X-Ray Induced DNA Damage and Repair in Germ Cells of PARP1−/− Male Mice

Abstract
:1. Introduction
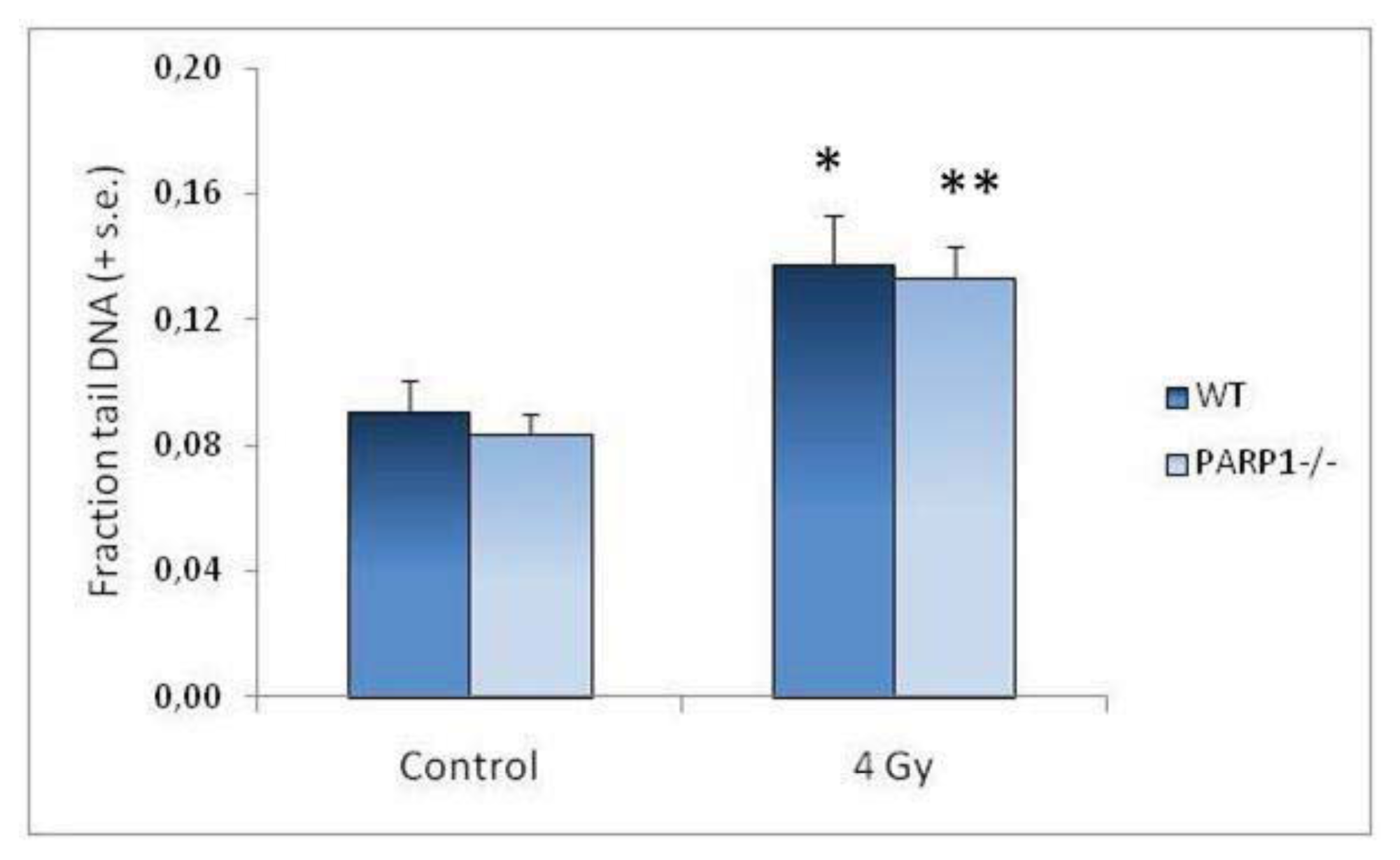
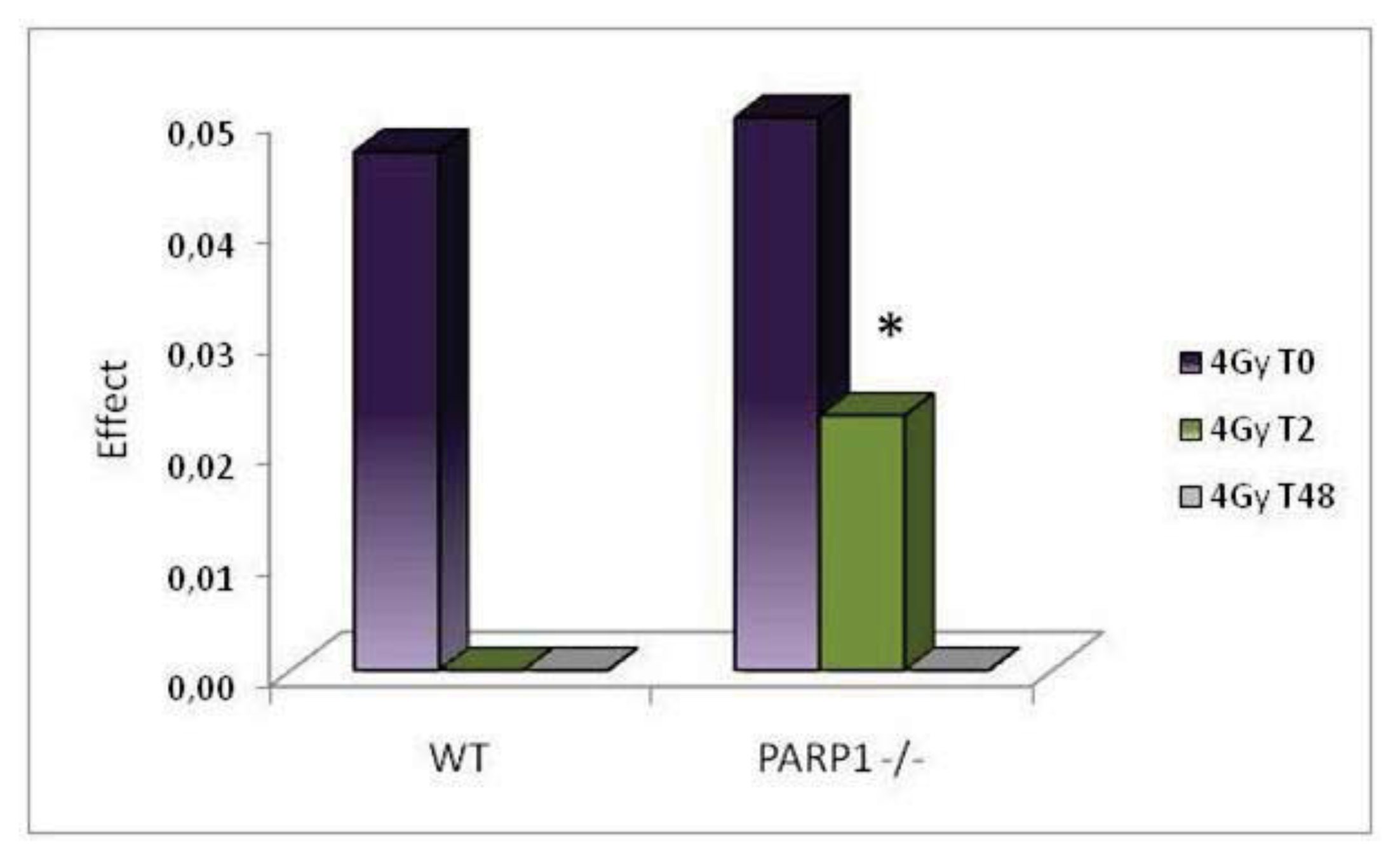
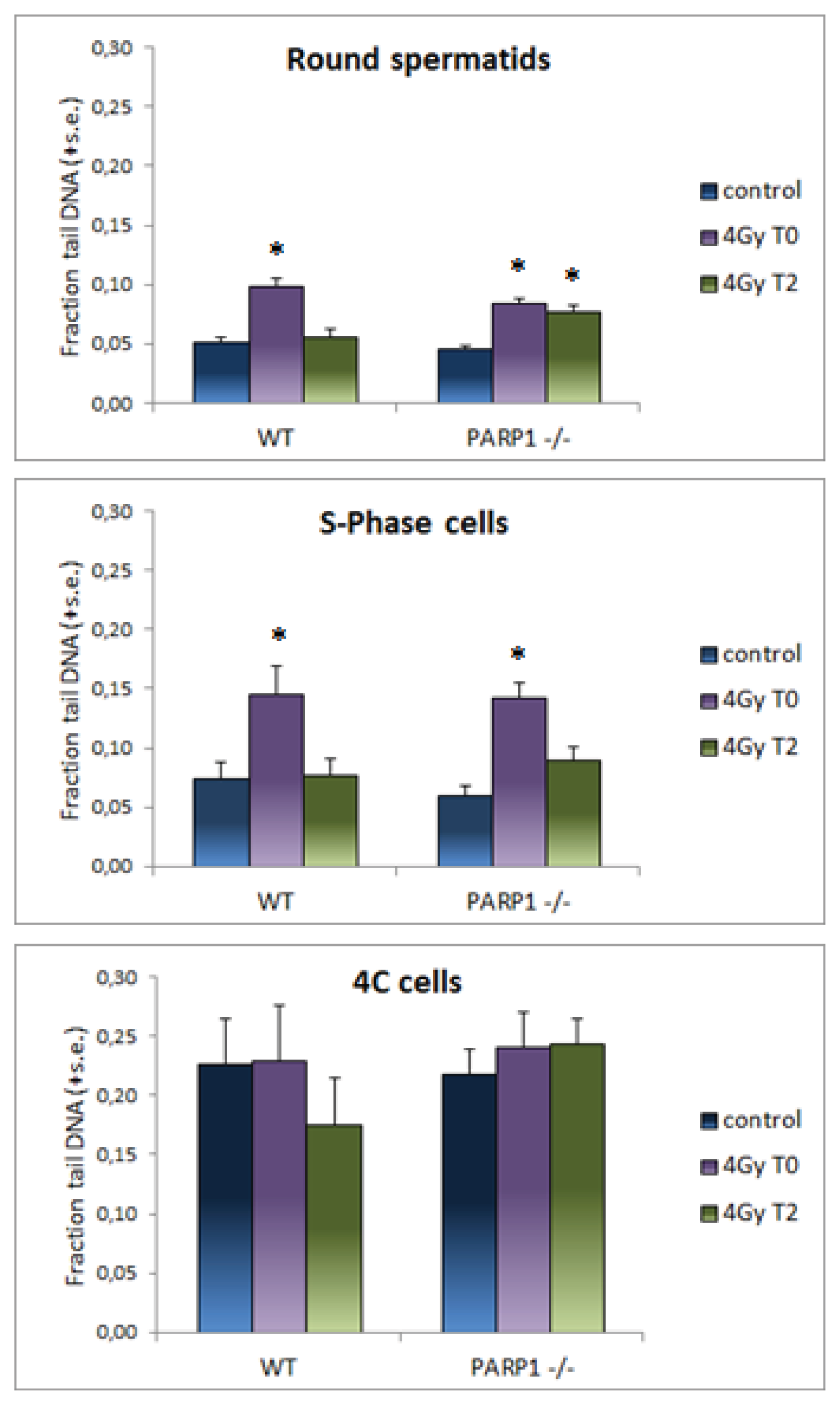
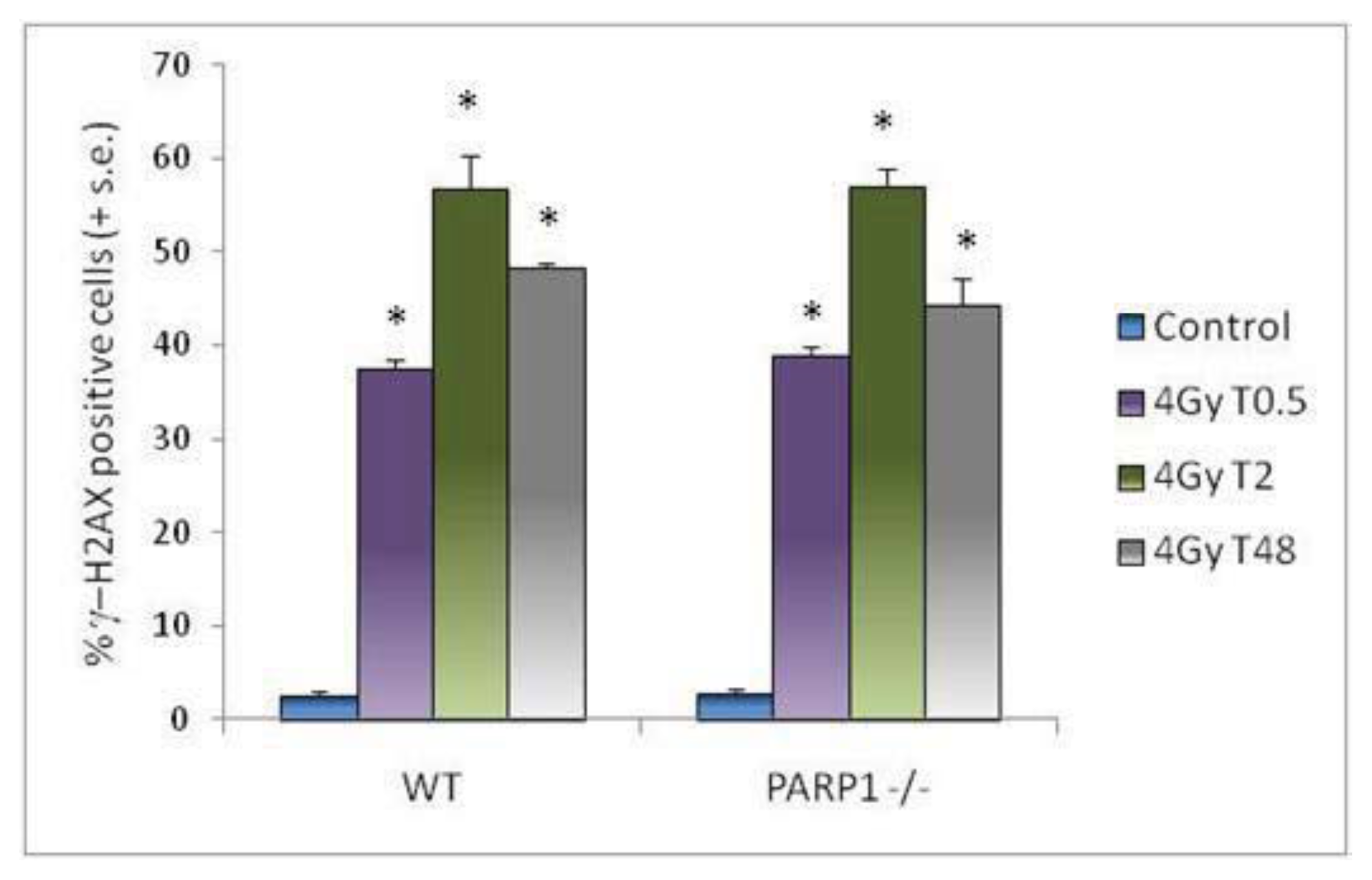
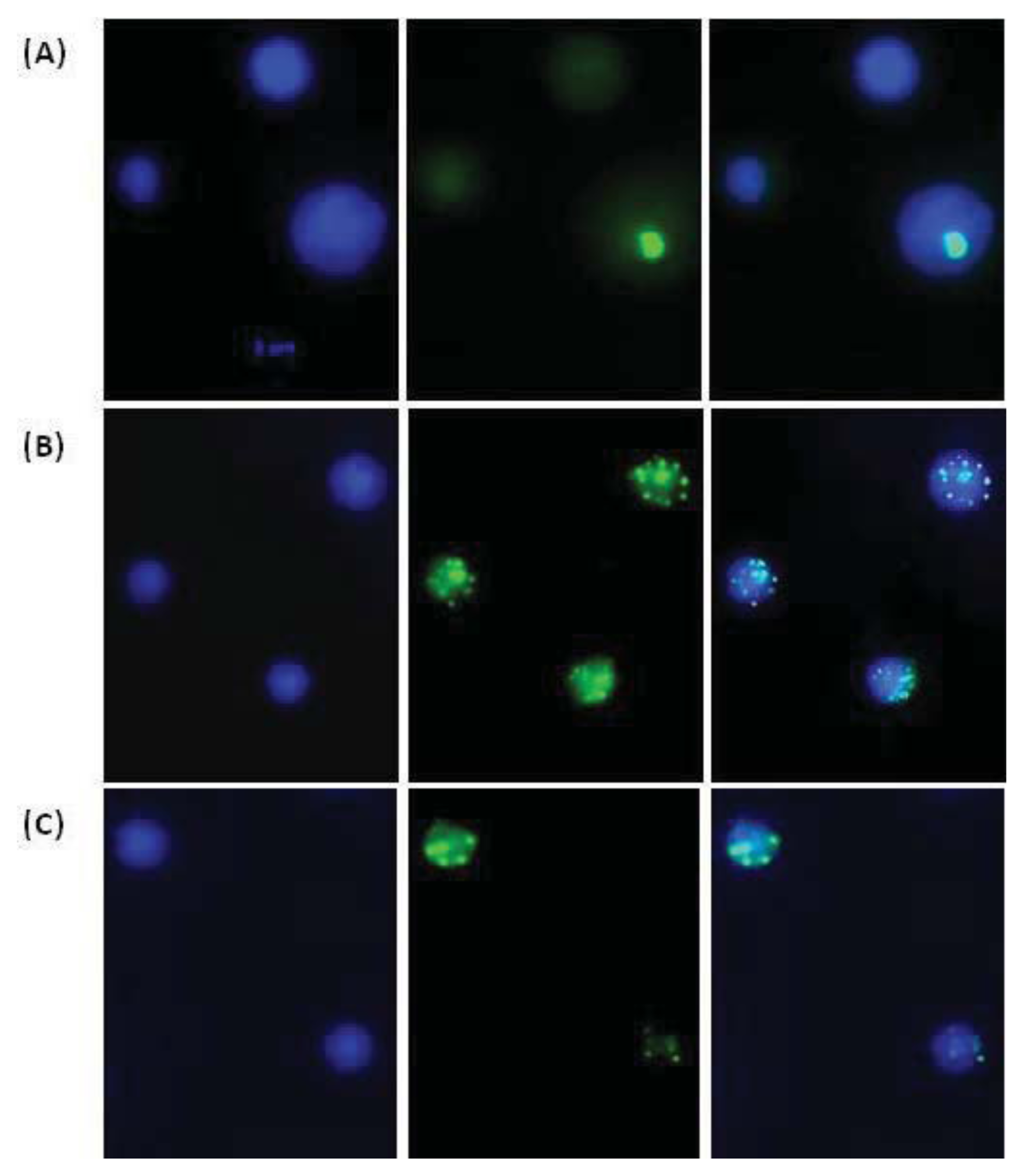
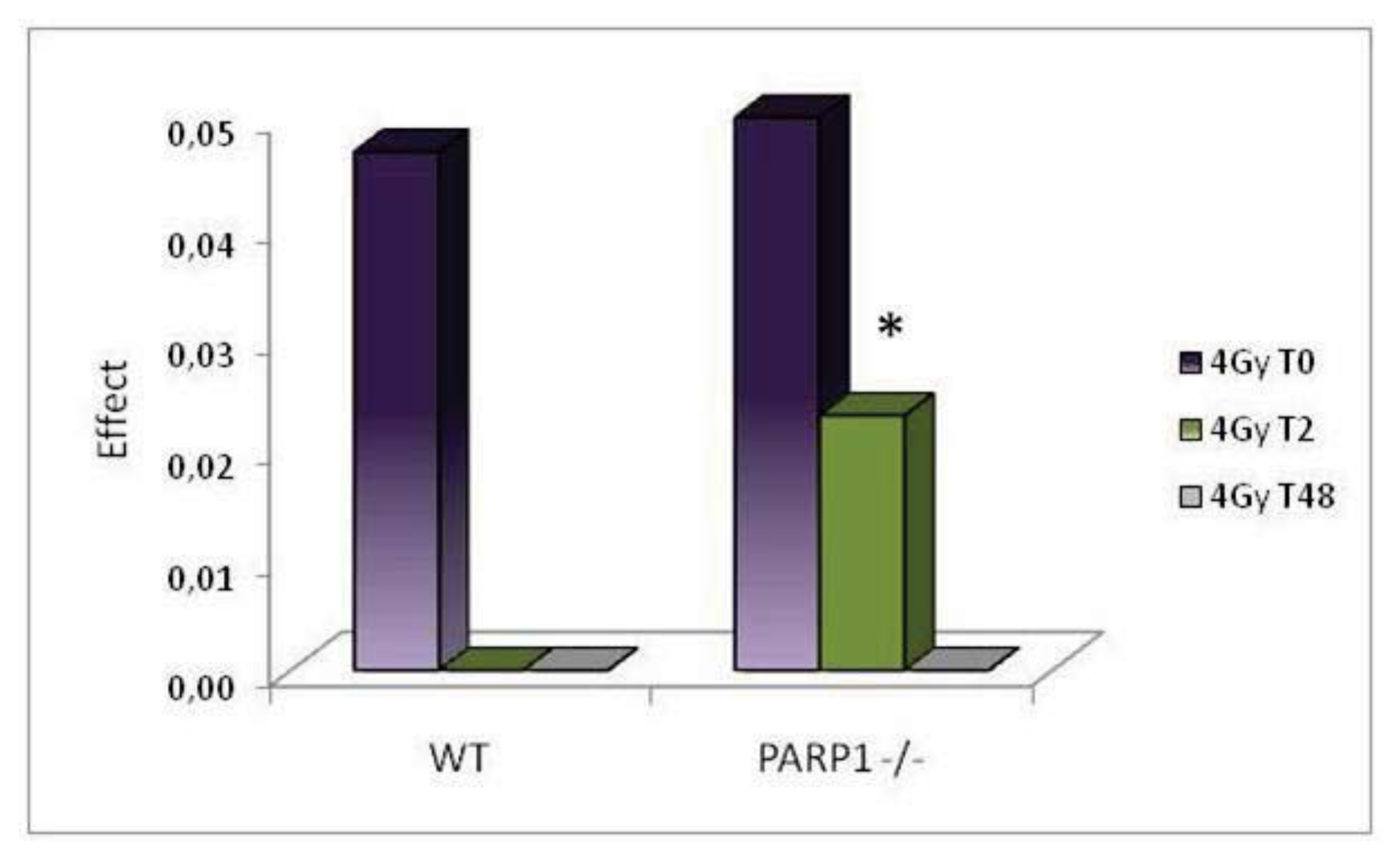
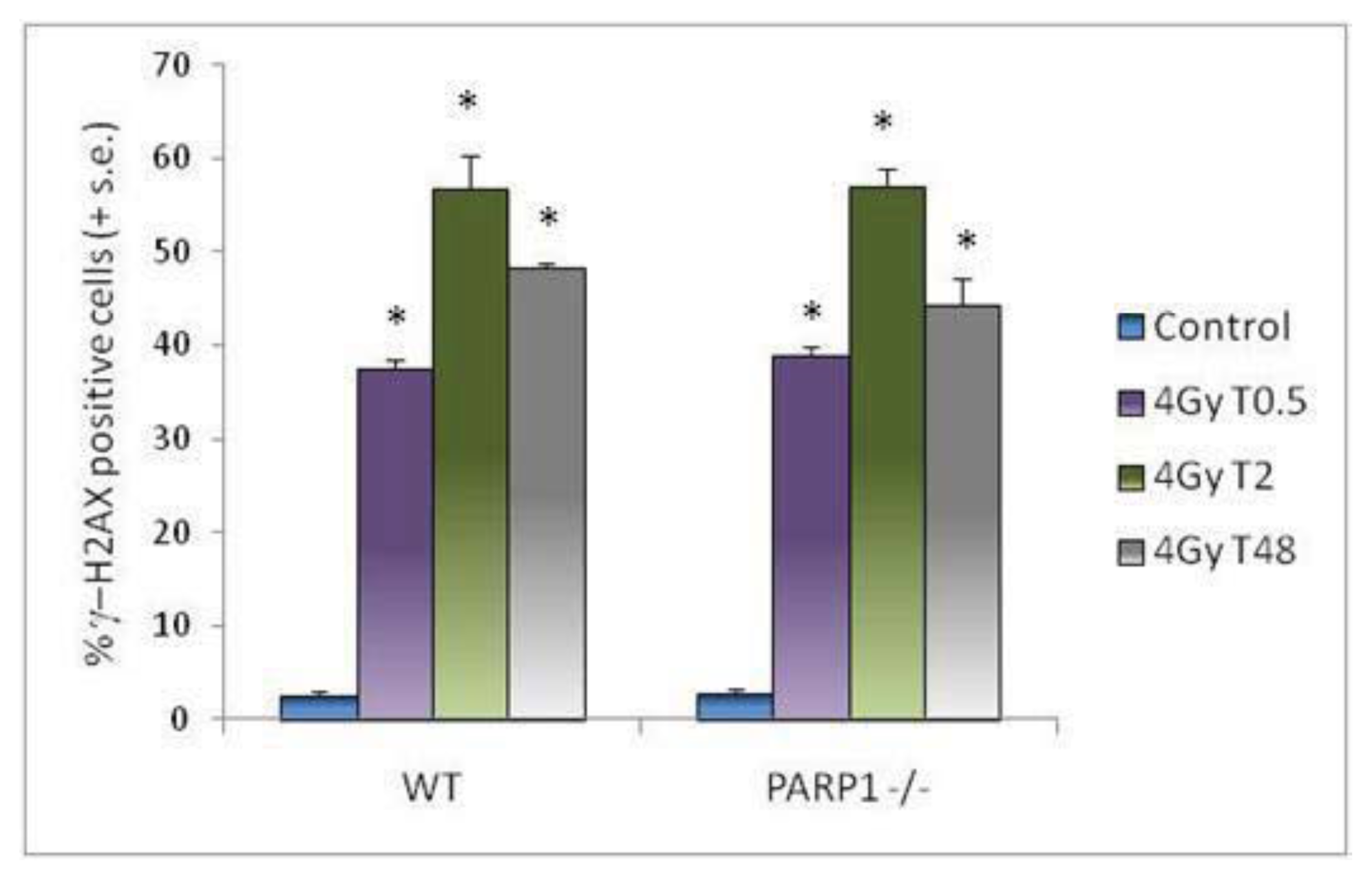
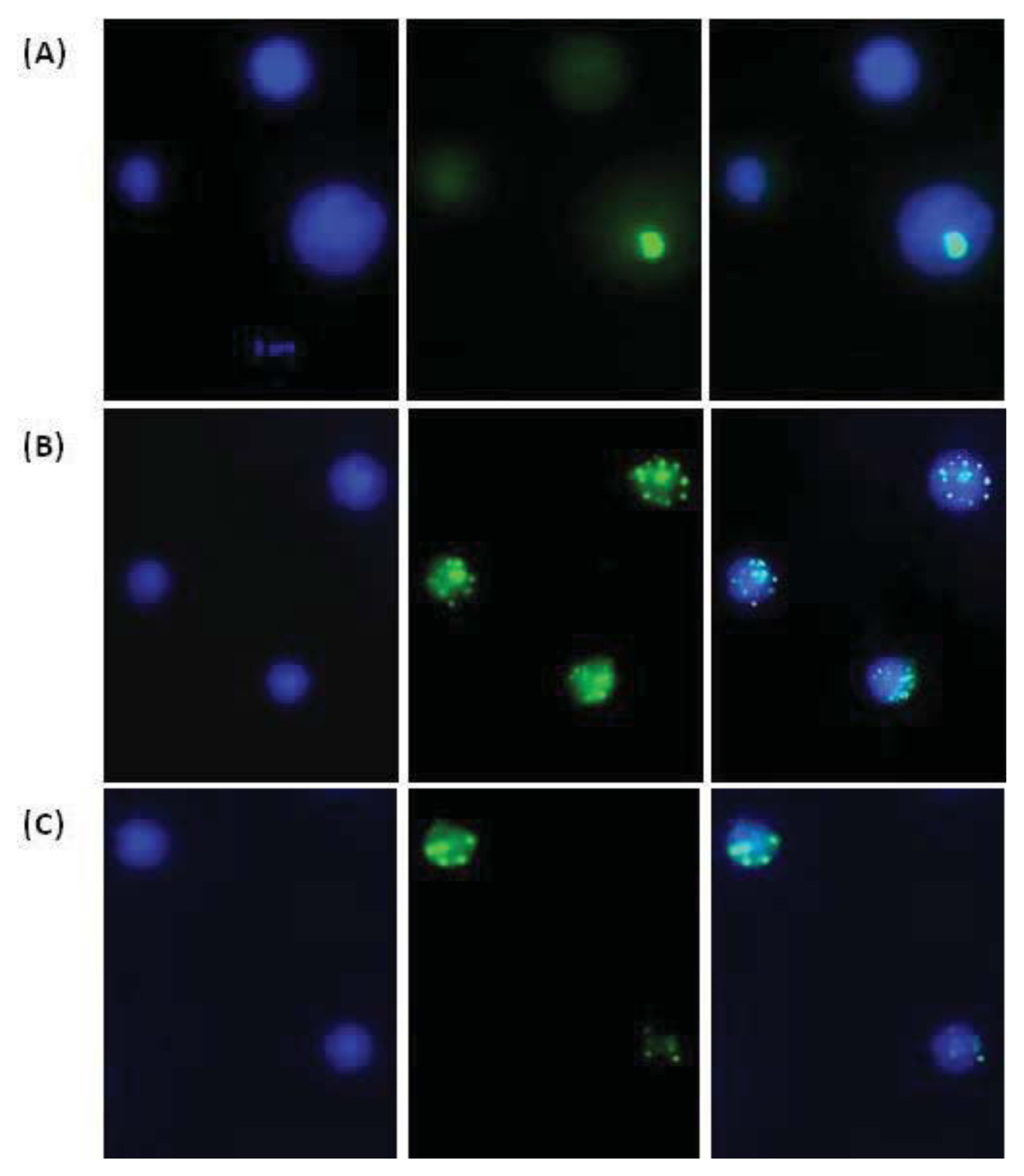
2. Results and Discussion
3. Experimental Section
3.1. Mice
3.2. X-Ray Irradiation
3.3. Comet Assay of Testis Cells
3.4. Immunofluorescent Analysis of H2AX Histone Phosphorylation
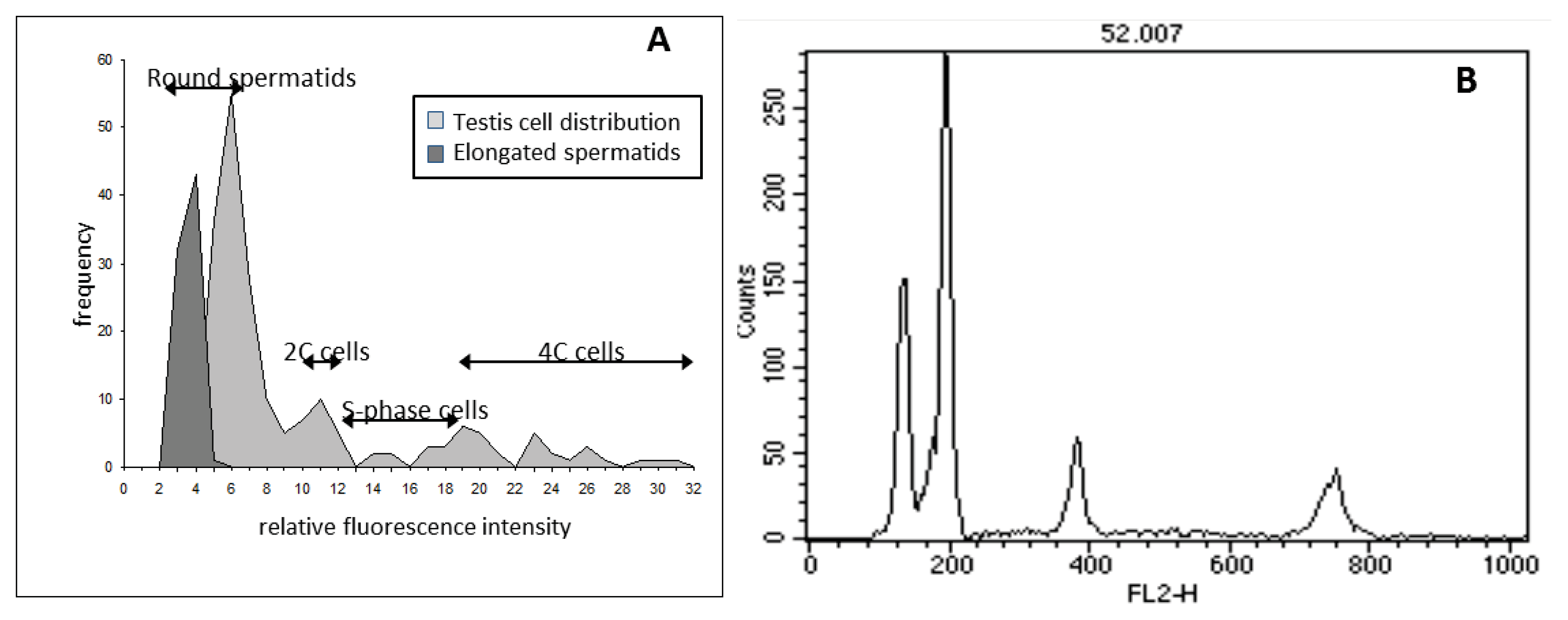
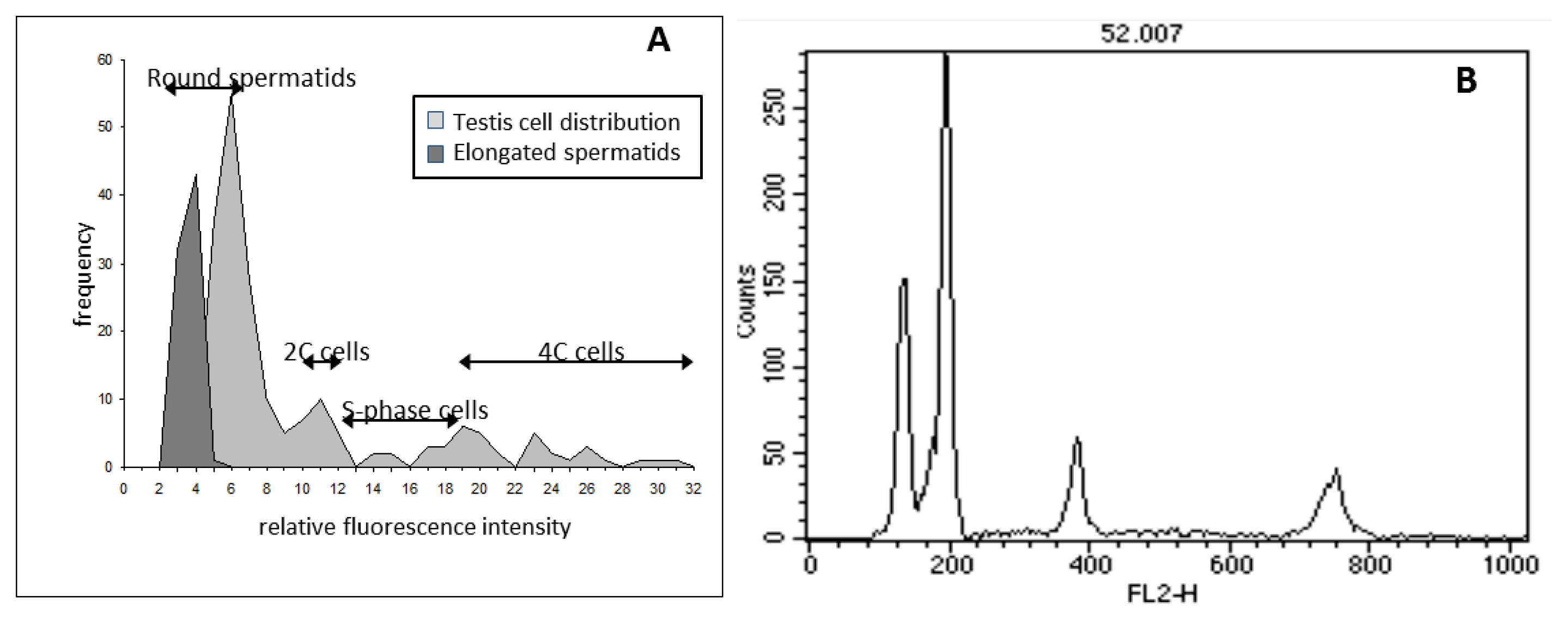
3.5. Flow Cytometric DNA Content Analysis of Testis Cells
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Brinkworth, M.H. Paternal trasmission of genetic damage: Findings in animals and humans. Int. J. Androl 2000, 23, 123–135. [Google Scholar]
- Ahmed, E.A.; van der Vaart, A.; Barten, A.; Kal, H.B.; Chen, J.; Lou, Z.; Minter-Dykhouse, K.; Bartkova, J.; Bartek, J.; de Boer, P.; et al. Differences in DNA double strand breaks repair in male germ cell types: Lessons learned from a differential expression of Mdc1 and 53BP1. DNA Repair (Amst. ) 2007, 6, 1243–1254. [Google Scholar]
- Ahmed, E.A.; de Boer, P.; Philippensc, E.P.; Kal, H.B.; de Rooij, D.G. Parp1–XRCC1 and the repair of DNA double strand breaks in mouse round spermatids. Mutat. Res 2010, 683, 84–90. [Google Scholar]
- Olsen, A.K.; Bjortuft, H.; Wiger, R.; Holme, J.; Seeberg, E.; Bjoras, M.; Brunborg, G. Highly efficient base excision repair (BER) in human and rat male germ cells. Nucleic Acid Res. 2001, 29, 1781–1790. [Google Scholar]
- Olsen, A.K.; Lindeman, B.; Wiger, R.; Duale, N.; Brunborg, G. How do male germ cells handle DNA damage? Toxicol. Appl. Pharmacol 2005, 207, 521–531. [Google Scholar]
- Sathees, C.R.; Raman, M.J. Mouse testicular extracts process DNA double-strand breaks efficiently by DNA end-to-end joining. Mutat. Res 1999, 433, 1–13. [Google Scholar]
- Srivastava, N.; Raman, M.J. Homologous recombination-mediated double-strand break repair in mouse testicular extracts and comparison with different germ cell stages. Cell Biochem. Funct 2007, 25, 75–86. [Google Scholar]
- Xu, G.; Spivak, G.; Mitchell, D.L.; Mori, T.; McCarrey, J.R.; McMahan, C.A.; Walter, R.B.; Hanawalt, P.C.; Walter, C.A. Nucleotide excision repair activity varies among murine spermatogenic cell types. Biol. Reprod 2005, 73, 123–130. [Google Scholar]
- Zheng, H.; Olive, P.L. Influence of oxigen on radiation induced DNA damage in testicular cells of C3H mice. Int. J. Radiat. Biol 1997, 71, 275–282. [Google Scholar]
- Kikland, J.B. Poly ADP-ribose polymerase-1 and health. Exp. Biol. Med 2010, 568, 235–561. [Google Scholar]
- Bouchard, V.J.; Rouleau, M.; Poirier, G.G. PARP-1, a determinant of cell survival in response to DNA damage. Exp. Hematol 2003, 31, 446–454. [Google Scholar]
- Kim, M.Y.; Zhang, T.W.; Kim, L.K. Poly(ADP-ribosyl)ation by PARP-1: ‘PAR-laying’ NAD+ into a nuclear signal. Genes Dev 2005, 19, 1951–1967. [Google Scholar]
- Dantzer, F.; Schreiber, V.; Niedergang, C.; Trucco, C.; Flatter, E.; de La Rubia, G.; Oliver, J.; Rolli, V.; Menisier-de Murcia, J.; de Murcia, G. Involvement of poly(ADP-ribose)polymerase in base excision repair. Biochimie 1999, 81, 69–75. [Google Scholar]
- Schreiber, V.; Ame, J.; Dollè, P.; Schultz, I.; Rinaldi, B.; Fraulob, V.; deMurcia, J.M.; deMurcia, G. Poly(ADP-ribose) polimerase-2 (PARP-2) is required for efficient base excision DNA repair in association with PARP-1 and XRCC1. J. Biol. Chem 2002, 277, 23028–23036. [Google Scholar]
- Krishnakumar, R.; Kraus, W.L. The PARP side of the nucleus: Molecular actions, physiological outcomes, and clinical targets. Mol. Cell 2010, 39, 8–24. [Google Scholar]
- Cardoso, R.S.; Espanhol, A.R.; Passos, G.A.S.; Sakamoto-Hojo, E.T. Differential gene expression in γ-irradiated BALB/3T3 fibroblast under the influence of 3-aminobenzamide, an inhibitor of PARP enzyme. Mutat. Res 2002, 508, 33–40. [Google Scholar]
- D’Amours, D.; Desnoyers, S.; D’Silva, I.; Poirier, G.G. Poly(ADP-ribosyl)ation reactions in the regulation of nuclear functions. Biochem. J 1999, 342, 249–268. [Google Scholar]
- Masutani, M.; Nozaki, T.; Nakamoto, K.; Nakagama, H.; Suzuki, H.; Kusuoka, O.; Tsutsumi, M.; Sugimura, T. The response of PARP knockout mice against DNA damaging agents. Mutat. Res 2000, 462, 159–166. [Google Scholar]
- Masutani, M.; Nakagama, H.; Sugimura, T. Poly(ADP-ribose) and carcinogenesis. Genes Chromosom. Cancer 2003, 38, 339–348. [Google Scholar]
- Meyer-Ficca, M.L.; Meyer, R.G.; Jacobson, E.L.; Jacobson, M.K. Poly(ADP-ribose)polymerases: Managing genome stability. Int. J. Biochem. Cell Biol 2005, 37, 920–926. [Google Scholar]
- Muiras, M.L. Mammalian longevity under the protection of PARP-1’s multi-facets. Ageing Res. Rev 2003, 2, 129–148. [Google Scholar]
- Petermann, E.; Keil, C.; Oei, S.L. Importance of poly(ADP-ribose)polymerases in the regulation of DNA-dependent processes. Cell. Mol. Life Sci 2005, 62, 731–738. [Google Scholar]
- Satoh, M.S.; Lindahl, T. Role of poly(ADP-ribose) formation in DNA repair. Nature 1992, 356, 356–358. [Google Scholar]
- Shall, S.; de Murcia, G. Poly(ADP-ribose)polymerase-1: What have we learned from the deficient mouse model? Mutat. Res 2000, 460, 1–15. [Google Scholar]
- Atorino, L.; di Meglio, S.; Farina, B.; Jones, R.; Quesada, P. Rat germinal cells require PARP for repair of DNA damage induced by γ irradiation and H2O2 treatment. Eur. J. Cell Biol 2001, 80, 222–229. [Google Scholar]
- Di Meglio, S.; Denegri, M.; Vallefuoco, S.; Tramontano, F.; Scovassi, A.I.; Quesada, P. Poly(ADPR)polymerase-1 and poly(ADPR) glycohydrolase level and distribution in differentiating rat germinal cells. Mol. Cell. Biochem 2003, 248, 85–91. [Google Scholar]
- Di Meglio, S.; Tramontano, F.; Cimmino, G.; Jones, R.; Quesada, P. Dual role of poly(ADP-ribose) polymerase-1 and −2 and poly(ADP-ribose) glycohydrolase as DNA-repair and pro-apoptotic factors in rat germinal cells exposed to nitric oxide donors. Biochim. Biophys. Acta 2004, 1692, 35–44. [Google Scholar]
- Meyer-Ficca, M.L.; Scherthan, H.; Burkle, A.; Meyer, R.G. Poly(ADP-ribosyl)ation during chromatin remodeling steps in rat spermiogenesis. Chromosoma 2005, 114, 67–74. [Google Scholar]
- Quesada, P.; Atorino, L.; Cardone, A.; Ciarcia, G.; Farina, B. Poly(ADPribosyl)ation system in rat germinal cells at different stages of differentiation. Exp. Cell Res 1996, 226, 183–190. [Google Scholar]
- Tramontano, F.; Malanga, M.; Quesada, P. Differential contribution of poly(ADP-ribose)polymerase-1 and −2 (PARP-1 and −2) to the poly(ADP-ribosyl)ation reaction in rat primary spermatocytes. Mol. Hum. Reprod 2007, 13, 821–828. [Google Scholar]
- Meyer-Ficca, M.L.; Lonchar, J.; Credidio, C.; Ihara, M.; Li, Y.; Wang, Z.Q.; Meyer, R.G. Disruption of poly(ADP-ribose) homeostasis affects spermiogenesis and sperm chromatin integrity in mice. Biol. Reprod 2009, 81, 46–55. [Google Scholar]
- Meyer-Ficca, M.L.; Ihara, M.; Lonchar, J.D.; Meistrich, M.L.; Austin, C.A.; Min, W.; Wang, Z.Q.; Meyer, R.G. Poly(ADP-ribose) Metabolism is essential for proper nucleoprotein exchange during mouse spermiogenesis. Biol. Reprod 2011, 84, 218–228. [Google Scholar]
- Masutani, M.; Nozaki, T.; Nishiyama, E.; Shimokawa, T.; Tachi, Y.; Suzuki, H.; Nakagama, H.; Wakabayashi, K.; Sugimura, T. Function of poly(ADP-ribose)polymerase in response to DNA damage: Gene-distruption study in mice. Mol. Cell. Biochem 1999, 193, 149–152. [Google Scholar]
- Menisier-deMurcia, J.; Niedergang, C.; Trucco, C.; Recoul, M.; Dutrillaux, B.; Mark, M.; Olivier, F.J.; Massan, M.; Dierich, A.; LeMeur, M.; et al. Requirement of poly(ADP-ribose)polymerase in recovery from DNA damage in mice and in cells. Proc. Natl. Acad. Sci. USA 1997, 94, 7303–7307. [Google Scholar]
- Wang, Z.Q.; Auer, B.; Stingl, L.; Berghammer, H.; Haidacher, D.; Schweiger, M.; Wagner, E.F. Mice lacking ADPRT and poly(ADP-ribosyl)ation develop normally but are susceptible to skin disease. Genes Dev 1995, 9, 509–520. [Google Scholar]
- Cordelli, E.; Fresegna, A.M.; Leter, G.; Eleuteri, P.; Spanò, M.; Villani, P. Evaluation of DNA damage in different stages of mouse spermatogenesis after testicular X-irradiation. Radiat. Res 2003, 160, 443–451. [Google Scholar]
- Paris, L.; Cordelli, E.; Eleuteri, P.; Grollino, M.G.; Pasquali, E.; Ranaldi, R.; Meschini, R.; Pacchierotti, F. Kinetics of γ-H2AX induction and removal in bone marrow and testicular cells of mice after X-ray irradiation. Mutagenesis 2011, 26, 563–572. [Google Scholar]
- Hacker-Klom, U.; Goehde, W.; Schumann, J. Mammalian Spermatogenesis as a Biological Dosimeter for Ionizing Radiation. In Biological Dosimetry; Eisert, W.G., Mendelsohn, M.L., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 1984; pp. 127–137. [Google Scholar]
- Cieslar-Pobudaa, A.; Saenkoa, Y.; Rzeszowska-Wolnya, J. PARP-1 inhibition induces a late increase in the level of reactive oxygen species in cells after ionizing radiation. Mutat. Res 2012, 732, 9–15. [Google Scholar]
- Villani, P.; Altavista, P.L.; Castaldi, L.; Leter, G.; Cordelli, E. Analysis of DNA oxidative damage related to cell proliferation. Mutat. Res 2000, 464, 229–237. [Google Scholar]
- Mahadevaiah, S.K.; Turner, J.M.; Baudat, F.; Rogakou, E.P.; de Boer, P.; Blanco-Rodriguez, V.; Jasin, M.; Keeney, S.; Bonner, W.M.; Burgoyne, P.S. Recombinational DNA double-strand breaks in mice precede synapsis. Nat. Genet 2001, 27, 271–276. [Google Scholar]
- Hamer, G.; Roepers-Gajadien, H.L.; van Duyn-Goedhart, A.; Gademan, I.S.; Kal, H.B.; van Buul, P.P.W.; de Rooij, D.G. DNA double-strand breaks and γ-H2AX signaling in the testis. Biol. Reprod 2003, 68, 628–634. [Google Scholar]
- Hamer, G.; Roepers-Gajadien, H.L.; Gademan, I.S.; Kal, H.B.; de Rooij, D.G. Intracellular bridges and apoptosis in clones of male germ cells. Int. J. Androl 2003, 26, 348–353. [Google Scholar]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem 1998, 273, 5858–5868. [Google Scholar]
- Baure, J.; Izadi, A.; Suarez, V.; Giedzinski, E.; Cleaver, J.E.; Fike, J.R.; Limoli, C.L. Histone H2AX phosphorylation in response to changes in chromatin structure induced by altered osmolarity. Mutagenesis 2009, 24, 161–167. [Google Scholar]
- Soutoglou, E.; Misteli, T. Activation of the cellular DNA damage response in the absence of DNA lesions. Science 2008, 320, 1507–1510. [Google Scholar]
- Ichijima, Y.; Sakasai, R.; Okita, N.; Asahina, K.; Mizutani, S.; Teraoka, H. Phosphorylation of histone H2AX at M phase in human cells without DNA damage response. Biochem. Biophys. Res. Commun 2005, 336, 807–812. [Google Scholar]
- Fernandez-Capetillo, O.; Mahadevaiah, S.K.; Celeste, A.; Romanienko, P.J.; Camerini-Otero, R.D.; Bonner, W.M.; Manova, K.; Burgoyne, P.; Nussenzweig, A. H2AX is required for chromatin remodelling and inactivation of sex chromosomes in male mouse meiosis. Dev. Cell 2003, 4, 497–508. [Google Scholar]
- Olive, P.L.; Banath, J.P. Phosphorylation of histone H2AX as a measure of radiosensitivity. Int. J. Radiat. Oncol. Biol. Phys 2004, 58, 331–335. [Google Scholar]
- Forand, A.; Dutrillaux, B.; Bernardino-Sgherri, J. Gamma-H2AX expression pattern in non-irradiated neonatal mouse germ cells and after low-dose gamma-radiation: Relationships between chromatid breaks and DNA double-strand breaks. Biol. Reprod 2004, 71, 643–649. [Google Scholar]
- Haince, J.F.; Kozlov, S.; Dawson, V.L.; Dawson, T.M.; Hendzel, M.J.; Lavin, M.F.; Poirier, G.G. Ataxia Telangiectasia Mutated (ATM) signaling network is modulated by a novel poly(ADP-ribose)-dependent pathway in the early response to DNA-damaging agents. J. Biol. Chem 2007, 282, 16441–16453. [Google Scholar]
- Bouquet, F.; Muller, C.; Salles, B. The loss of gammaH2AX signal is a marker of DNA double strand breaks repair only at low levels of DNA damage. Cell Cycle 2006, 5, 1116–1122. [Google Scholar]
- Costes, S.V.; Chiolo, I.; Pluth, J.M.; Barcellos-Hoff, M.H.; Jakob, B. Spatiotemporal characterization of ionizing radiation induced DNA damage foci and their relation to chromatin organization. Mutat. Res 2010, 704, 78–87. [Google Scholar]
- Markova, E.; Schultz, N.; Belyaev, I.Y. Kinetics and dose response of residual 53BP1/gamma-H2AX foci: Co-localization, relationship with DSB repair and clonogenic survival. Int. J. Radiat. Biol 2007, 83, 319–329. [Google Scholar]
- Godon, C.; Cordelieres, F.P.; Biard, D.; Giocanti, N.; Megnin-Chanet, F.; Hall, J.; Favaudon, V. PARP inhibition versus PARP-1 silencing: Different outcomes in terms of single-strand break repair and radiation susceptibility. Nucleic Acids Res 2008, 36, 4454–4464. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res 1988, 175, 184–191. [Google Scholar]
- Spanò, M.; Bartoleschi, C.; Cordelli, E.; Leter, G.; Segre, L.; Mantovani, A.; Fazzi, P.; Pacchierotti, F. Flow cytometric and histological assessment of 1,2,3,4-diepoxybutane toxicity on mouse spermatogenesis. J. Toxicol. Environ. Health 1996, 47, 101–119. [Google Scholar]
- Zante, J.; Schumann, J.; Gohde, W.; Hacker, U. DNA fluorocytometry of mammalian sperm. Histochemistry 1977, 54, 1–7. [Google Scholar]
- Lampariello, F.; Mauro, F.; Uccelli, R.; Spanò, M. Automatic analysis of flow cytometric DNA histograms from irradiated mouse male germ cells. Cytometry 1989, 10, 62–69. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elongated spermatids | Round spermatids | Diploid cells | S-phase cells | Tetraploid cells | |
|---|---|---|---|---|---|
| WT control | 23.11 (1.06) | 49.85 (1.13) | 11.32 (0.52) | 2.69 (0.13) | 10.90 (0.20) |
| PARP1−/− control | 23.71 (0.31) | 48.42 (0.66) | 11.36 (0.31) | 2.46 (0.16) | 11.07 (0.27) |
| WT 4 Gy 48 h | 23.21 (0.61) | 49.32 (0.93) | 12.13 (0.91) | 1.04 (0.22) b | 10.94 (0.33) |
| PARP1−/− 4 Gy 48 h | 26.28 (0.79) a | 46.83 (0.93) | 11.57 (0.54) | 0.70 (0.11) b | 11.29 (0.27) |
| WT Fraction tail DNA (± s.e.) | PARP1−/− Fraction tail DNA (± s.e.) | |
|---|---|---|
| Control | 0.09 (0.01) | 0.08 (0.01) |
| 4 Gy T0 | 0.14 (0.02) a | 0.13 (0.01) a |
| 4 Gy T2 | 0.08 (0.01) | 0.11 (0.01) a |
| 4 Gy T48 | 0.09 (0.01) | 0.08 (0.01) |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Villani, P.; Fresegna, A.M.; Ranaldi, R.; Eleuteri, P.; Paris, L.; Pacchierotti, F.; Cordelli, E. X-Ray Induced DNA Damage and Repair in Germ Cells of PARP1−/− Male Mice. Int. J. Mol. Sci. 2013, 14, 18078-18092. https://doi.org/10.3390/ijms140918078
Villani P, Fresegna AM, Ranaldi R, Eleuteri P, Paris L, Pacchierotti F, Cordelli E. X-Ray Induced DNA Damage and Repair in Germ Cells of PARP1−/− Male Mice. International Journal of Molecular Sciences. 2013; 14(9):18078-18092. https://doi.org/10.3390/ijms140918078
Chicago/Turabian StyleVillani, Paola, Anna Maria Fresegna, Roberto Ranaldi, Patrizia Eleuteri, Lorena Paris, Francesca Pacchierotti, and Eugenia Cordelli. 2013. "X-Ray Induced DNA Damage and Repair in Germ Cells of PARP1−/− Male Mice" International Journal of Molecular Sciences 14, no. 9: 18078-18092. https://doi.org/10.3390/ijms140918078
APA StyleVillani, P., Fresegna, A. M., Ranaldi, R., Eleuteri, P., Paris, L., Pacchierotti, F., & Cordelli, E. (2013). X-Ray Induced DNA Damage and Repair in Germ Cells of PARP1−/− Male Mice. International Journal of Molecular Sciences, 14(9), 18078-18092. https://doi.org/10.3390/ijms140918078