Biochemistry and Molecular Biology of Carotenoid Biosynthesis in Chili Peppers (Capsicum spp.)

Abstract
:1. Introduction
2. Capsicum Fruit Carotenoids
3. Carotenoid Content and Composition during Chili Pepper Fruit Ripening
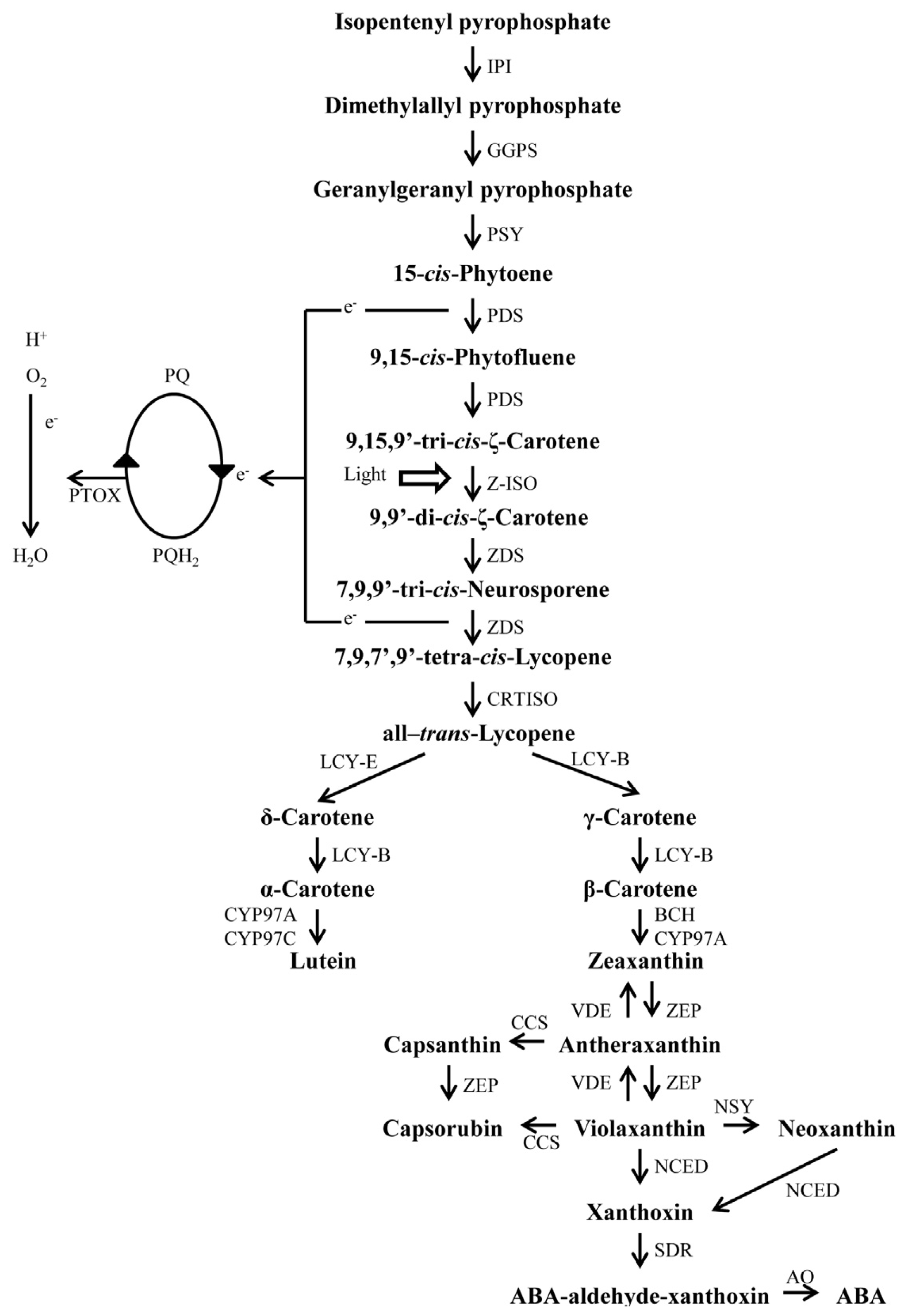
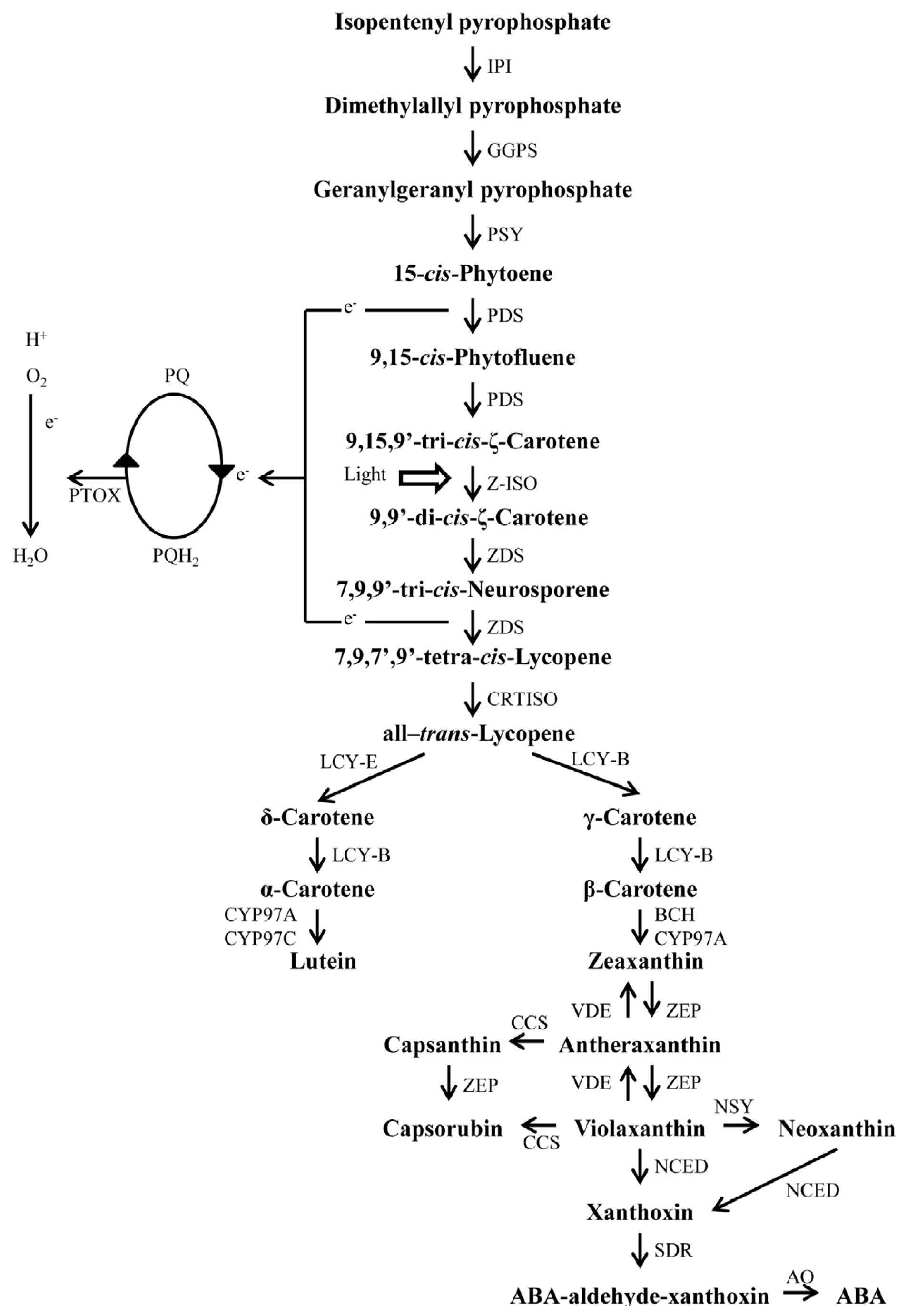
4. Carotenoid Biosynthesis Pathway in Plants
5. Carotenoid Biosynthesis in Chili Pepper Fruits
6. Regulation of Carotenoid Biosynthesis in Capsicum
7. Future Research on Chili Pepper Carotenoid Biosynthesis
8. Plausible Applications
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Botella-Pavía, P.; Rodríguez-Concepción, M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol. Plant 2006, 126, 369–381. [Google Scholar]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch. Biochem. Biophys 2009, 483, 191–195. [Google Scholar]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Progr. Lipid Res 2004, 43, 228–265. [Google Scholar]
- Delgado-Vargas, F.; Peredes-López, O. Natural Colorants for Food and Nutraceutical Uses; CRC Press: Boca Raton, FL, USA, 2003; p. 327. [Google Scholar]
- Hadden, W.L.; Watkins, R.H.; Levy, L.W.; Regalado, E. Carotenoid composition of Marigold (Tagetes erecta) flower extract used as nutritional supplement. J. Agric. Food Chem 1999, 47, 4189–4194. [Google Scholar]
- Jordan, B.R. The Molecular Biology and Biotechnology of Flowering, 2nd ed; CABI Publishing: Cambridge, MA, USA, 2006; p. 404. [Google Scholar]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P. Carotenoid biosynthesis during tomato fruit development. Plant Physiol 1994, 105, 405–413. [Google Scholar]
- Skelton, R.L.; Yu, Q.; Srinivasan, R.; Manshardt, R.; Moore, P.H.; Ray, M. Tissue differential expression of lycopene β-cyclase gene in papaya. Cell Res 2006, 16, 731–739. [Google Scholar]
- Kato, M.; Ikoma, Y.; Matsumoto, H.; Sugiura, M.; Hyodo, H.; Yano, M. Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit. Plant Physiol 2004, 134, 824–837. [Google Scholar]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar]
- Weedon, B.C.L.; Moss, G.P. Structure and Nomenclature. In Carotenoids; Britton, G., Liaaen-Jensen, S., Pfander, H.P., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; Volume I B, Spectroscopy; pp. 27–70. [Google Scholar]
- Sandmann, G.; Albrecht, M.; Schnurr, G.; Knörzer, O.; Böger, P. The biotechnological potential and design of novel carotenoids by gene combination in Escherichia coli. Trends Biotechnol 1999, 17, 233–236. [Google Scholar]
- Zhu, C.; Naqvi, S.; Capell, T.; Christou, P. Metabolic engineering of ketocarotenoid biosynthesis in higher plants. Arch. Biochem. Biophys 2009, 483, 182–190. [Google Scholar]
- Auldridge, M.E.; McCarthy, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Curr. Opin. Plant Biol 2006, 9, 315–321. [Google Scholar]
- Walter, M.H.; Floss, D.S.; Strack, D. Apocarotenoids: Hormones, mycorrhizal metabolies and aroma volatiles. Planta 2010, 232, 1–17. [Google Scholar]
- Wang, R.K.; Lu, J.J.; Xing, G.N.; Gai, J.Y.; Zhao, T.J. Molecular evolution of two consecutive carotenoid cleavage dioxygenase genes in strigolactone biosynthesis in plants. Genet. Mol. Res 2011, 10, 3664–3673. [Google Scholar]
- Sharoni, Y.; Linnewiel-Hermoni, K.; Khanin, M.; Salman, H.; Veprik, A.; Danilenko, M.; Levy, J. Carotenoids and apocarotenoids in cellular signaling related to cancer: A review. Mol. Nutr. Food Res 2012, 56, 259–269. [Google Scholar]
- Li, L.; van Eck, J. Metabolic engineering of carotenoid accumulation by creating a metabolic sink. Transgenic Res 2007, 16, 581–585. [Google Scholar]
- Nishino, H.; Tokuda, H.; Satomi, Y.; Masuda, M.; Bu, P.; Onozuka, M.; Yamaguchi, S.; Okuda, Y.; Takayasu, J.; Tsuruta, J.; et al. Cancer prevention by carotenoids. Pure Appl. Chem 1999, 71, 2273–2278. [Google Scholar]
- Sporn, M.B.; Liby, K.T. Is lycopene an effective agent for preventing prostate cancer? Cancer Prev. Res 2013, 6, 384–386. [Google Scholar]
- Snodderly, D.M. Evidence for protection against age-related macular degeneration by carotenoids and antioxidant vitamins. Am. J. Clin. Nutr 1995, 62, 1448S–1461S. [Google Scholar]
- Rodríguez-Concepción, M.; Forés, O.; Martinez-Garcia, J.F.; Gonzalez, V.; Phillips, M.A.; Ferrer, A.; Boronat, A. Distinct light-mediated pathways regulate the biosynthesis and exchange of isoprenoid precursors during Arabidopsis seedling development. Plant Cell 2004, 16, 144–156. [Google Scholar]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K.; Akimoto, N. Isolation of a series of apocarotenoids from the fruits of the red paprika Capsicum annuum L. J. Agric. Food Chem 2001, 49, 1601–1606. [Google Scholar]
- Braconnot, H. Mémoire sur le principe extractif et sur les extraits en général. J. Phys. Chim. Hist. Nat. Arts 1817, 84, 267–296. [Google Scholar]
- Ikan, R. Natural Products: A Laboratory Guide, 2nd ed; Academic Press: San Diego, CA, USA, 1991; p. 360. [Google Scholar]
- Von Zechmeister, L.; von Cholnoky, L. Untersuchungen über den paprika-farbstoff. XI. Isomerisierungs-Erscheinungen. Ann. Chem 1940, 543, 248–257. [Google Scholar]
- Curl, A.L. The carotenoids of red bell peppers. J. Agric. Food Chem 1962, 10, 504–509. [Google Scholar]
- Von Cholnoky, L.; Györgyfy, K.; Nagy, E.; Pánczél, M. Investigations of carotenoid pigments. I. Pigments of red paprika. Acta Chim. Acad. Sci. Hung 1955, 6, 143–147. [Google Scholar]
- Barber, M.S.; Jackman, L.M.; Warren, C.K.; Weedon, B.C.L. The structures of the paprika ketones. Proc. Chem. Soc. 1960, 19–20. [Google Scholar]
- Entschel, R.; Karrer, P. Zur Konstitution des capsanthins und capsorubins. Helv. Chim. Acta 1960, 43, 89–94. [Google Scholar]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem 2000, 48, 1713–1720. [Google Scholar]
- Marín, A.; Ferreres, F.; Tómas-Barberán, F.A.; Gill, M.I. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.). J. Agric. Food Chem 2004, 52, 3861–3869. [Google Scholar]
- Sun, T.; Xu, Z.; Wu, C.T.; Janes, M.; Prinyawiwatkul, W.; No, H.K. Antioxidant activities of different colored Sweet Bell peppers (Capsicum annuum L.). J. Food Sci 2007, 72, S98–S102. [Google Scholar]
- Karrer, P.; Oswald, A. Carotinoide aus den staubbeuteln von Lilium tigrinum. Ein nenes carotenoid: Anthetaxanthin. Helv. Chim. Acta 1935, 13, 1303–1305. [Google Scholar]
- Seybold, A. Untersuchungen übe den farbwechsel von blumenblättern, früchten un samenschalen. Sber. Heilb. Akad. Wiss. Math-Naturwiss Kl 1953, 4, 31–124. [Google Scholar]
- Valadon, L.R.G.; Mummery, R.S. Carotenoids of lilies and of red pepper: Biogenesis of capsanthin and capsorubin. Z. Pflanzephysiol 1976, 82, 407–416. [Google Scholar]
- Rüttimann, A. Synthesis and Stereochemistry of Red Pepper Carotenoids. In Carotenoid Chemistry and Biochemistry; Britton, G., Goodwin, T.W., Eds.; Pergamon Press: New York, NY, USA, 1982; pp. 71–86. [Google Scholar]
- Märki-Fischer, E.; Eugster, C.H. Das carotinoidspektrum der antheren un petalen von Lilium tigrinum cv. “Red Night”. Helv. Chim. Acta 1985, 68, 1708–1715. [Google Scholar]
- Partali, V.; Liaaen-Jensen, S.; Huneck, S.; Khaidav, T. Carotenoids from the flowers of Lilium pumilum. Die Pharm 1987, 42, 208. [Google Scholar]
- Deli, J.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of Asparagus officinalis. J. Agric. Food Chem 2000, 48, 2793–2796. [Google Scholar]
- Yamagishi, M.; Kishimoto, S.; Nakayama, M. Carotenoid composition and changes in expression of carotenoid biosynthetic genes in tepals of Asiatic hybrid lily. Plant Breed 2010, 129, 100–107. [Google Scholar]
- Jeknic, Z.; Moré, J.T.; Jeknic, S.; Jevremovic, S.; Subotic, A.; Chen, T.H.H. Cloning and functional characterization of a gene for capsanthin-capsorubin synthase from tiger lily (Lolium lancifolium Thunb. “Splendens”). Plant Cell Physiol 2012, 53, 1899–1912. [Google Scholar]
- Matsufuji, H.; Nakamura, H.; Chino, M.; Takeda, M. Antioxidant activity of capsanthin and the fatty acid esters in paprika (Capsicum annuum). J. Agric. Food Chem 1998, 46, 3468–3472. [Google Scholar]
- Kim, S.; Ha, T.Y.; Hwang, I.K. Analysis, bioavailability, and potential healthy effects of capsanthin, natural red pigment from Capsicum spp. Food Rev. Int 2009, 25, 198–213. [Google Scholar]
- Deruère, J.; Römer, S.; d’Harlingue, A.; Backhaus, R.A.; Kuntz, M.; Camara, B. Fibril assembly and carotenoid overaccumulation in chromoplasts: A model for supramolecular lipoprotein structures. Plant Cell 1994, 6, 119–133. [Google Scholar]
- Mínguez-Mosquera, M.I.; Hornero-Méndez, D. Formation and transformation of pigments during the fruit ripening of Capsicum annuum cv. Bola and Agridulce. J. Agric. Food Chem 1994, 42, 38–44. [Google Scholar]
- Rodríguez-Burruezo, A.; González-Mas, M.C.; Nuez, F. Carotenoid composition and vitamin A value in ají (Capsicum baccatum L.) and rocoto (C. pubescens R. & P.), 2 pepper species from the Andean region. J. Food Sci 2010, 75, S446–S453. [Google Scholar]
- Deli, J.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of Capsicum annuum cv. Szentesi Kosszarvú during ripening. J. Agric. Food Chem 1996, 44, 711–716. [Google Scholar]
- Mínguez-Mosquera, M.I.; Hornero-Méndez, D. Separation and quantification of the carotenoid pigments in red peppers (Capsicum annuum L.), paprika and oleoresin by reversed-phase HPLC. J. Agric. Food Chem 1993, 41, 1616–1620. [Google Scholar]
- Hornero-Méndez, D.; Gómez-Ladrón de Guevara, R.; Mínguez-Mosquera, M.I. Carotenoid biosynthesis changes in five red pepper (Capsicum annuum L.) cultivars during ripening. Cultivar selection for breeding. J. Agric. Food Chem 2000, 48, 3857–3864. [Google Scholar]
- Mínguez-Mosquera, M.I.; Hornero-Méndez, D. Changes in carotenoid esterification during the fruit ripening of Capsicum annuum cv. Bola. J. Agric. Food Chem 1994, 42, 640–644. [Google Scholar]
- Curl, A.L. The carotenoids of green bell peppers. J. Agric. Food Chem 1964, 12, 522–524. [Google Scholar]
- Matus, Z.; Deli, J.; Szabolcs, J. Carotenoid composition of yellow pepper during ripening: Isolation of β-cryptoxanthin 5,6-epoxide. J. Agric. Food Chem 1991, 39, 1907–1914. [Google Scholar]
- Deli, J.; Matus, Z.; Szabolcs, J. Carotenoid composition in the fruits of black paprika (Capsicum annuum variety longum nigrum) during ripening. J. Agric. Food Chem 1992, 40, 2072–2076. [Google Scholar]
- Deli, J.; Molnár, P.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of red paprika (Capsicum annuum var. lycopersiciforme rubrum) during ripening; Biosynthesis of carotenoids in red paprika. J. Agric. Food Chem 2001, 49, 1517–1523. [Google Scholar]
- Collera-Zuñiga, O.; García, J.F.; Meléndez, G.R. Comparative study of carotenoid composition in three mexican varieties of Capsicum annuum L. Food Chem 2005, 90, 109–114. [Google Scholar]
- Matsufuji, H.; Ishikawa, K.; Nunomura, O.; Chino, M.; Takeda, M. Anti-oxidant content of different coloured sweet peppers, white, green, yellow, orange and red (Capsicum annuum L.). Int. J. Food Sci. Technol 2007, 42, 1482–1488. [Google Scholar]
- Roca, M.; Hornero-Méndez, D.; Gandul-Rojas, B.; Mínguez-Mosquera, M.I. Stay-green phenotype slows the carotenogenic process in Capsicum annuum (L.) fruits. J. Agric. Food Chem 2006, 54, 8782–8787. [Google Scholar]
- Almela, L.; Fernández-López, J.A.; Candela, M.E.; Egea, C.; Alcázar, M.D. Changes in pigments, chlorophyllase activity, and chloroplast ultrastructure in ripening pepper for paprika. J. Agric. Food Chem 1996, 44, 1704–1711. [Google Scholar]
- Camara, B.; Brangeon, J. Carotenoid metabolism during chloroplast to chromoplast transformation in Capsicum annuum fruit. Planta 1981, 151, 359–364. [Google Scholar]
- Guzman, I.; Hamby, S.; Romero, J.; Bosland, P.W.; O’Connell, M.A. Variability of carotenoid biosynthesis in orange colored Capsicum spp. Plant Sci. 2010, 179, 49–59. [Google Scholar]
- Deepa, N.; Kaur, C.; George, B.; Singh, B.; Kapoor, H.C. Antioxidant constituents in some sweet pepper (Capsicum annuum L.) genotypes during maturity. LWT-Food Sci. Technol 2007, 40, 121–129. [Google Scholar]
- Maoka, T.; Akimoto, N.; Fujiwara, Y.; Hashimoto, K. Structure of new carotenoids with the 6-oxo-κ end group from the fruits of paprika, Capsicum annuum. J. Nat. Prod 2004, 67, 115–117. [Google Scholar]
- Lichtenthaler, H.K. The 1-deoxy-d-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 47–65. [Google Scholar]
- Eisenreich, W.; Rohdich, F.; Bacher, A. Deoxyxylulose phosphate pathway to terpenoids. Trends Plant Sci 2001, 6, 78–84. [Google Scholar]
- Rodríguez-Concepción, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol 2002, 130, 1079–1089. [Google Scholar]
- Dogbo, O.; Camara, B. Purification of isopentenyl pyrophosphate isomerase and geranylgeranyl pyrophosphate synthase from Capsicum chromoplasts by affinity chromatography. Biochim. Biophys. Acta 1987, 920, 140–148. [Google Scholar]
- Kuntz, M.; Römer, S.; Suire, C.; Hugueney, P.; Weil, J.H.; Schantz, R.; Camara, B. Identification of a cDNA for the plastid-located geranylgeranyl pyrophosphate synthase from Capsicum annuum: Correlative increase in enzyme activity and transcript level during fruit ripening. Plant J 1992, 2, 25–34. [Google Scholar]
- Carol, P.; Stevenson, D.; Bisanz, C.; Breitenbach, J.; Sandman, G.; Mache, R.; Coupland, G.; Kuntz, M. Mutations in the Arabidopsis gene immutants cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell 1999, 11, 57–68. [Google Scholar]
- Carol, P.; Kuntz, M. A plastid terminal oxidase comes to light: Implications for carotenoid biosynthesis and chlororespiration. Trends Plant Sci 2001, 6, 31–36. [Google Scholar]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci 2002, 7, 41–48. [Google Scholar]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A.D. Elucidation of the indirect pathway of abscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol 2003, 131, 1591–1601. [Google Scholar]
- Aluru, M.R.; Rodermel, S.R. Control of chloroplast redox by IMMUTANTS terminal oxidase. Physiol. Plant 2004, 120, 4–11. [Google Scholar]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol 2008, 50, 778–785. [Google Scholar]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage productos: Biosynthesis and functions. Nat. Prod. Rep 2011, 28, 663–692. [Google Scholar]
- Li, Q.; Farre, G.; Naqvi, S.; Breitenbach, J.; Sanahuja, G.; Bai, C.; Sandmann, G.; Capel, T.; Christou, P.; Zhu, C. Cloning and functional charactarization of the maize carotenoid isomerase and β-carotene hydroxylase genes and their regulation during endosperm maturation. Plant Physiol. 2010, 19, 1053–1068. [Google Scholar]
- Zhu, C.; Sanahuja, G.; Yuan, D.; Farré, G.; Arjó, G.; Berman, J.; Zorrilla-López, U.; Banakar, R.; Bai, C.; Pérez-Massot, E.; et al. Biofortification of plants with altered antioxidant content and composition: Genetic engineering strategies. Plant Biotechnol. J 2013, 11, 129–141. [Google Scholar]
- Dogbo, O.; Bardat, F.; Laferriere, A.; Quennement, J.; Brangeon, J.; Camara, B. Metabolism of plastid terpenoids. I. Biosynthesis in plastid stroma isolated from higher plants. Plant Sci 1987, 49, 89–101. [Google Scholar]
- Dogbo, O.; Laferriere, A.D.; d’Harlingue, A.; Camara, B. Carotenoid biosynthesis; Isolation of a bifunctional enzyme catalyzing the synthesis of phytoene. Proc. Natl. Acad. Sci. USA 1988, 85, 7054–7058. [Google Scholar]
- Cunningham, F.X.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol 1998, 49, 557–583. [Google Scholar]
- Hugueney, P.; Römer, S.; Kuntz, M.; Camara, B. Characterization and molecular cloning of a flavoprotein catalyzing the synthesis of phytofluene and ζ-carotene in Capsicum chromoplasts. Eur. J. Biochem 1992, 209, 399–407. [Google Scholar]
- Li, F.; Murillo, C.; Wurtzel, E.T. Maize Y9 encodes a product essential for 15-cis zetacarotene isomerization. Plant Physiol 2007, 144, 1181–1189. [Google Scholar]
- Chen, Y.; Li, F.; Wurtzel, E.T. Isolation and characterization of the Z-ISO gene encoding a missing component of carotenoid biosynthesis in plants. Plant Physiol 2010, 153, 66–79. [Google Scholar]
- Albrecht, M.; Klein, A.; Hugueney, P.; Sandmann, G.; Kuntz, M. Molecular cloning and functional expression in E. coli of a novel plant enzyme mediating zeta-carotene desaturation. FEBS Lett 1995, 372, 199–202. [Google Scholar]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato revels a carotenoid isomerase essential for the production of beta-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar]
- Park, H.; Kreunen, S.S.; Cuttriss, A.J.; DellaPenna, D.; Pogson, B.J. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis. Plant Cell 2002, 14, 321–332. [Google Scholar]
- Scolnik, P.A.; Hinton, P.; Greenblatt, I.M.; Giuliano, G.; Delanoy, M.R.; Spector, D.L.; Pollock, D. Somatic instability of carotenoid biosynthesis in the tomato ghost mutant and its effect on plastid development. Planta 1987, 171, 11–18. [Google Scholar]
- Wetzel, C.M.; Jiang, C.-Z.; Meehan, L.J.; Voytas, D.F.; Ridermel, S.R. Nuclear-organelle interactions: The immutants variegation mutant of Arabidopsis is plastid autonomous and impaired in carotenoid biosynthesis. Plant J 1994, 6, 161–175. [Google Scholar]
- Wu, D.; Wright, D.A.; Wetzel, C.; Voytas, D.F.; Rodermel, S. The immutants variegation locus of Arabidopsis defines a mitochondrial alternative oxidase homolog that functions during early choroplasts biogenesis. Plant Cell 1999, 11, 43–55. [Google Scholar]
- Shahbazi, M.; Gilbert, M.; Mabouré, A.-M.; Kuntz, M. Dual role of the plastid terminal oxidase in tomato. Plant Physiol 2007, 145, 691–702. [Google Scholar]
- Rumeau, D.; Peltier, G.; Cournac, L. Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response. Plant Cell Environ 2007, 30, 1041–1051. [Google Scholar]
- Parker, R.S.; Swanson, J.E.; You, C.S.; Edwards, A.J.; Huang, T. Bioavailability of carotenoids in human subjects. Proc. Nutr. Soc 1999, 58, 155–162. [Google Scholar]
- During, A.; Hussain, M.M.; Morel, D.W.; Harrison, E.H. Carotenoid uptake and secretion by CaCo-2 cells: β-carotene isomer selectivity and carotenoid interactions. J. Lip. Res 2002, 43, 1086–1095. [Google Scholar]
- Quinlan, R.F.; Shumskaya, M.; Bradbury, L.M.T.; Beltrán, J.; Ma, C.; Kennelly, E.J.; Wurtzel, E.T. Synergistic interactions between carotene ring hydroxylases drive lutein formation in plant carotenoid biosynthesis. Plant Physiol. 2012, 160, 204–214. [Google Scholar]
- Demming-Adams, B.; Gilmore, A.M.; Adams, W.W. In vivo functions of carotenoids in higher plants. FASEB J 1996, 10, 403–412. [Google Scholar]
- Camara, B.; Bardat, F.; Monéger, R. Sites of biosynthesis of carotenoids in Capsicum chromoplasts. Eur. J. Biochem 1982, 127, 255–258. [Google Scholar]
- Camara, B.; Bardat, F.; Dogbo, O.; Brangeon, J.; Monéger, R. Terpenoid mtabolism in plastids. Isolation and biochemical characteristics of Capsicum annuum chromoplasts. Plant Physiol 1983, 73, 94–99. [Google Scholar]
- Camara, B. Biosynthesis of keto-carotenoids in Capsicum Annuum fruits. FEBS Lett 1980, 118, 315–318. [Google Scholar]
- Camara, B.; Monéger, R. Carotenoid biosynthesis. In vitro conversion of antheraxanthin to capsanthin by a chromoplast enriched fraction of Capsicum fruits. Biochem. Biophys. Res. Commun 1981, 99, 1117–1122. [Google Scholar]
- Camara, B. Prenylation of chlorophyllide a in Capsicum plastids. Methods Enzymol 1985, 110, 274–281. [Google Scholar]
- Bouvier, F.; Hugueney, P.; d’Harlingue, A.; Kuntz, M.; Camara, B. Xanthophyll biosynthesis in chromoplasts: Isolation and molecular cloning of an enzyme catalyzing the conversion of 5,6-epoxycarotenoid into ketocarotenoid. Plant J 1994, 6, 45–54. [Google Scholar]
- Bouvier, F.; d’Harlingue, A.; Hugueney, P.; Marin, E.; Marion-Poll, A.; Camara, B. Xanthophyll biosynthesis. J. Biol. Chem 1996, 271, 28861–28867. [Google Scholar]
- Badillo, A.; Steppuhn, J.; Deruère, J.; Camara, B.; Kuntz, M. Structure of a functional geranylgeranyl pyrophosphate synthase gene from Capsicum annuum. Plant Mol. Biol 1995, 27, 425–428. [Google Scholar]
- Bouvier, F.; Backhaus, R.A.; Camara, B. Induction and control of chromoplast-specific carotenoid genes by oxidative stress. J. Biol. Chem 1998, 273, 30651–30659. [Google Scholar]
- Camara, B. Terpenoid metabolism in plastids. Sites of phytoene synthetase activity and synthesis in plant cells. Plant Physiol 1984, 74, 112–116. [Google Scholar]
- Breitenbach, J.; Kuntz, M.; Takaichi, S.; Sandmann, G. Catalytic properties of an expressed and purified higher plant type ζ-carotene desaturase from Capsicum annuum. Eur. J. Biochem. 1999, 265, 376–383. [Google Scholar]
- Josse, E.-M.; Simkin, A.J.; Gaffé, J.; Labouré, A.-M.; Kuntz, M.; Carol, P. A plastid terminal oxidase associated with carotenoid desaturation during chromoplast differentiation. Plant Physiol 2000, 123, 1427–1436. [Google Scholar]
- Camara, B.; Dogbo, O. Demonstration and solubilization of lycopene cyclase from Capsicum chromoplast membranes. Plant Physiol 1986, 80, 172–174. [Google Scholar]
- Hugueney, P.; Badillo, A.; Chen, H.; Klein, A.; Hirschberg, J.; Camara, B.; Kuntz, M. Metabolism of cyclic carotenoids: A model for the alteration of this biosynthetic pathway in Capsicum annuum chromoplasts. Plant J 1995, 8, 417–424. [Google Scholar]
- Simkin, A.J.; Zhu, C.; Kuntz, M.; Sandmann, G. Light-dark regulation of carotenoid biosynthesis in pepper (Capsicum annuum) leaves. J. Plant Physiol 2003, 160, 439–443. [Google Scholar]
- Ha, S.H.; Kim, J.B.; Park, J.S.; Lee, S.W.; Cho, K.J. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot 2007, 58, 3135–3144. [Google Scholar]
- Ronen, G.; Cohen, M.; Zamir, D.; Hirschberg, J. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J 1999, 17, 341–351. [Google Scholar]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to beta-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 1102–1107. [Google Scholar]
- Moehs, C.P.; Tian, L.; Osteryoung, K.W.; DellaPenna, D. Analysis of carotenoids biosynthetic gene expression during marigold petal development. Plant Mol. Biol 2001, 45, 281–293. [Google Scholar]
- Levy, A.; Harel, S.; Palevitch, D.; Akiri, B.; Menagem, E.; Kanner, J. Carotenoid pigments and β-carotene in paprika fruits (Capsicum spp.) with different genotypes. J. Agric. Food Chem 1995, 43, 362–366. [Google Scholar]
- Hurtado-Hernandez, H.; Smith, P.G. Inheritance of mature fruit color in Capsicum annuum L. J. Hered 1985, 76, 211–213. [Google Scholar]
- Lefebvre, V.; Kuntz, M.; Camara, B.; Palloix, A. The capsanthin-capsorubin synthase gene: A candidate gene for the y locus controlling the red fruit colour in pepper. Plant Mol. Biol 1998, 36, 785–789. [Google Scholar]
- Popovsky, S.; Paran, I. Molecular genetics of the y locus in pepper: Its relation to capsanthin-capsorubin synthase and to fruit color. Theor. Appl. Genet 2000, 101, 86–89. [Google Scholar]
- Huh, J.H.; Kang, B.C.; Nahm, S.H.; Kim, S.; Ha, K.S.; Lee, M.H.; Kim, B.D. A candidate gene approach identifies phytoene synthase as the locus for mature fruit color in red pepper (Capsicum spp.). Theor. Appl. Genet 2001, 102, 524–530. [Google Scholar]
- Kim, O.R.; Cho, M.-C.; Kim, B.-D.; Huh, J.H. A splicing mutation in the gene encoding phytoene synthase causes orange coloration in Habanero pepper fruits. Mol. Cells 2010, 30, 569–574. [Google Scholar]
- Borovsky, Y.; Tadmor, Y.; Bar, E.; Meir, A.; Lewinsohn, E.; Paran, I. Induced mutation in β-CAROTENE HYDROXYLASE results in accumulation of β-carotene and conversion of red to orange color in pepper fruits. Theor. Appl. Genet 2013, 126, 557–565. [Google Scholar]
- Lang, Y.Q.; Yanagawa, S.; Sasanuma, T.; Sasakuma, T. Orange fruit color in Capsicum due to deletion of capsanthin-capsorubin synthesis gene. Breed. Sci 2004, 54, 33–39. [Google Scholar]
- Buhler, M.; Mohn, F.; Stalder, L.; Muhlemann, O. Transcriptional silencing of nonsense codon-containing immunoglobulin minigenes. Mol. Cells 2005, 18, 307–317. [Google Scholar]
- Li, Z.; Wang, S.; Gui, X.-L.; Chang, X.-B.; Gong, Z.-H. A further analysis of the relationship between yellow ripe-fruit color and the capsanthin-capsorubin synthase gene in pepper (Capsicum sp.) indicated a new mutant variant in C. annuum and a tandem repeat structure in promoter region. PLoS One 2013, 8, e61996. [Google Scholar]
- Kumagai, M.H.; Keller, Y.; Bouvier, F.; Clary, D.; Camara, B. Functional integration of non-native carotenoids into chloroplasts by viral-derived expression of capsanthin-capsorubin synthase in Nicotiana benthamiana. Plant J 1998, 14, 305–315. [Google Scholar]
- Rodríguez-Uribe, L.; Guzman, I.; Rajapakse, W.; Richins, R.D.; O’Connell, M.A. Carotenoid accumulation in orange-pigmente Capsicum fruit, regulated at multiple levels. J. Exp. Bot. 2012, 63, 517–526. [Google Scholar]
- Welsch, R.; Maass, D.; Voegel, T.; DellaPenna, D.; Beyer, P. Transcription factor RAP2.2 and its interacting partner SINAT2: Stable elements in the carotenogenesis of Arabidopsis leaves. Plant Physiol 2007, 145, 1073–1085. [Google Scholar]
- Toledo-Ortiz, G.; Huq, E.; Rodríguez-Concepción, M. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA 2010, 107, 11626–11631. [Google Scholar]
- Daood, H.G.; Tömöskozi-Farkas, R.; Kapitány, J. Antioxidant content of bio and conventional spice red pepper (Capsicum annuum L.) as determined by HPLC. Acta Agron. Hung 2006, 54, 133–140. [Google Scholar]
- Guil-Guerrero, J.L.; Martínez-Guirado, C.; Rebolloso-Fuentes, M.M.; Carrique-Pérez, A. Nutrient composition and antioxidant activity of 10 pepper (Capsicum annuum) varieties. Eur. Food Res. Technol 2006, 224, 1–9. [Google Scholar]
- Rodríguez-Burruezo, A.; Prohens, J.; Raigón, M.D.; Nuez, F. Variations for bioactive compounds in ají (Capsicum baccatum L.) and rocoto (C. pubescens R. & P.) and implications for breeding. Euphytica 2009, 170, 169–181. [Google Scholar]
- Herbers, K. Vitamin production in transgenic plants. J. Plant Physiol 2003, 160, 821–829. [Google Scholar]
- Kim, M.-J.; Kim, J.K.; Kim, H.J.; Pak, J.H.; Lee, J.-H.; Kim, D.-H.; Choi, H.K.; Jung, H.W.; Lee, J.-D.; Chung, Y.-S.; et al. PLos One 2012, 7, e48287.


{kind=link}
{kind=link}
| Chili pepper type | Ripening stage | Main carotenoids | Total carotenoids | Reference |
|---|---|---|---|---|
| Red bell pepper | Mature (Red) | Capsanthin (34.7%), β-carotene (11.6%) and violaxanthin (9.9%) | 284 and 127 mg per kg f.wt. (as β-carotene) (two lots of peppers) | [27] |
| Green bell pepper | Immature (Green) | Lutein (40.8%), neoxanthin (15.1%), violaxanthin (13.8%) and β-carotene (13.4%) | 10.6, 11.2 and 9.0 mg per kg f.wt. (as β-carotene) (three lots of green peppers) | [52] |
| Yellow pepper | Immature (Green) | Violaxanthin (34%), antheraxanthin (10.5%), lutein (9.2%) and zeaxanthin (8.5%) | 13.2 mg/100 g d.wt. | [53] |
| Mature (Orange) | Lutein (37.8%), β-carotene (19.8%) and neoxanthin (5.5%) | 488.6 mg/100 g d.wt. | ||
| Black paprika | Immature (Black) | Lutein (28.5%) and zeaxanthin (11.96%) | 48.5 mg/100 g of d.wt. | [54] |
| Mature (Red) | Capsanthin (42%), zeaxanthin (8%), capsorubin (3.2%) and β-carotene (7%) | 3211 mg/100 g d.wt. | ||
| Szentesi Kosszarvú | Immature (Green) | Lutein (31.9%) and β-carotene (11.3%) | 11.5 mg/100 g d.wt. | [48] |
| Mature (Red) | Capsanthin (29%) and zeaxanthin (15%) | 994.7 mg/100 g d.wt. | ||
| Capsicum annuum var. lycopersiciforme rubrum | Immature (Green) | Lutein (31.6%) and β-carotene (13.7%) | 19.6 mg/100 g d.wt. | [55] |
| Mature (Red) | Capsanthin (37%), zeaxanthin (8%), and β-carotene (9%) | 1297.1 mg/100 g d.wt. | ||
| Sweet peppers | Immature (Green) | Lutein (2.3 mg/100 g f.wt.) and β-carotene (1.7 mg/100 g f.wt.) | 5.1 mg/100 g f.wt. | [32] |
| Green | Lutein (1.4 mg/100 g f.wt.) and β-carotene (2.1 mg/100 g f.wt.) | 4.9 mg/100 g f.wt. | ||
| Immature (Red) | β-carotene (1.9 mg/100 g f.wt.) and zeaxanthin (2.9 mg/100 g f.wt.) | 9.5 mg/100 g f.wt. | ||
| Red | β-carotene (4.3 mg/100 g f.wt.) and capsanthin (19.9 mg/100 g f.wt.) | 45.6 mg/100 g f.wt. | ||
| Ancho, guajillo and mulato | Ancho (Mature) | β-carotene (20.9%), and violaxanthin (14.5%) | 7.5 mg/100 g d.wt. | [56] |
| Guajillo (Mature) | β-carotene (17.9%), violaxanthin (13.2%) | 6.8 mg/100 g d.wt. | ||
| Mulato (Mature) | Violaxanthin (22%) β-carotene (14.9%) | 7.2 mg/100 g d.wt. |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gómez-García, M.D.R.; Ochoa-Alejo, N. Biochemistry and Molecular Biology of Carotenoid Biosynthesis in Chili Peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025-19053. https://doi.org/10.3390/ijms140919025
Gómez-García MDR, Ochoa-Alejo N. Biochemistry and Molecular Biology of Carotenoid Biosynthesis in Chili Peppers (Capsicum spp.). International Journal of Molecular Sciences. 2013; 14(9):19025-19053. https://doi.org/10.3390/ijms140919025
Chicago/Turabian StyleGómez-García, María Del Rocío, and Neftalí Ochoa-Alejo. 2013. "Biochemistry and Molecular Biology of Carotenoid Biosynthesis in Chili Peppers (Capsicum spp.)" International Journal of Molecular Sciences 14, no. 9: 19025-19053. https://doi.org/10.3390/ijms140919025
APA StyleGómez-García, M. D. R., & Ochoa-Alejo, N. (2013). Biochemistry and Molecular Biology of Carotenoid Biosynthesis in Chili Peppers (Capsicum spp.). International Journal of Molecular Sciences, 14(9), 19025-19053. https://doi.org/10.3390/ijms140919025