Proteogenomic Characterization of Novel x-Type High Molecular Weight Glutenin Subunit 1Ax1.1



Abstract
:1. Introduction
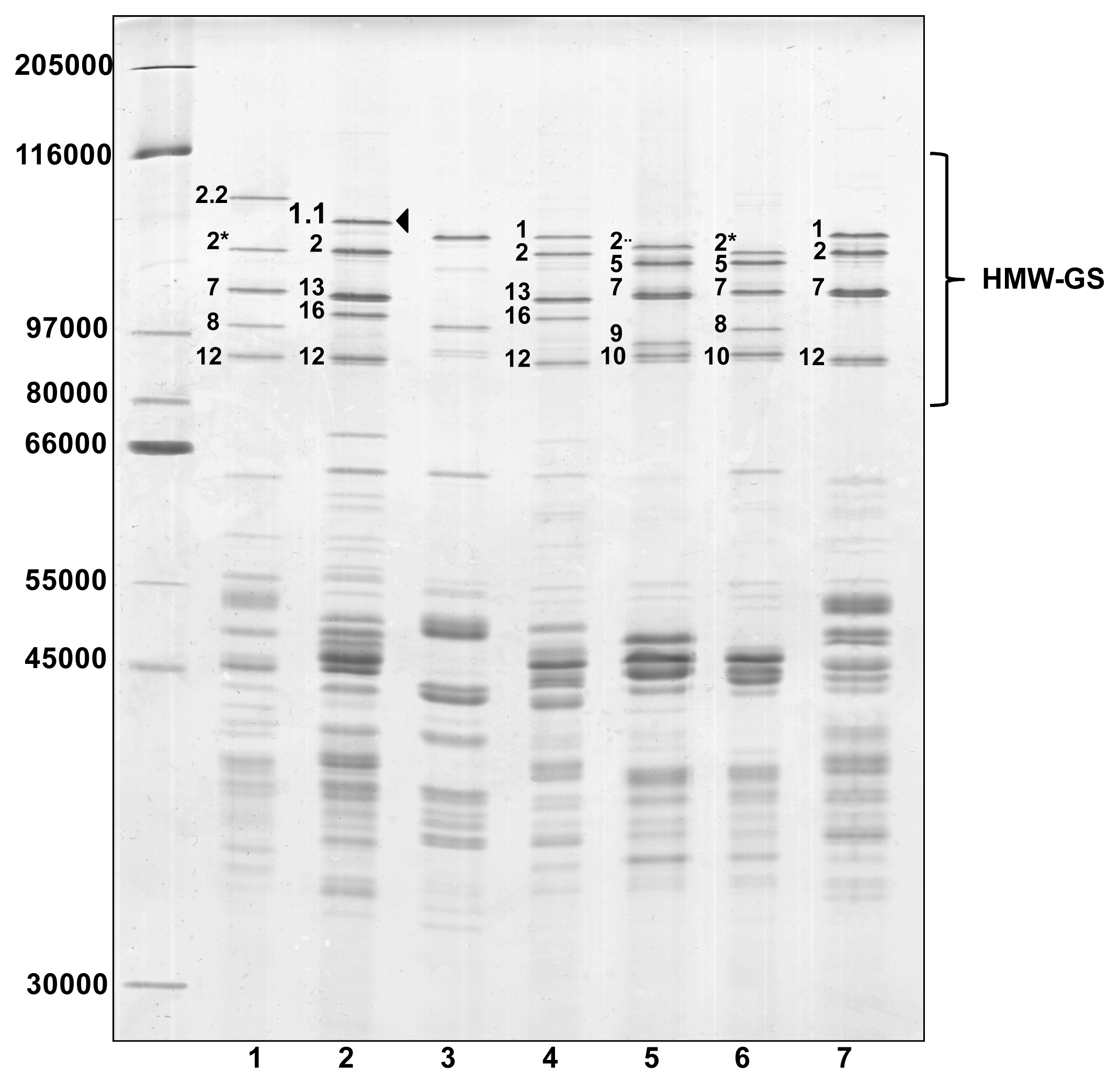
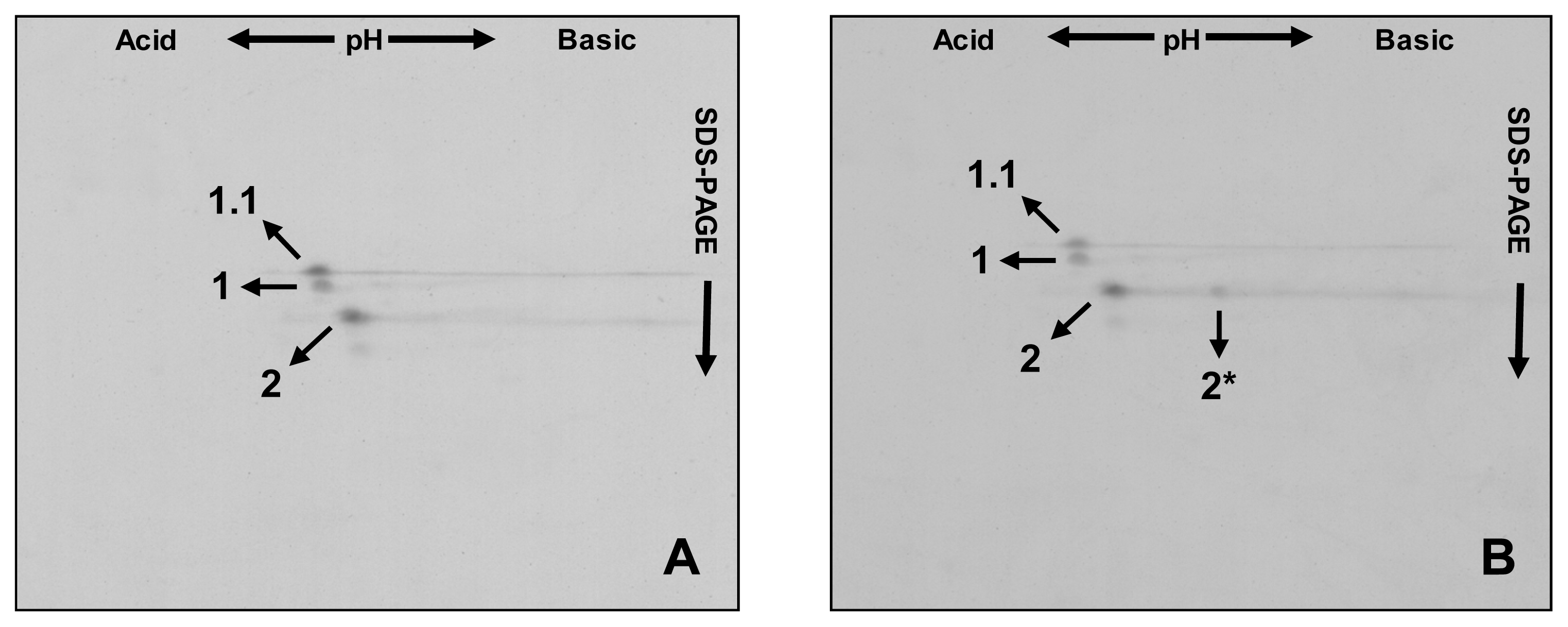
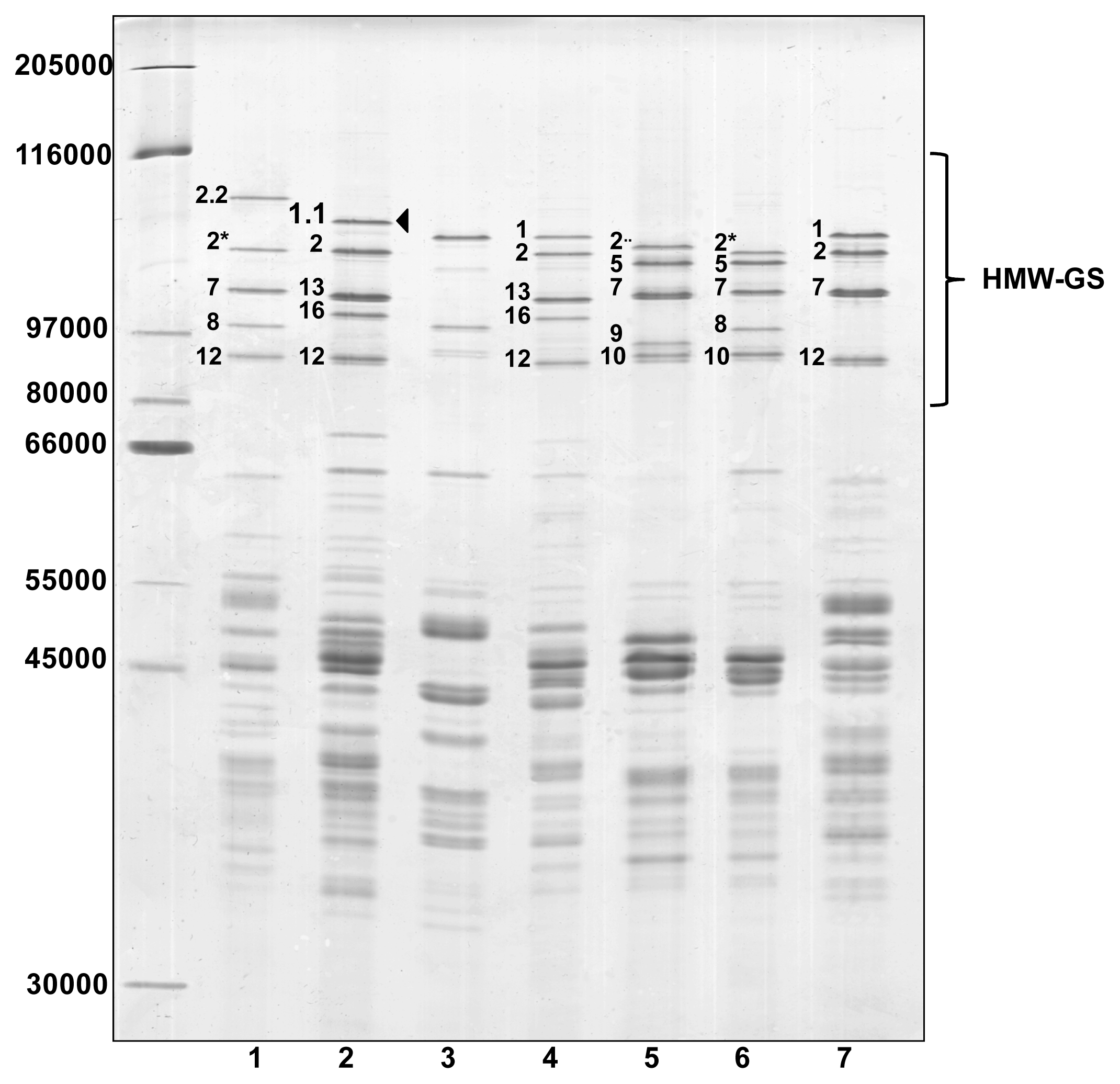
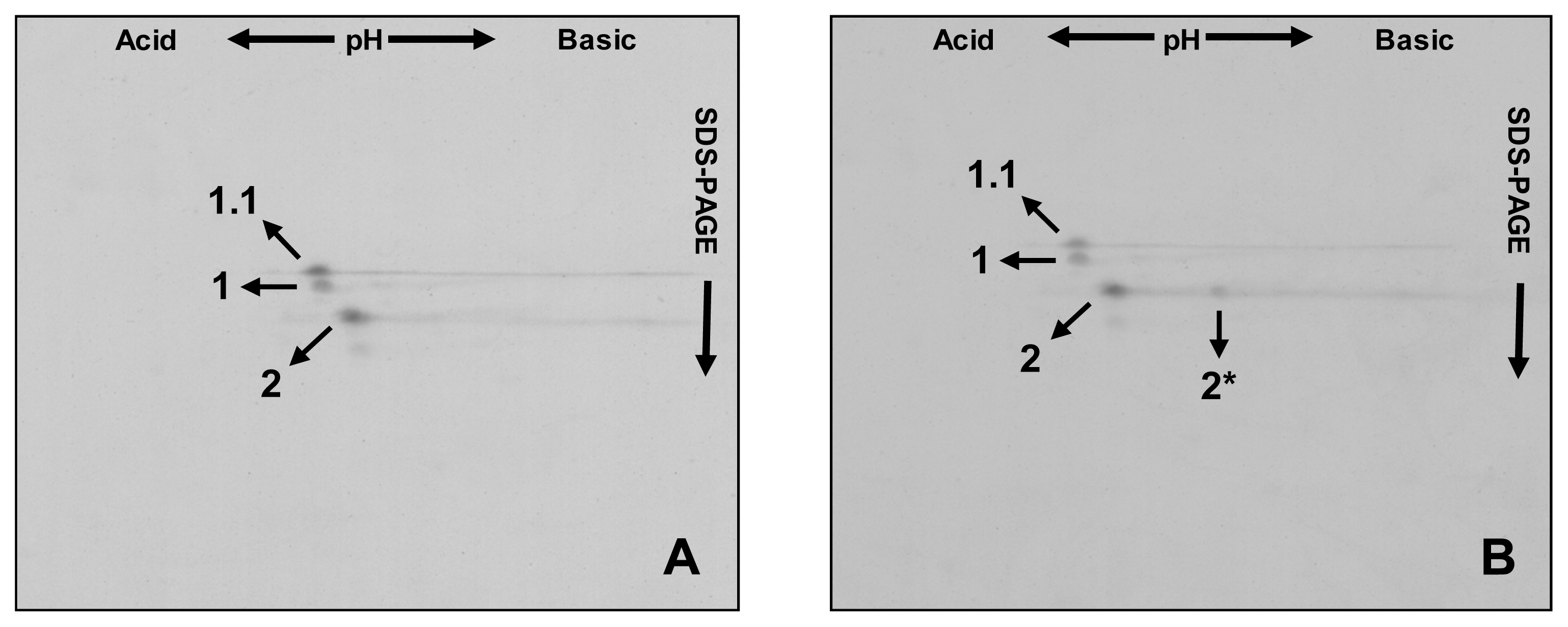
2. Results and Discussion
3. Experimental Section
3.1. Plant Materials
3.2. Technological Tests
3.3. Electrophoresis
3.4. Protein Identification by Mass Spectrometry (MS)
3.5. Analysis of the HMW-GS 1Ax1.1 Coding Sequence
4. Conclusions
Acknowledgments
References
- Osborne, T.B. The Proteins of the Wheat Kernel; Carnegie Institution of Washington: Washington, DC, USA; p. 1907.
- Lawrence, G.J.; Shepherd, K.W. Variation in glutenin protein subunits of wheat. Aust. J. Biol. Sci 1980, 33, 221–233. [Google Scholar]
- Payne, P.I.; Holt, L.M.; Jackson, E.A.; Law, C.N.; Damania, A.B. Wheat storage proteins: Their genetics and their potential for manipulation by plant breeding. Philos. Trans. R. Soc. Lond. B Biol. Sci 1984, 304, 359–371. [Google Scholar]
- Macritchie, F. Physicochemical Properties of Wheat Proteins in Relation to Functionality. In Advances in Food and Nutrition Research; John, E.K., Ed.; Academic Press: New York, NY, USA, 1992; Volume 36, pp. 1–87. [Google Scholar]
- Payne, P.; Holt, L.; Law, C. Structural and genetical studies on the high-molecular-weight subunits of wheat glutenin. Theor. Appl. Genet 1981, 60, 229–236. [Google Scholar]
- Tatham, A.S.; Miflin, B.J.; Shewry, P.R. The beta-turn conformation in wheat gluten proteins— Relationship to gluten elasticity. Cereal Chem 1985, 62, 405–412. [Google Scholar]
- Tatham, A.S.; Drake, A.F.; Shewry, P.R. Conformational studies of synthetic peptides corresponding to the repetitive regions of the High-molecular-weight (Hmw) glutenin subunits of wheat. J. Cereal Sci 1990, 11, 189–200. [Google Scholar]
- Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet 2003, 49, 111–184. [Google Scholar]
- Shewry, P.R.; Tatham, A.S. Disulphide bonds in wheat gluten proteins. J. Cereal Sci 1997, 25, 207–227. [Google Scholar]
- Buonocore, F.; Bertini, L.; Ronchi, C.; Bekes, F.; Caporale, C.; Lafiandra, D.; Gras, P.; Tatham, A.S.; Greenfield, J.A.; Halford, N.G. Expression and functional analysis of Mr 58,000 peptides derived from the repetitive domain of high molecular weight glutenin subunit 1Dx5. J. Cereal Sci 1998, 27, 209–215. [Google Scholar]
- Anderson, O.D.; Greene, F.C.; Yip, R.E.; Halford, N.G.; Shewry, P.R.; Malpica-Romero, J.-M. Nucleotide sequences of the two high-molecular-weight glutenin genes from the D-genome of a hexaploid bread wheat, Triticum aestivum L. cv Cheyenne. Nucleic Acids Res 1989, 17, 461–462. [Google Scholar]
- Juhász, A.; Tamás, L.; Karsai, I.; Vida, G.; Láng, L.; Bedő, Z. Identification, cloning and characterisation of a HMW-glutenin gene from an old Hungarian wheat variety, Bánkúti 1201. Euphytica 2001, 119, 75–79. [Google Scholar]
- Veraverbeke, W.S.; Delcour, J.A. Wheat Protein Composition and Properties of Wheat Glutenin in Relation to Breadmaking Functionality. Crit. Rev. Food Sci. Nutr 2002, 42, 179–208. [Google Scholar]
- Anjum, F.M.; Khan, M.R.; Din, A.; Saeed, M.; Pasha, I.; Arshad, M.U. Wheat gluten: High molecular weight glutenin subunits—Structure, genetics, and relation to dough elasticity. J. Food Sci 2007, 72, R56–R63. [Google Scholar]
- Tilley, K.A.; Benjamin, R.E.; Bagorogoza, K.E.; Okot-Kotber, B.M.; Prakash, O.; Kwen, H. Tyrosine cross-links: Molecular basis of gluten structure and function. J. Agric. Food Chem 2001, 49, 2627–2632. [Google Scholar]
- Shewry, P.R.; Halford, N.G.; Tatham, A.S. High molecular weight subunits of wheat glutenin. J. Cereal Sci 1992, 15, 105–120. [Google Scholar]
- Gianibelli, M.C.; Larroque, O.R.; MacRitchie, F.; Wrigley, C.W. Biochemical, genetic, and molecular characterization of wheat glutenin and its component subunits. Cereal Chem 2001, 78, 635–646. [Google Scholar]
- Payne, P.I.; Lawrence, G.J. Catalogue of alleles for the complex gene loci Glu-A1, Glu-B1 and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun 1983, 11, 29–35. [Google Scholar]
- Branlard, G.; Autran, J.-C.; Rousset, M.; Dardevet, M.E.; Koenig, J. Catalogue des sous unités de haut poids moléculaire des gluténines des blés (T. aestivum e T. durum); Rapport AIP ‘Qualité des Blés’, INRA: Paris, France, 1990; p. 60. [Google Scholar]
- Shewry, P.R.; Popineau, Y.; Lafiandra, D.; Belton, P. Wheat glutenin subunits and dough elasticity: Findings of the EUROWHEAT project. Trends Food Sci. Technol 2000, 11, 433–441. [Google Scholar]
- Igrejas, G.; Branlard, G.; Carnide, V.; Gateau, I.; Guedes-Pinto, H. Storage protein diversity within the old Portuguese bread wheat Barbela population. J. Genet. Breed 1997, 51, 167–173. [Google Scholar]
- Igrejas, G.; Guedes-Pinto, H.; Carnide, V.; Branlard, G. The high and low molecular weight glutenin subunits and ω-gliadin composition of bread and durum wheats commonly grown in Portugal. Plant Breed 1999, 118, 297–302. [Google Scholar]
- Ribeiro, M.; Carvalho, C.; Carnide, V.; Guedes-Pinto, H.; Igrejas, G. Towards allelic diversity in the storage proteins of old and currently growing tetraploid and hexaploid wheats in Portugal. Genet. Resour. Crop Evol 2011, 58, 1051–1073. [Google Scholar]
- Igrejas, G.; Guedes-Pinto, H.; Carnide, V.; Clement, J.; Branlard, G. Genetical, biochemical and technological parameters associated with biscuit quality. II. Prediction using storage proteins and quality characteristics in a soft wheat population. J. Cereal Sci 2002, 36, 187–197. [Google Scholar]
- Li, Q.Y.; Yan, Y.M.; Wang, A.L.; An, X.L.; Zhang, Y.Z.; Hsam, S.L.K.; Zeller, F.J. Detection of HMW glutenin subunit variations among 205 cultivated emmer accessions (Triticum turgidum ssp. dicoccum). Plant Breed 2006, 125, 120–124. [Google Scholar]
- Gobaa, S.; Kleijer, G.; Stamp, P. 2··, a new high molecular weight glutenin subunit coded by Glu-A1: Its predicted structure and its impact on bread-making quality. Plant Breed 2007, 126, 1–4. [Google Scholar]
- Payne, P.I.; Nightingale, M.A.; Krattiger, A.F.; Holt, L.M. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric 1987, 40, 51–65. [Google Scholar]
- Halford, N.G.; Field, J.M.; Blair, H.; Urwin, P.; Moore, K.; Robert, L.; Thompson, R.; Flavell, R.B.; Tatham, A.S.; Shewry, P.R. Analysis of HMW glutenin subunits encoded by chromosome 1A of bread wheat (Triticum aestivum L.) indicates quantitative effects on grain quality. Theor. Appl. Genet 1992, 83, 373–378. [Google Scholar]
- Qian, Y.; Preston, K.; Krokhin, O.; Mellish, J.; Ens, W. Characterization of wheat gluten proteins by HPLC and MALDI TOF mass spectrometry. J. Am. Soc. Mass Spectrom 2008, 19, 1542–1550. [Google Scholar]
- Hickman, D.R.; Roepstorff, P.; Shewry, P.R.; Tatham, A.S. Molecular weights of high molecular weight subunits of glutenin determined by mass spectrometry. J. Cereal Sci 1995, 22, 99–103. [Google Scholar]
- Mamone, G.; de Caro, S.; di Luccia, A.; Addeo, F.; Ferranti, P. Proteomic-based analytical approach for the characterization of glutenin subunits in durum wheat. J. Mass Spectrom 2009, 44, 1709–1723. [Google Scholar]
- Don, C.; Mann, G.; Bekes, F.; Hamer, R.J. Linking Glutenin Particle size to HMW Glutenin Subunit Composition. In The Gluten Proteins; The Royal Society of Chemistry: London, UK, 2004; pp. 285–287. [Google Scholar]
- Pritchard, P.E. The glutenin fraction (gel-protein) of wheat protein—A new tool in the prediction of baking quality. Aspects Appl. Biol 1993, 36, 75–84. [Google Scholar]
- Weegels, P.L.; vandePijpekamp, A.M.; Graveland, A.; Hamer, R.J.; Schofield, J.D. Depolymerisation and re-polymerisation of wheat glutenin during dough processing. 1. Relationships between glutenin macropolymer content and quality parameters. J. Cereal Sci 1996, 23, 103–111. [Google Scholar]
- Kelfkens, M.; Lichtendonk, W.J. Verbesserungen in der Beurteilung der Backfahigkeit von Weizensorten. Getreide Mehl und Brot 2000, 54, 363–365. [Google Scholar]
- D’Ovidio, R.; Anderson, O.D.; Masci, S.; Skerritt, J.; Porceddu, E. Construction of novel wheat High-Mr glutenin subunit gene variability: Modification of the repetitive domain and expression in E. coli. J. Cereal Sci 1997, 25, 1–8. [Google Scholar]
- D’Ovidio, R.; Lafiandra, D.; Porceddu, E. Identification and molecular characterization of a large insertion within the repetitive domain of a high-molecular-weight glutenin subunit gene from hexaploid wheat. TAG Theor. Appl. Genet 1996, 93, 1048–1053. [Google Scholar]
- He, G.Y.; Jones, H.D.; D’Ovidio, R.; Masci, S.; Chen, M.; West, J.; Butow, B.; Anderson, O.D.; Lazzeri, P.; Fido, R.; Shewry, P.R. Expression of an extended HMW subunit in transgenic wheat and the effect on dough mixing properties. J. Cereal Sci 2005, 42, 225–231. [Google Scholar]
- Singh, N.K.; Shepherd, K.W.; Cornish, G.B. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J. Cereal Sci 1991, 14, 203–208. [Google Scholar]
- Dumur, J.; Jahier, J.; Bancel, E.; Lauriere, M.; Bernard, M.; Branlard, G. Proteomic analysis of aneuploid lines in the homeologous group 1 of the hexaploid wheat cultivar Courtot. Proteomics 2004, 4, 2685–2695. [Google Scholar]
- Liu, L.; Wang, A.; Appels, R.; Ma, J.; Xia, X.; Lan, P.; He, Z.; Bekes, F.; Yan, Y.; Ma, W. A MALDI-TOF based analysis of high molecular weight glutenin subunits for wheat breeding. J. Cereal Sci 2009, 50, 295–301. [Google Scholar]
- Stein, N.; Herren, G.; Keller, B. A new DNA extraction method for high-throughput marker analysis in a large-genome species such as Triticum aestivum. Plant Breed 2001, 120, 354–356. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
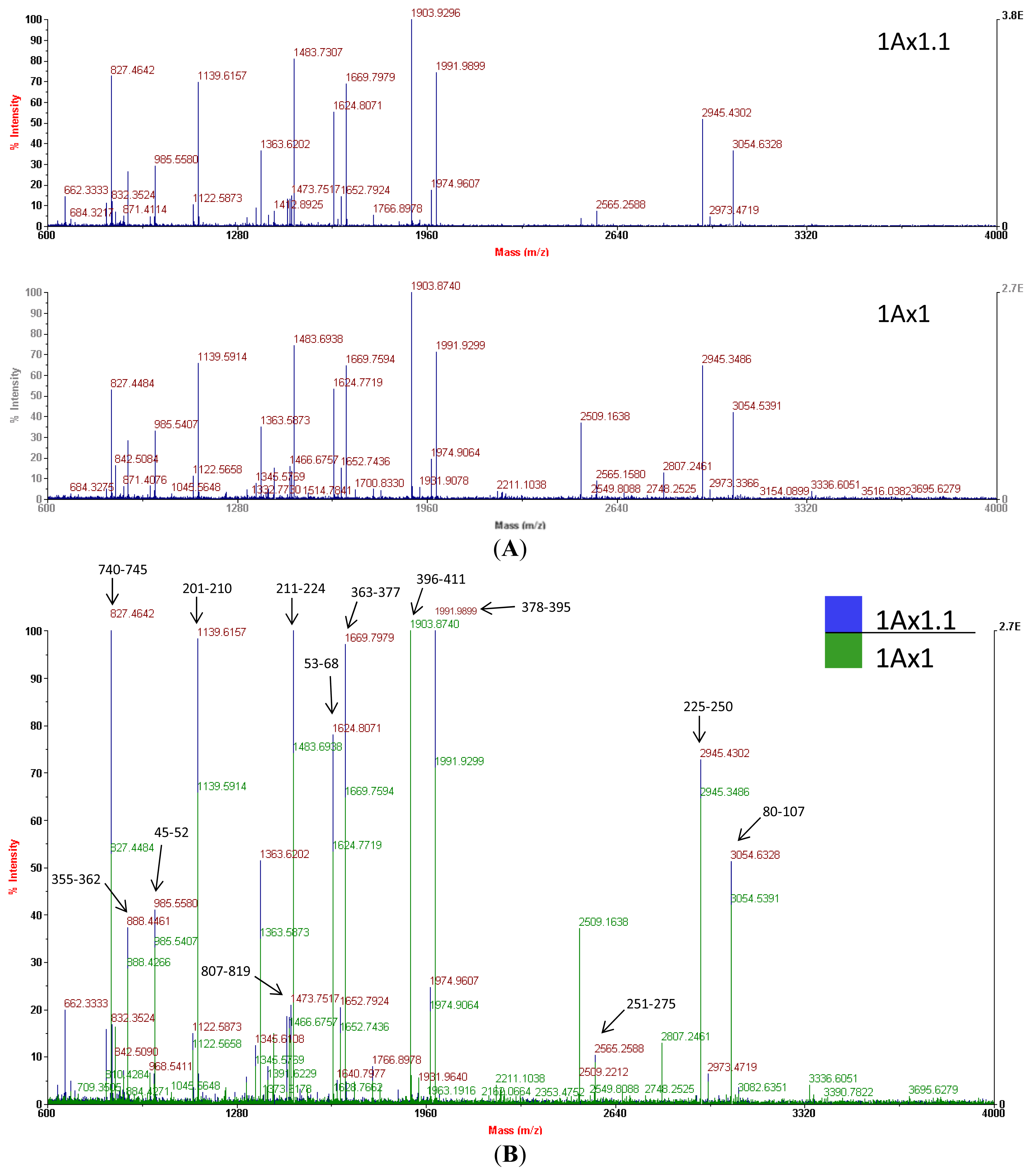{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HMW-GS | Deduced Mr from coding gene | Mr by MALDI-TOF-MS | Variation | |||
|---|---|---|---|---|---|---|
| Origin | Mass (Da) | Origin | Mass (Da) | Difference (Da) | Error (%) | |
| 1Ax1 | ‘Hope’ | 87,678 | ‘Carala’ | 87,859 | −181 | 0.21 |
| 1Ax1.1 | ‘Barbela 28’ | 91,508 | ‘Barbela 28’ | 91,858 | −350 | 0.38 |
| Comparison of Means | Alveograph Parameters | Protein Content % dm | GrainHardness | |||||
|---|---|---|---|---|---|---|---|---|
| Location | Genotype (Glu-A1; Glu-B1) | P mmH20 | L mm | W 10−4 J | P/L mmH20 mm−1 | Ie | ||
| Portugal | 1; 7 + 8 | 44.8 | 101.3 | 80.3 | 0.5 | 25.1 | 15.0 | 53.7 |
| 1.1; 7 + 8 | 51.7 | 110.7 | 114.4 | 0.5 | 27.6 | 14.2 | 35.6 | |
| 1; 13 + 16 | 63.8 | 143.4 | 184.0 | 0.4 | 35.2 | 14.3 | 23.0 | |
| 1.1; 13 + 16 | 57.8 | 119.6 | 141.1 | 0.5 | 30.9 | 15.3 | 41.9 | |
| France | 1; 7 + 8 | 70.4 | 208.6 | 295.1 | 0.33 | 44.7 | 17.5 | 77.0 |
| 1.1; 7 + 8 | 72.6 | 181.8 | 243.1 | 0.40 | 37.0 | 14.8 | 49.7 | |
| 1; 13 + 16 | 93.5 | 148.8 | 370.5 | 0.63 | 50.2 | 14.8 | 49.1 | |
| 1.1; 13 + 16 | 70.4 | 131.8 | 224.9 | 0.53 | 42.7 | 15.1 | 60.7 | |
| Variety | Glu-A1 | Glu-B1 | Glu-D1 | |||
|---|---|---|---|---|---|---|
| Allele | HMW-GS | Allele | HMW-GS | Allele | HMW-GS | |
| ‘Fukihokomugi’ | b | 2 * | b | 7 + 8 | f | 2.2 + 12 |
| ‘Barbela 28’ | * | 1.1 | f | 13 + 16 | a | 2 + 12 |
| ‘PI 355’ | * | * | * | * | * | * |
| ‘Carala’ | a | 1 | f | 13 + 16 | a | 2 + 12 |
| ‘211.12014’ | f | 2·· | c | 7 + 9 | d | 5 + 10 |
| ‘13-21’ | b | 2* | b | 7 + 8 | d | 5 + 10 |
| ‘Konini’ | a | 1 | a | 7 | a | 2 + 12 |
| ‘Atlas 66’ | b | 2* | f | 13 + 16 | a | 2 + 12 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ribeiro, M.; Bancel, E.; Faye, A.; Dardevet, M.; Ravel, C.; Branlard, G.; Igrejas, G. Proteogenomic Characterization of Novel x-Type High Molecular Weight Glutenin Subunit 1Ax1.1. Int. J. Mol. Sci. 2013, 14, 5650-5667. https://doi.org/10.3390/ijms14035650
Ribeiro M, Bancel E, Faye A, Dardevet M, Ravel C, Branlard G, Igrejas G. Proteogenomic Characterization of Novel x-Type High Molecular Weight Glutenin Subunit 1Ax1.1. International Journal of Molecular Sciences. 2013; 14(3):5650-5667. https://doi.org/10.3390/ijms14035650
Chicago/Turabian StyleRibeiro, Miguel, Emmanuelle Bancel, Annie Faye, Mireille Dardevet, Catherine Ravel, Gérard Branlard, and Gilberto Igrejas. 2013. "Proteogenomic Characterization of Novel x-Type High Molecular Weight Glutenin Subunit 1Ax1.1" International Journal of Molecular Sciences 14, no. 3: 5650-5667. https://doi.org/10.3390/ijms14035650
APA StyleRibeiro, M., Bancel, E., Faye, A., Dardevet, M., Ravel, C., Branlard, G., & Igrejas, G. (2013). Proteogenomic Characterization of Novel x-Type High Molecular Weight Glutenin Subunit 1Ax1.1. International Journal of Molecular Sciences, 14(3), 5650-5667. https://doi.org/10.3390/ijms14035650