Q.; Deangelis, T.; Sepp-Lorenzinoi, L.; Baserga, R. Mechanism of growth inhibition by MicroRNA 145: The role of the IGF-I receptor signaling pathway. J. Cell. Physiol 2009, 220, 485–491. [Google Scholar]
- Gusscott, S.; Kuchenbauer, F.; Humphries, R.K.; Weng, A.P. Notch-mediated repression of miR-223 contributes to IGF1R regulation in T-ALL. Leuk. Res 2012, 36, 905–911. [Google Scholar]
- Kong, K.L.; Kwong, D.L.; Chan, T.H.; Law, S.Y.; Chen, L.; Li, Y.; Qin, Y.R.; Guan, X.Y. MicroRNA-375 inhibits tumour growth and metastasis in oesophageal squamous cell carcinoma through repressing insulin-like growth factor 1 receptor. Gut 2012, 61, 33–42. [Google Scholar]
- Zehavi, L.; Avraham, R.; Barzilai, A.; Bar-Ilan, D.; Navon, R.; Sidi, Y.; Avni, D.; Leibowitz-Amit, R. Silencing of a large microRNA cluster on human chromosome 14q32 in melanoma: Biological effects of mir-376a and mir-376c on insulin growth factor 1 receptor. Mol. Cancer 2012, 11, 44, :1–44:15.. [Google Scholar]
- Liang, R.; Khanna, A.; Muthusamy, S.; Li, N.; Sarojini, H.; Kopchick, J.J.; Masternak, M.M.; Bartke, A.; Wang, E. Post-transcriptional regulation of IGF1R by key microRNAs in long-lived mutant mice. Aging Cell 2011, 10, 1080–1088. [Google Scholar]
- Farhana, L.; Dawson, M.I.; Murshed, F.; Das, J.K.; Rishi, A.K.; Fontana, J.A. Upregulation of miR-150* and miR-630 induces apoptosis in pancreatic cancer cells by targeting IGF-1R. PLoS One 2013, 8, e61015. [Google Scholar]
- Santoro, F.; Pauler, F.M. Silencing by the imprinted Airn macro lncRNA: Transcription is the answer. Cell Cycle 2013, 12, 711–712. [Google Scholar]
- Wang, E.T.; Cody, N.A.; Jog, S.; Biancolella, M.; Wang, T.T.; Treacy, D.J.; Luo, S.; Schroth, G.P.; Housman, D.E.; Reddy, S.; et al. Transcriptome-wide regulation of pre-mRNA splicing and mRNA localization by muscleblind proteins. Cell 2012, 150, 710–724. [Google Scholar]
- Pascale, A.; Amadio, M.; Scapagnini, G.; Lanni, C.; Racchi, M.; Provenzani, A.; Govoni, S.; Alkon, D.L.; Quattrone, A. Neuronal ELAV proteins enhance mRNA stability by a PKCalpha-dependent pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 12065–12070. [Google Scholar]
- Bonifacio, L.N.; Jarstfer, M.B. MiRNA profile associated with replicative senescence, extended cell culture, and ectopic telomerase expression in human foreskin fibroblasts. PLoS One 2010, 5, e12519. [Google Scholar]
- Levy, J.R.; Hug, V. Regulation of insulin receptor gene expression. Cell cycle-mediated effects on insulin receptor mRNA stability. J. Biol. Chem 1992, 267, 25289–25295. [Google Scholar]
- Levy, J.R.; Hannah, S.; Mooney, R.L.; Hug, V.; Stevens, W. Sequence and functional characterization of the terminal exon of the human insulin receptor gene. Biochim. Biophys. Acta 1995, 1263, 253–257. [Google Scholar]
- Mukherjee, N.; Corcoran, D.L.; Nusbaum, J.D.; Reid, D.W.; Georgiev, S.; Hafner, M.; Ascano, M., Jr; Tuschl, T.; Ohler, U.; Keene, J.D. Integrative regulatory mapping indicates that the RNA-binding protein HuR couples pre-mRNA processing and mRNA stability. Mol. Cell 2011, 43, 327–339. [Google Scholar]
- Hafner, M.; Max, K.E.; Bandaru, P.; Morozov, P.; Gerstberger, S.; Brown, M.; Molina, H.; Tuschl, T. Identification of mRNAs bound and regulated by human LIN28 proteins and molecular requirements for RNA recognition. RNA 2013, 19, 613–626. [Google Scholar]
- Ascano, M., Jr; Mukherjee, N.; Bandaru, P.; Miller, J.B.; Nusbaum, J.D.; Corcoran, D.L.; Langlois, C.; Munschauer, M.; Dewell, S.; Hafner, M.; et al. Nature 2012, 492, 382–386.
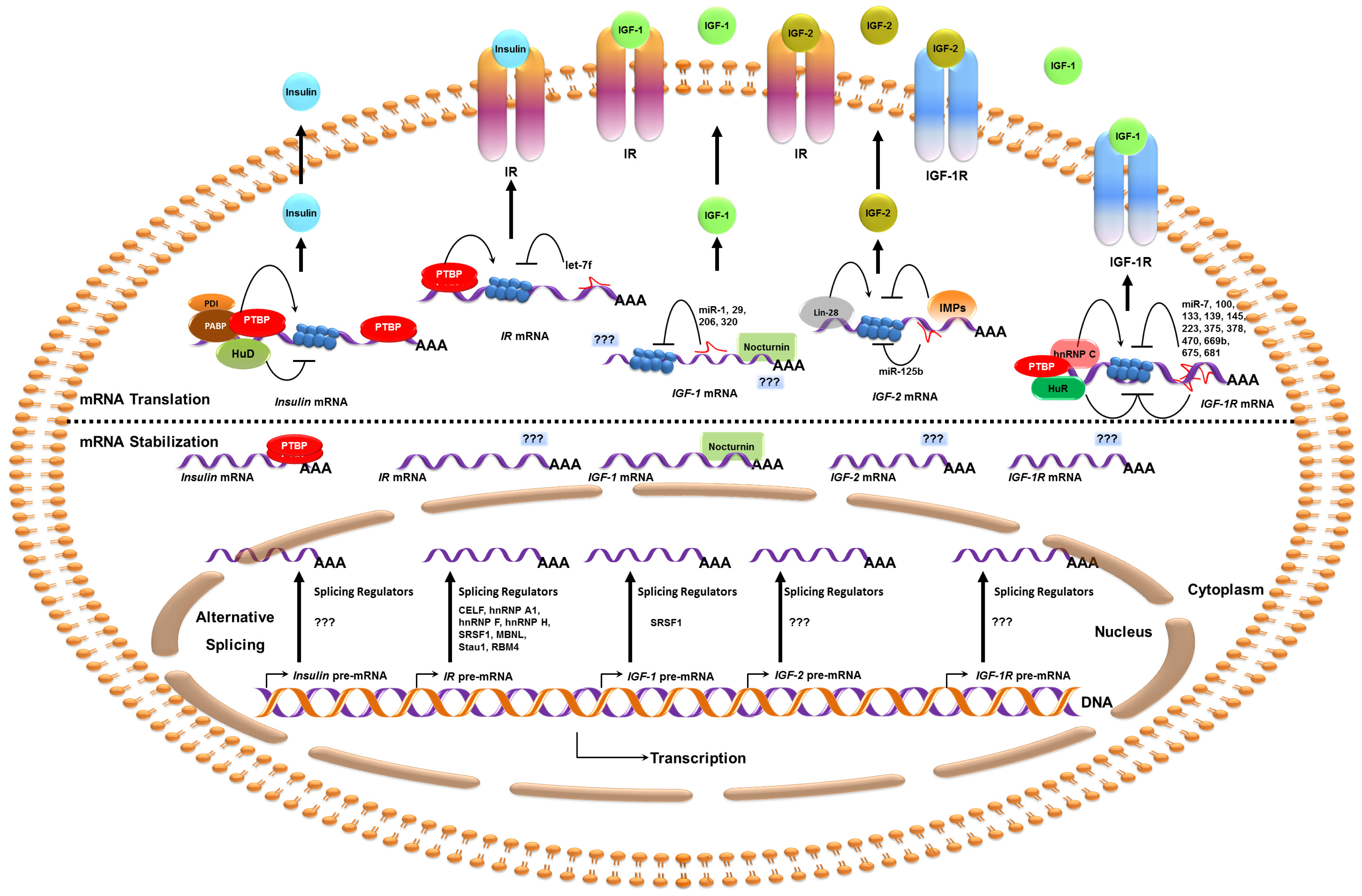

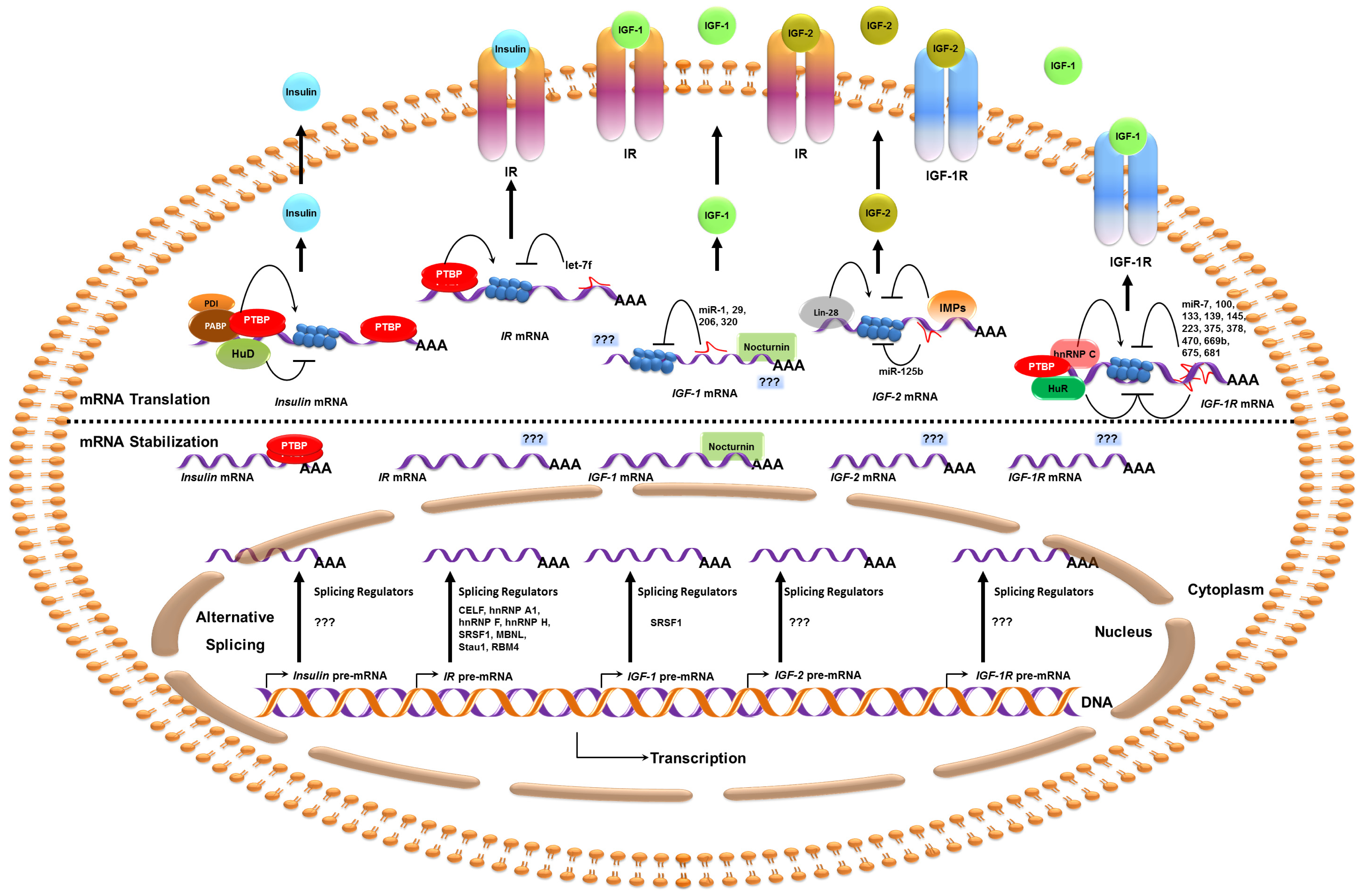{kind=link}
| Target | Regulators | Target Region(s) | Functions | Condition/Disease | Reference |
|---|---|---|---|---|---|
| Insulin | PABP | 5′ UTR | Translation activation | High glucose | [39] |
| PDI | 5′ UTR | Translation activation | High glucose | [39] | |
| HuD | 5′ UTR | Translation repression | Low glucose | [43] | |
| PTBP | 3′ UTR | mRNA stabilization | Hypoxia and High glucose | [13,46] | |
| PTBP | 5′ UTR | IRES translation activation | Nitrosative stress | [47] | |
| IR | CELF1 | Intron 10, Exon 11 | Exon 11 skipping | Muscular Dystrophy | [63,66] |
| SRp20 | Exon 11 | Exon 11 inclusion | Basal | [63] | |
| SRSF1 | Exon 11 | Exon 11 inclusion | Basal | [63,68] | |
| hnRNP H | Intron 10 | Exon 11 skipping | Muscular Dystrophy | [66] | |
| hnRNP A1 | Intron 10, Exon 11 | Exon 11 skipping | Muscular Dystrophy | [68] | |
| hnRNP F | Intron 10 | Exon 11 inclusion | Basal | [68] | |
| MBNL | Intron 10, 11 and exon 11 | Exon 11 inclusion | Basal | [63,66,71] | |
| RBM4 | NA | Exon 11 inclusion | High glucose | [76] | |
| PTBP1, PTBP2 | 5′ UTR | IRES translation activation | Insulin stimulus | [79] | |
| LIN-28 | CR and 3′ UTR | NA | Basal | [151] | |
| FMRP | CR and 3′ UTR | NA | Basal | [152] | |
| let-7f | 3′ UTR | Translation Repression * | Diabetes | [21] | |
| IGF-1 | SRSF1 | Exon 5 | Exon 5 inclusion | Growth hormone treatment | [103] |
| Nocturnin | 3′ UTR | mRNA decay | Circadian rhythm | [108] | |
| miR-29 | 3′ UTR | mRNA decay | Fibrosis | [128] | |
| miR-1 | 3′ UTR | Translation repression | Muscle differentiation | [22] | |
| miR-1 | 3′ UTR | Translation repression | Apoptosis | [129,130] | |
| miR-206 | 3′ UTR | Translation repression | Apoptosis | [130] | |
| miR-320 | 3′ UTR | Translation repression * | Angiogenesis | [131] | |
| IGF-2 | Lin28 | NA | Translation activation | Skeletal myogenesis | [16] |
| IMP1, IMP2, IMP3 | 5′ UTR (L3) | Translation repression | Embryogenesis | [121] | |
| IMP3 | 5′ UTR (L3 and L4) | Translation activation | Cell proliferation | [122] | |
| IMP2 | 5′ UTR (L3) | IRES translation activation | Embryogenesis | [17] | |
| miR-125b | 3′ UTR | mRNA decay | Muscle regeneration | [23] | |
| IGF-1R | HuR | 5′ UTR | Translation repression | Amino acid deprivation | [15] |
| HuR | 5′ UTR | IRES translation repression | Amino acid deprivation | [15] | |
| hnRNP C | 5′ UTR | IRES translation activation | Metaphase block | [115] | |
| PTBP | 5′ UTR | IRES translation activation | Basal | [119] | |
| LIN-28 | Intron, CR and 3′ UTR | NA | Basal | [151] | |
| FMRP | Intron, CR and 3′ UTR | NA | Basal | [152] | |
| miR-7 | 3′ UTR | mRNA decay | Apoptosis | [132] | |
| miR-7 | 3′ UTR | Translation repression | Metastasis | [133] | |
| miR-100 | 3′ UTR | Translation repression * | Adrenocortical tumors | [24] | |
| miR-133 | 3′ UTR | Translation repression | Muscle differentiation | [134] | |
| miR-145 | 3′ UTR | Translation repression | Colon cancer | [138] | |
| miR-223 | 3′ UTR | Translation repression | Muscle differentiation | [135] | |
| miR-223 | 3′ UTR | Translation repression | Lukemia | [139] | |
| miR-375 | 3′ UTR | mRNA decay | Cancer | [140] | |
| miR-378 | 3′ UTR | Translation repression | Cardiomyocyte survival | [136] | |
| miR-675-3p | 3′ UTR | mRNA decay | Placental growth | [26] | |
| miR-139 | 3′ UTR | mRNA decay | Colorectal cancer | [137] | |
| miR-376a and 376c | 3′ UTR | mRNA decay | Melanoma | [141] | |
| miR-470, 669b and 681 | 3′ UTR | Translation repression * | GH-deficiency | [142] | |
| miR-150* and 630 | 3′ UTR | mRNA decay | Growth arrest and apoptosis | [143] | |
| IGF-2R | HuR | Intron and 3′ UTR | NA | Basal | [150] |
| LIN-28 | Intron and CR | NA | Basal | [151] | |
| FMRP | CR | NA | Basal | [152] | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Panda, A.C.; Grammatikakis, I.; Yoon, J.-H.; Abdelmohsen, K. Posttranscriptional Regulation of Insulin Family Ligands and Receptors. Int. J. Mol. Sci. 2013, 14, 19202-19229. https://doi.org/10.3390/ijms140919202
Panda AC, Grammatikakis I, Yoon J-H, Abdelmohsen K. Posttranscriptional Regulation of Insulin Family Ligands and Receptors. International Journal of Molecular Sciences. 2013; 14(9):19202-19229. https://doi.org/10.3390/ijms140919202
Chicago/Turabian StylePanda, Amaresh C., Ioannis Grammatikakis, Je-Hyun Yoon, and Kotb Abdelmohsen. 2013. "Posttranscriptional Regulation of Insulin Family Ligands and Receptors" International Journal of Molecular Sciences 14, no. 9: 19202-19229. https://doi.org/10.3390/ijms140919202
APA StylePanda, A. C., Grammatikakis, I., Yoon, J.-H., & Abdelmohsen, K. (2013). Posttranscriptional Regulation of Insulin Family Ligands and Receptors. International Journal of Molecular Sciences, 14(9), 19202-19229. https://doi.org/10.3390/ijms140919202