Metabolic Difference of CZ48 in Human and Mouse Liver Microsomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
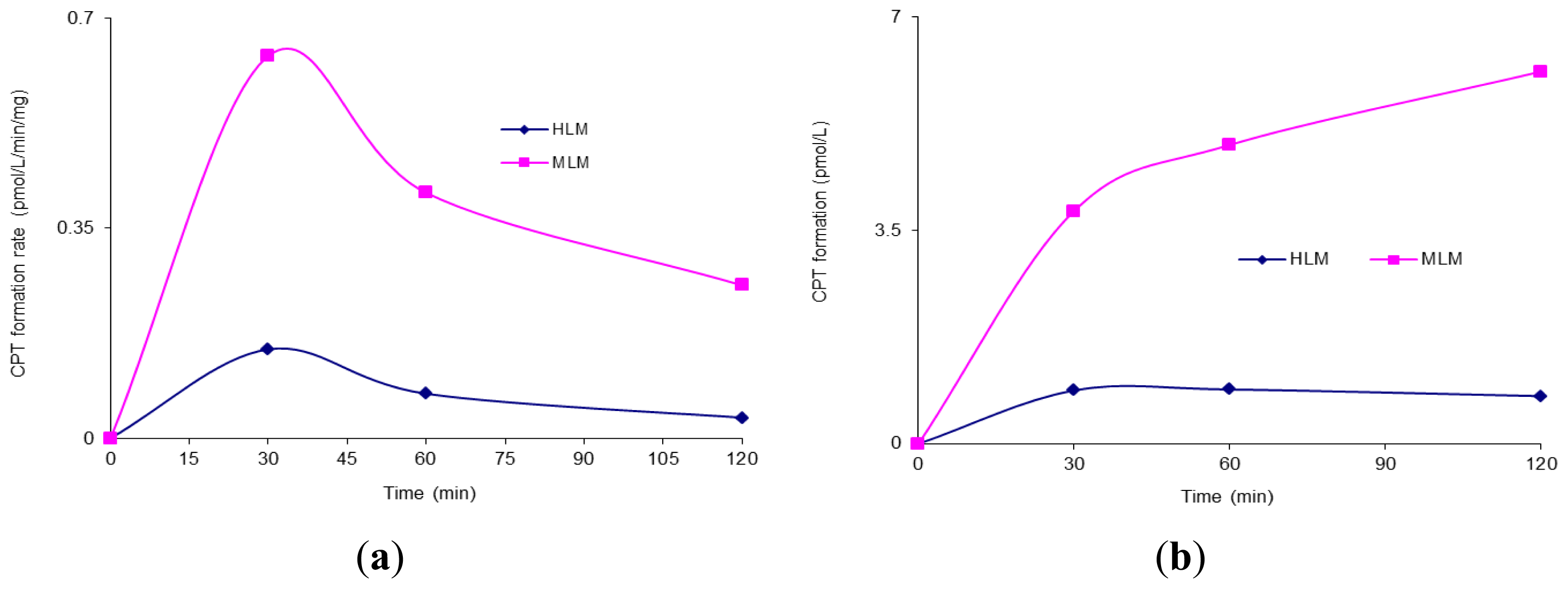
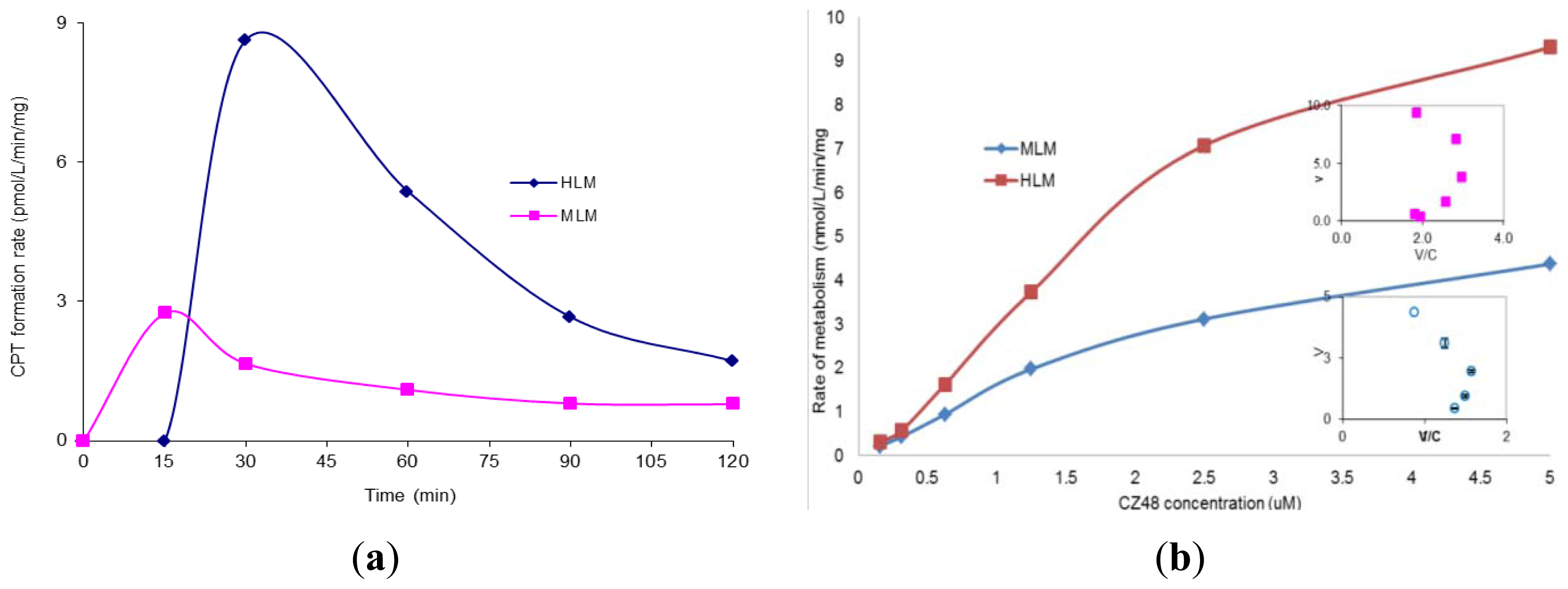
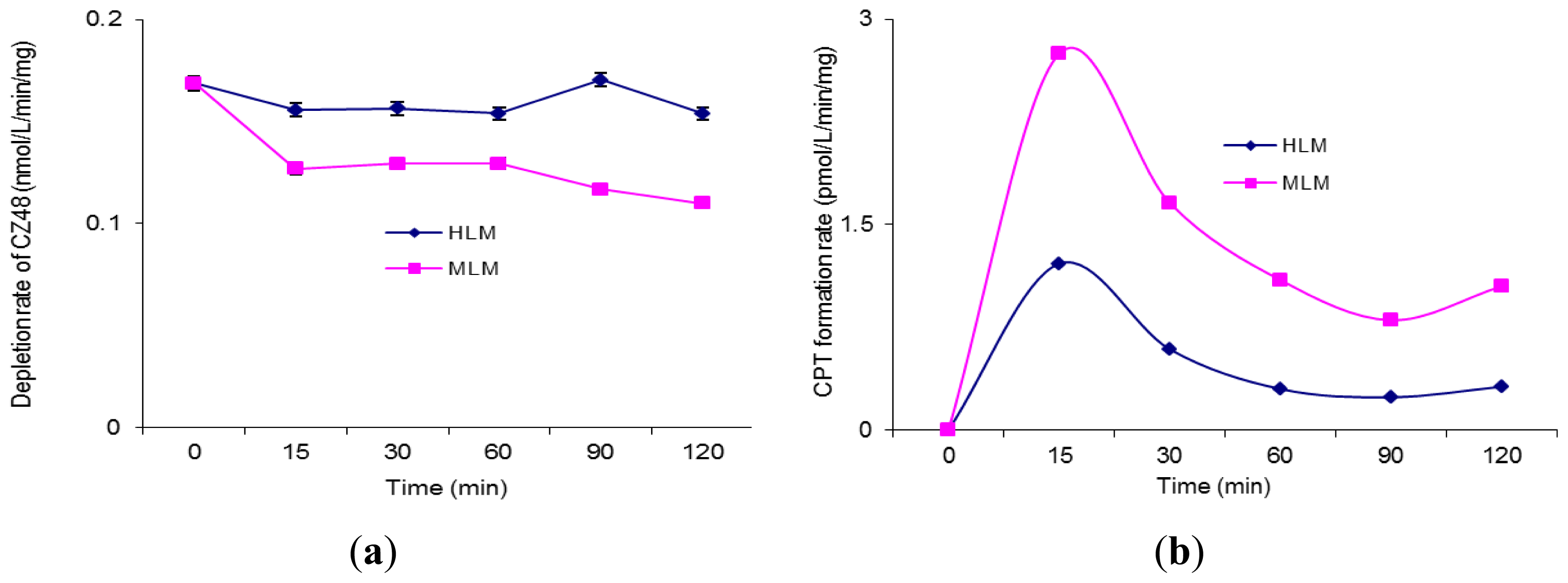
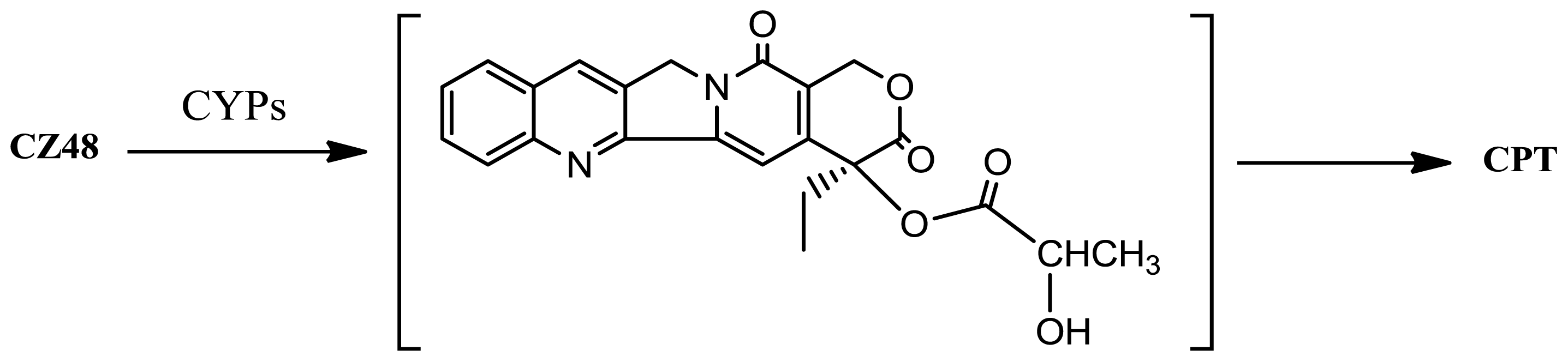
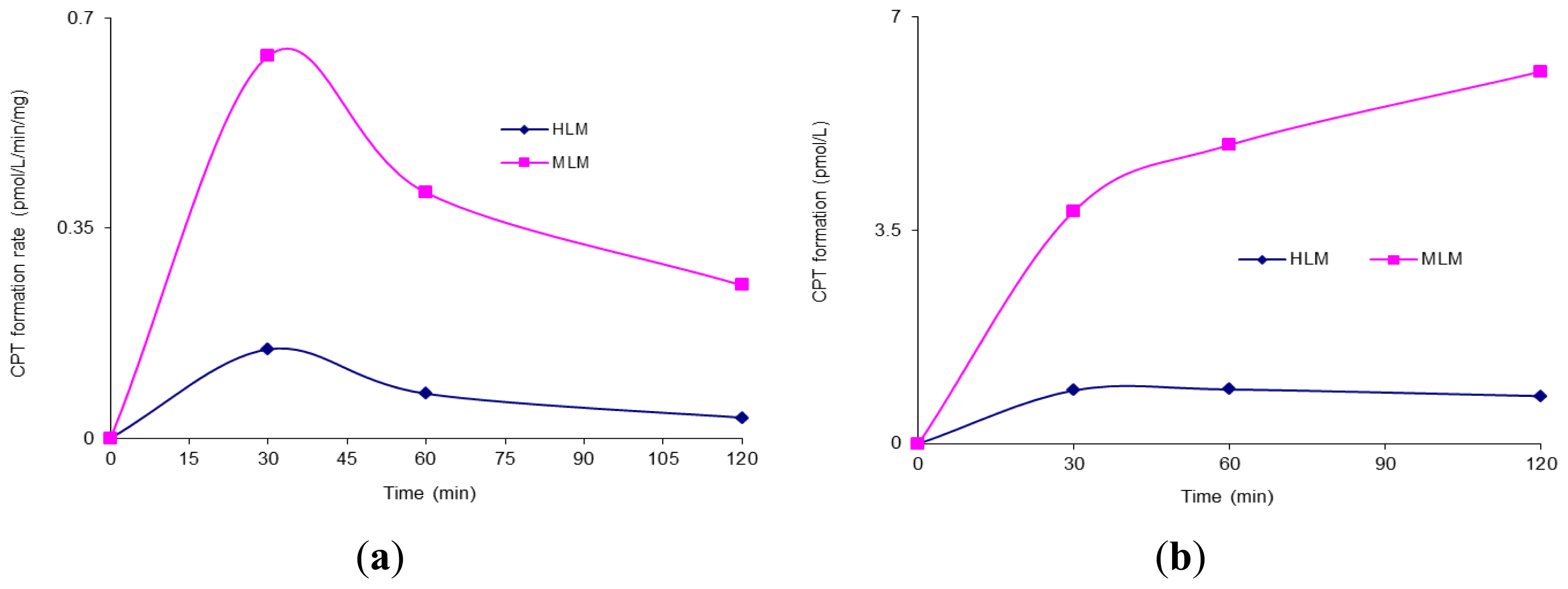
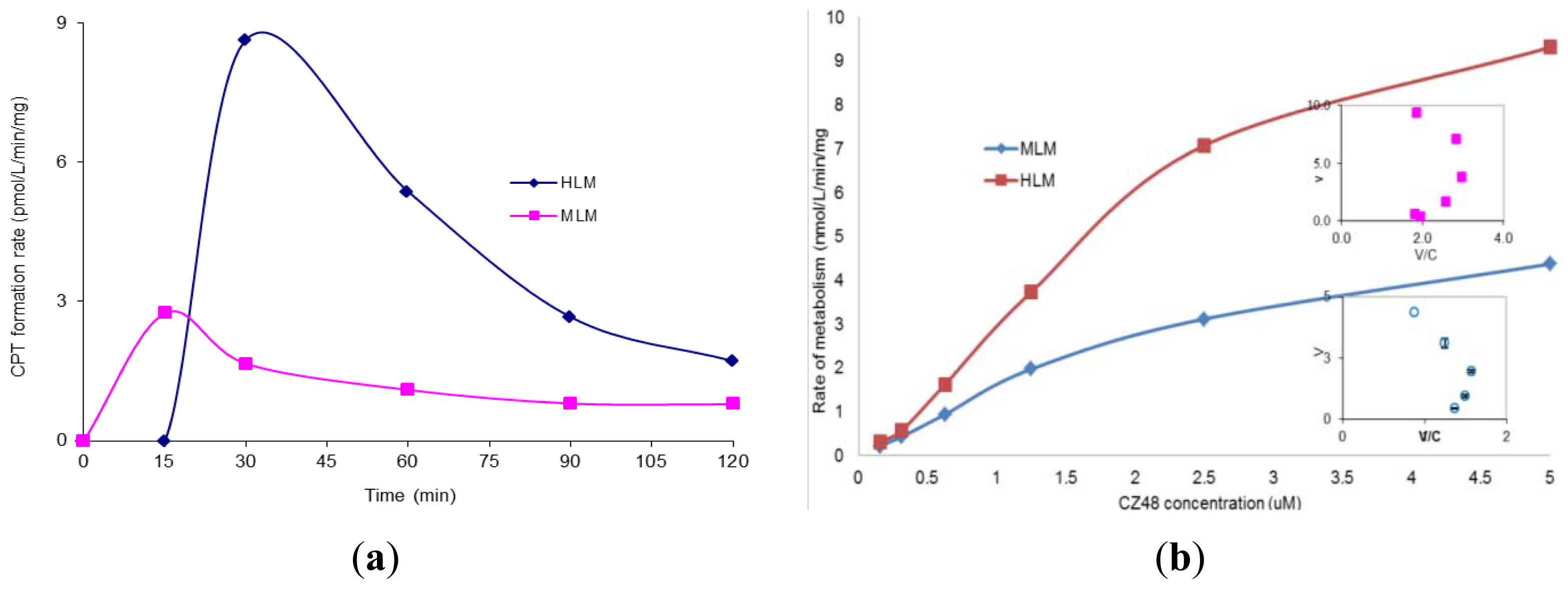
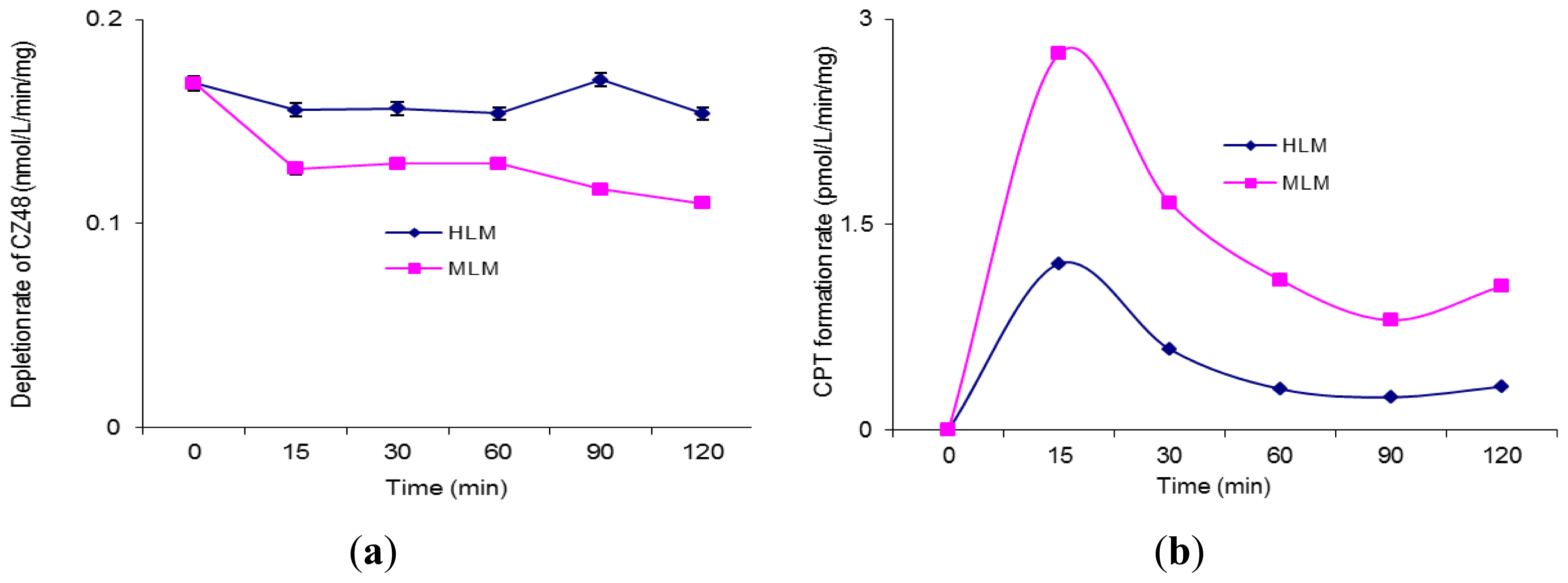
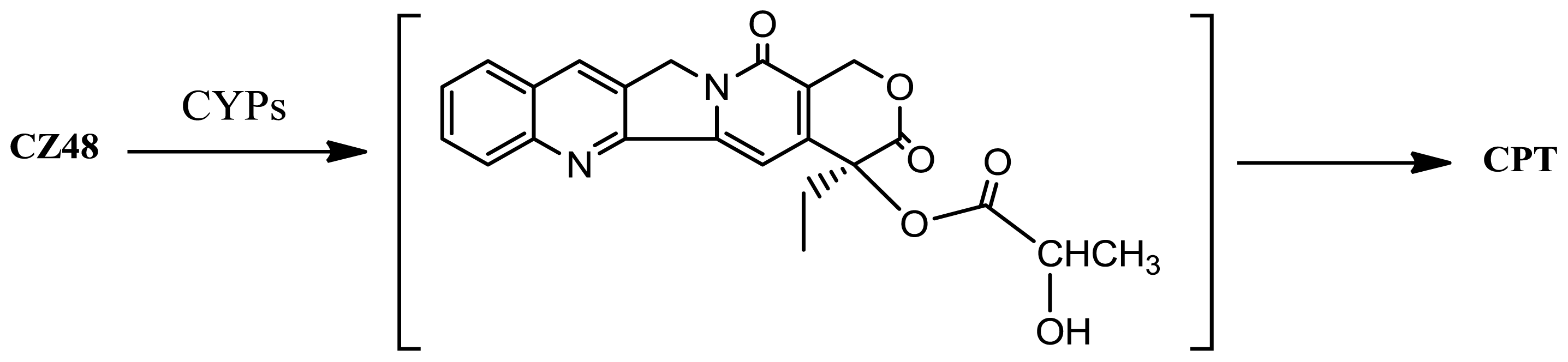
2. Results and Discussion
3. Experimental Section
3.1. Chemicals
3.2. CYP Activity
3.3. UGT Activity
3.4. Kinetic Data
4. Conclusion
Acknowledgement
- Conflict of InterestAuthors declare no conflict of interest.
References
- Guengerrich, F.P. Cytochrome P450 and chemical toxicity. Chem. Res. Toxicol 2008, 21, 70–83. [Google Scholar]
- Emoto, C.; Murayama, N.; Rostami-Hodjegan, A.; Yamazaki, H. Methodologies for investigating drug metabolism at the early drug discovery stage: Prediction of hepatic drug clearance and P450 contribution. Curr. Drug Metab 2010, 11, 678–685. [Google Scholar]
- Singh, S.S. Preclinical pharmacokinetics: An approach towards safer and efficacious drugs. Curr. Drug Metab 2006, 7, 165–182. [Google Scholar]
- Cao, Z.; Kozielski, A.; Liu, X.; Wang, Y.; Vardeman, D.; Giovanella, B. Crystalline camptothecin-20(S)-O-propionate hydrate: A novel anticancer agent with strong activity against 19 human tumor xenografts. Cancer Res 2009, 69, 4742–4749. [Google Scholar]
- Ludden, L.K.; Ludden, T.M.; Collins, J.M.; Pentikis, H.S.; Strong, J.M. Effect of albumin on the estimation, in vitro, of phenytoin Vmax and Km values: Implications for clinical correlation. J. Pharmacol. Exp. Ther 1997, 282, 391–396. [Google Scholar]
- Vickers, A.E.; Fischer, V.; Connors, S.; Fisher, R.L.; Baldeck, J.P.; Maurer, G.; Brendel, K. Cyclosporin A metabolism in human liver, kidney, and intestine slices: Comparision to rat and dog slices and human cell lines. Drug Metab. Dispos 1992, 20, 802–809. [Google Scholar]
- Vickers, A.E.; Connors, S.; Zollinger, M.; Biggi, W.A.; Larrauri, A.; Vogellaar, J.P.; Brendel, K. The biotransformation of the ergot derivative CQA 206-291 in human, dog and rat liver slice cultures and prediction of in vivo plasma clearance. Drug Metab. Dispos 1993, 21, 454–459. [Google Scholar]
- Lin, J.; Lu, A.Y.H. Inhibition and induction of P450 and the clinical implications. Clin. Pharmacokinet 1998, 35, 361–390. [Google Scholar]
- Cao, Z.; Harris, N.; Kozielski, A.; Vardeman, D.; Stehlin, J.; Giovanella, B. Alkyl esters of camptothecin and 9-nitrocamptothecin: Synthesis, in vitro pharmacokinetics, toxicity, and antitumor activity. J. Med. Chem 1998, 41, 31–37. [Google Scholar]
- Burke, T.G. Chemistry of the camptothecins in the bloodstream: Drug stabilization and optimization of activity in the camptothecins—From discovery to patients. Ann. N. Y. Acad. Sci 1993, 803, 29–31. [Google Scholar]
- Wani, M.C.; Ronman, P.E.; Lindley, L.T.; Wall, M.E. Plant antitumor agents. 18. Synthesis and biological activity of camptothecin analogs. J. Med. Chem 1980, 23, 554–560. [Google Scholar]
- Cao, Z. Sulfuric acid-catalyzed preparation of camptothecin esters. Open J. Med. Chem 2012, in press. [Google Scholar]
- Fujino, H.; Yamada, I.; Shimada, S.; Yoneda, M.; Kojima, J. Metabolic fate of Pitavastatin, a new inhibitor of HMG-CoA reductase: Human UDP-glucuronosyltransferase enzymes involved in lactonization. Xenobiotica 2003, 33, 27–41. [Google Scholar]
- Fujino, H.; Saito, T.; Tsunenari, Y.; Kojima, J.; Sakaeda, T. Metabolic properties of the acid and lactone form of HMG-CoA reductase inhibitors. Xenobiotica 2004, 34, 961–971. [Google Scholar]
- Liu, X.; Wang, Y.; Vardeman, D.; Cao, Z.; Giovanella, B. Development and validation of a reversed-phase HPLC with fluorescence detector method for simultaneous determination of CZ48 and its active metabolite camptothecin in mouse plasma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci 2008, 867, 84–89. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.; DeJesus, A.; Cao, Z.; Vardeman, D.; Giovanella, B. Metabolic Difference of CZ48 in Human and Mouse Liver Microsomes. Int. J. Mol. Sci. 2012, 13, 5498-5505. https://doi.org/10.3390/ijms13055498
Liu X, DeJesus A, Cao Z, Vardeman D, Giovanella B. Metabolic Difference of CZ48 in Human and Mouse Liver Microsomes. International Journal of Molecular Sciences. 2012; 13(5):5498-5505. https://doi.org/10.3390/ijms13055498
Chicago/Turabian StyleLiu, Xing, Albert DeJesus, Zhisong Cao, Dana Vardeman, and Beppino Giovanella. 2012. "Metabolic Difference of CZ48 in Human and Mouse Liver Microsomes" International Journal of Molecular Sciences 13, no. 5: 5498-5505. https://doi.org/10.3390/ijms13055498
APA StyleLiu, X., DeJesus, A., Cao, Z., Vardeman, D., & Giovanella, B. (2012). Metabolic Difference of CZ48 in Human and Mouse Liver Microsomes. International Journal of Molecular Sciences, 13(5), 5498-5505. https://doi.org/10.3390/ijms13055498