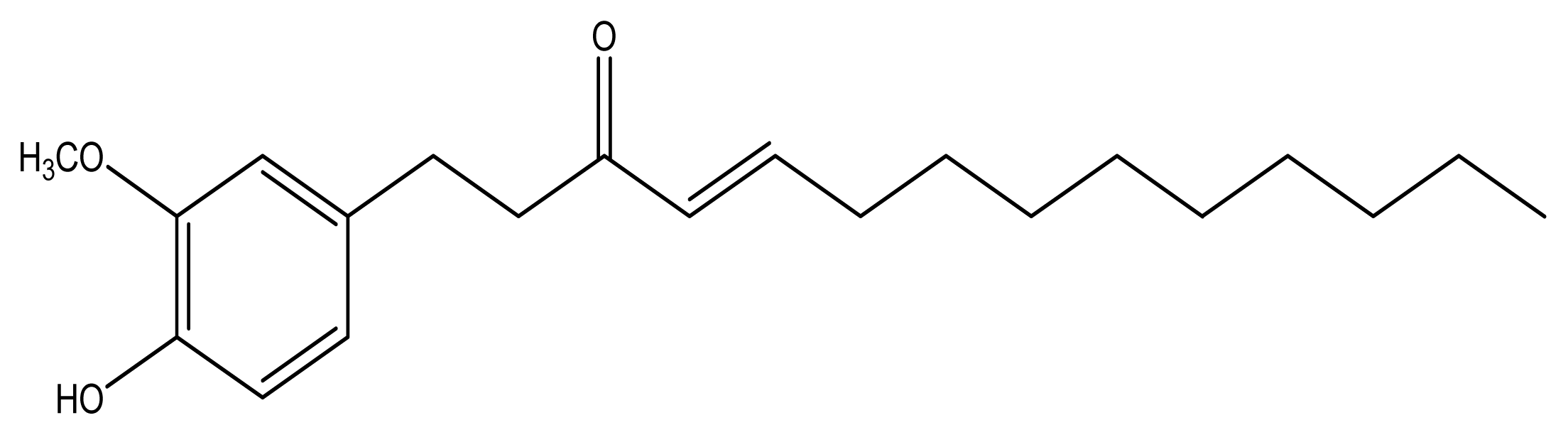
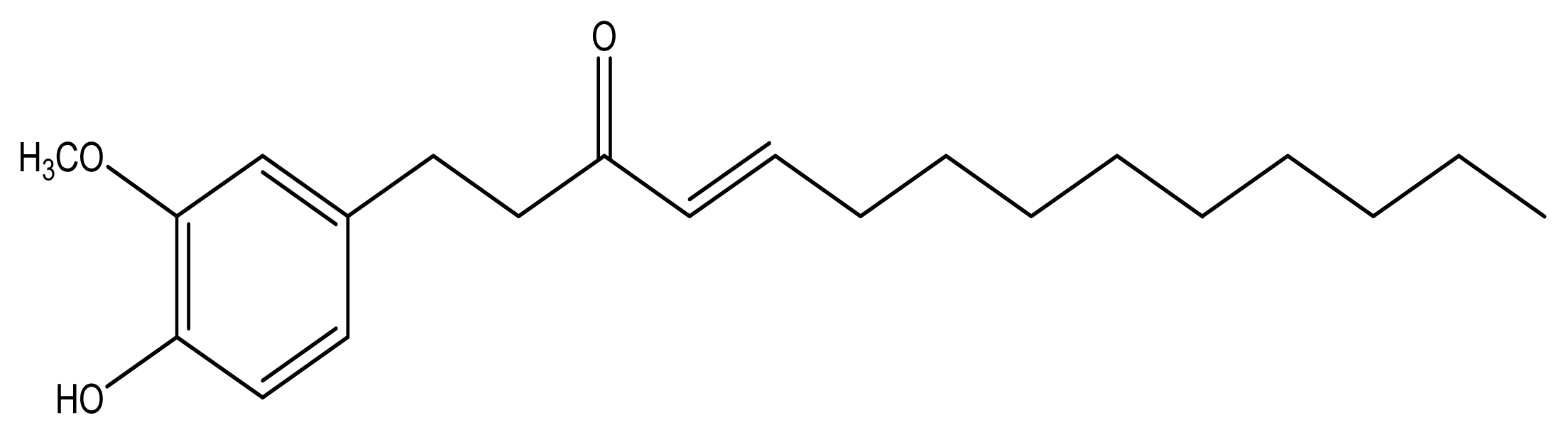
10-Shogaol, an Antioxidant from Zingiber officinale for Skin Cell Proliferation and Migration Enhancer



Abstract
:1. Introduction
2. Results and Discussion
2.1. Antioxidant Activities
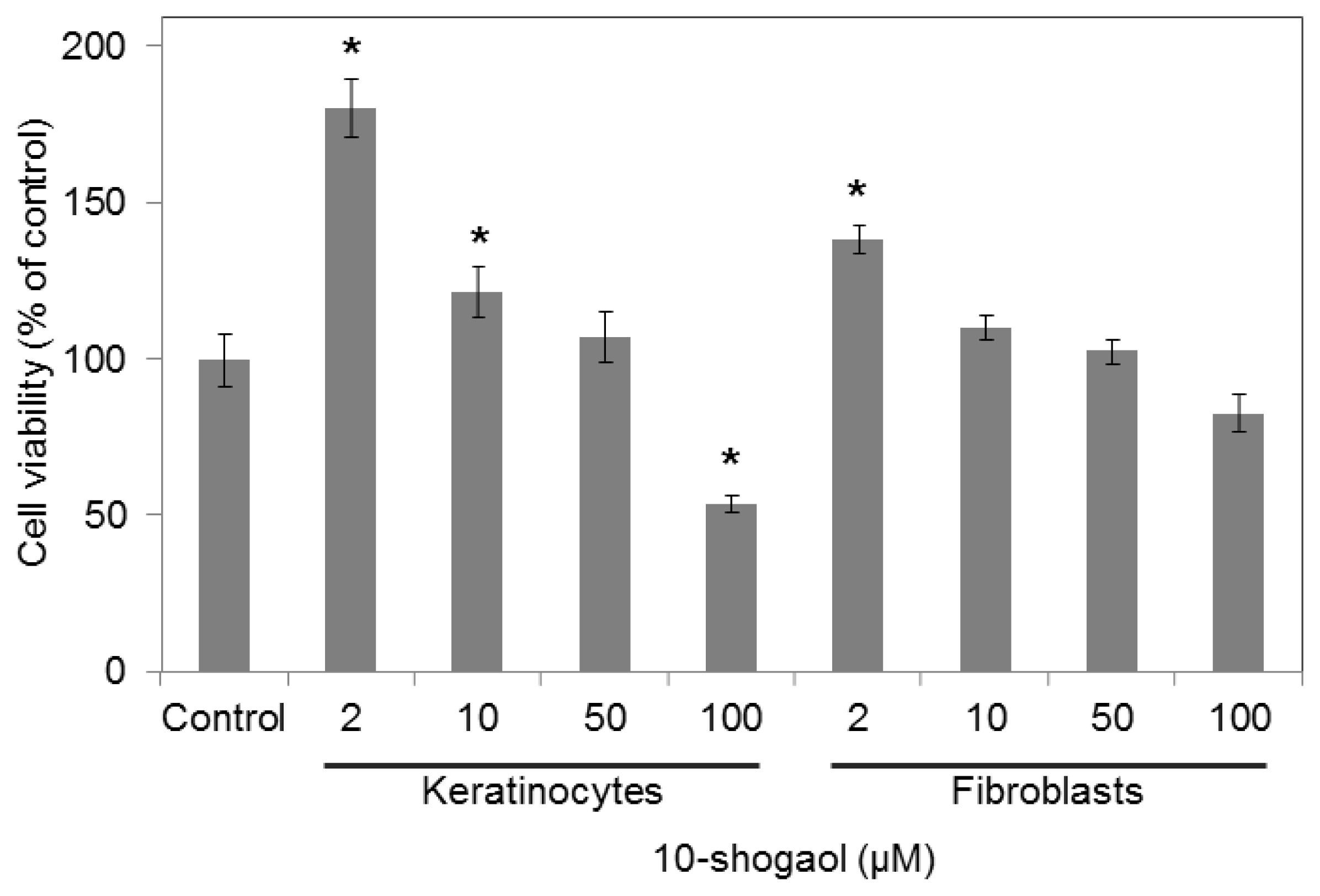
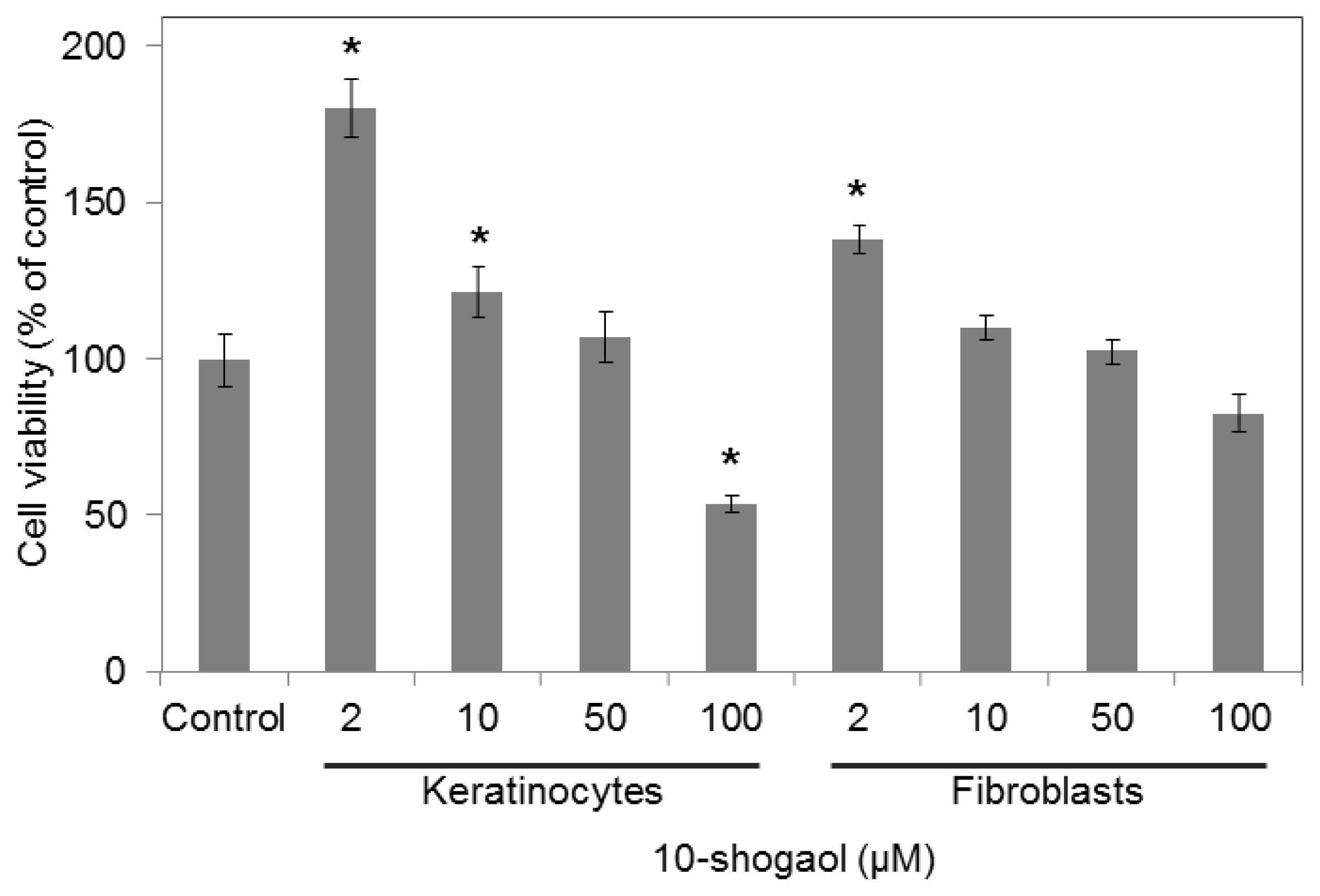
2.2. Cell Viabilities
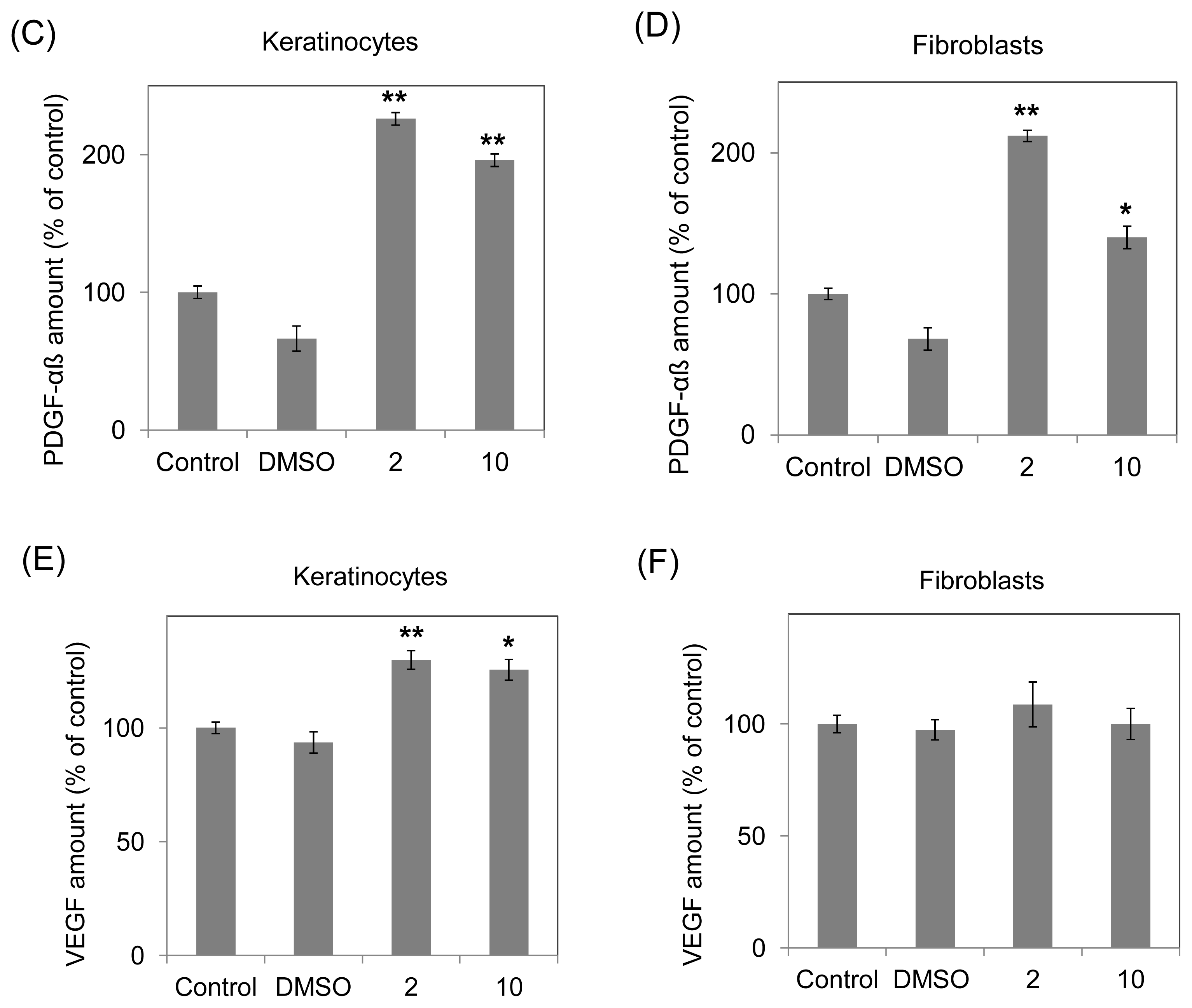
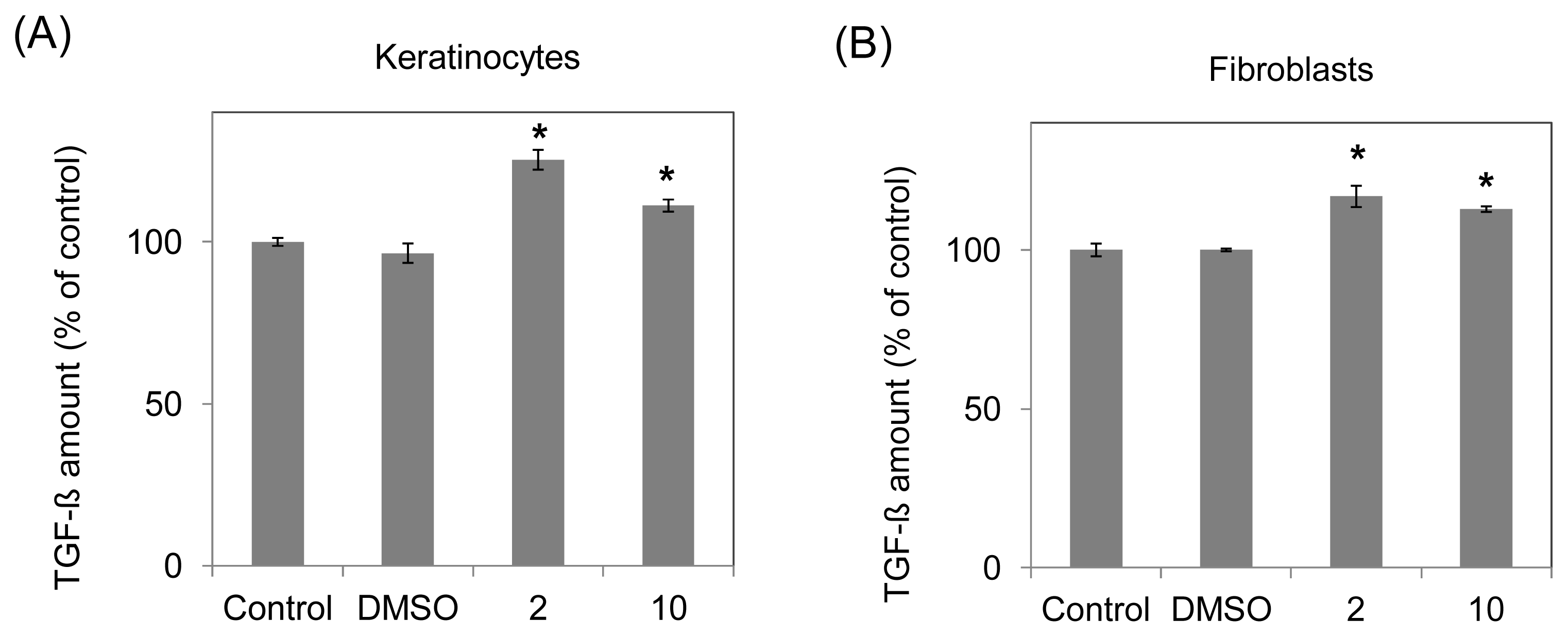
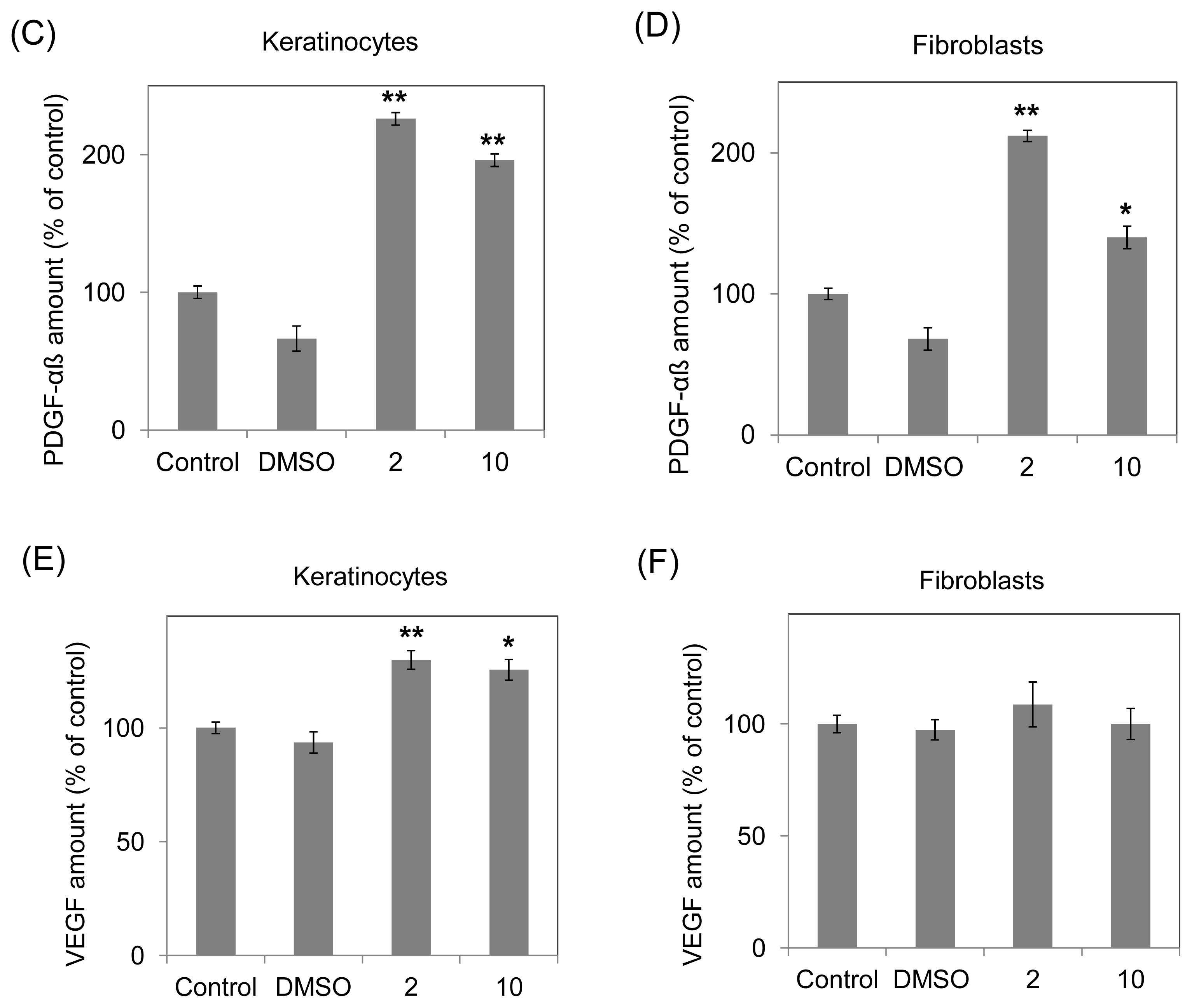
2.3. Increasing Growth Factor Production (TGF-β, PDGF-αβ and VEGF) of Human Fibroblasts and Keratinocytes by 10-Shogaol
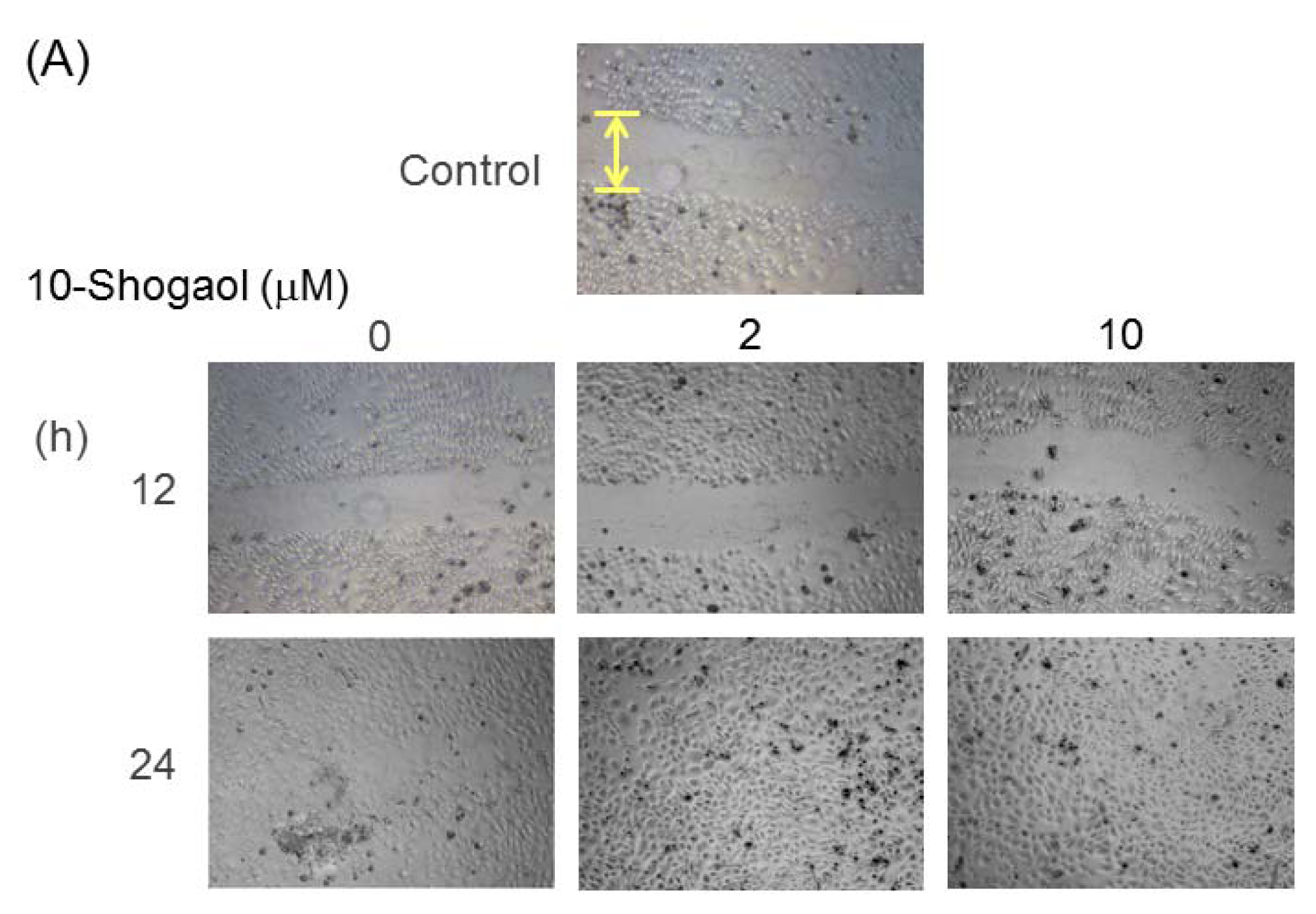
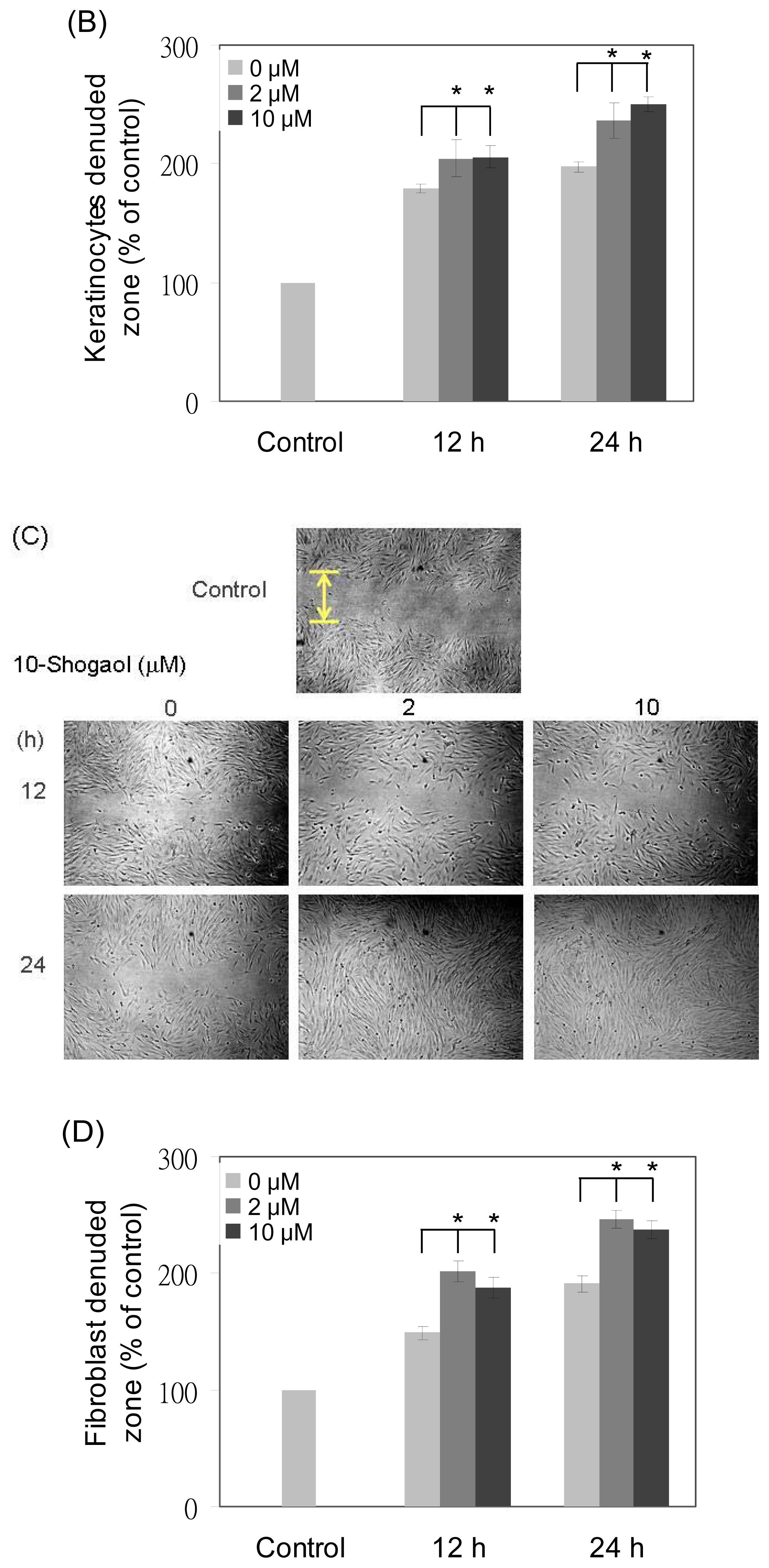
2.4. Cell Migrations of Fibroblasts and Keratinocytes Enhanced by 10-Shogaol
3. Experimental Section
3.1. Materials
3.2. Plant Materials
3.3. Determination of DPPH Radical Scavenging Capacity
3.4. Metal Chelating Activity
3.5. Reducing Power Assay
3.6. Human Dermal Fibroblasts and Epidermal Keratinocytes Cultures
3.7. Cell Growth Assay
3.8. ELISA Assays
3.9. In Vitro Wound Healing Assay
3.10. Statistical Analysis
4. Conclusions
Acknowledgements
Abbreviations
| BHA | 3-tert-butyl-4-hydroxyanisole |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | dimethyl sulfoxide |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| EDTA | ethylene diamine tetra-acetic acid |
| FBS | fetal bovine serum |
| PDGF-αβ | platelet derived growth factor-αβ |
| K3Fe(CN)6 | potassium ferricyanide |
| TGF-β | transforming growth factor-β |
| VEGF | vascular endothelial growth factors |
References
- Chen, B.H.; Chang, H.W.; Huang, H.M.; Chong, I.W.; Chen, J.S.; Chen, C.Y.; Wang, H.M. (−)-Anonaine induces oxidative stress and DNA damage to inhibit growth and migration of human lung carcinoma H1299 cells. J. Agric. Food Chem. 2011, 59, 2284–2290. [Google Scholar]
- Wang, H.M.; Chou, Y.T.; Hong, Z.L.; Chen, H.A.; Chang, Y.C.; Yang, W.L.; Chang, H.C.; Mai, C.T.; Chen, C.Y. Bioconstituents from stems of Synsepalum dulcificum Daniell (Sapotaceae) inhibit human melanoma proliferation, reduce mushroom tyrosinase activity and to be antioxidants. J. Taiwan Inst. Chem. Eng 2011, 42, 204–211. [Google Scholar]
- Wang, H.M.; Pan, J.L.; Chiu, C.C.; Chen, C.Y.; Yang, M.H.; Chang, J.S. Identification of anti-lung cancer extract from Chlorella vulgaris C-C by antioxidant property using supercritical carbon dioxide extraction. Process Biochem 2010, 45, 1865–1872. [Google Scholar]
- Wang, H.M.; Chen, C.Y.; Chen, H.A.; Huang, W.C.; Lin, W.R.; Chen, T.C.; Lin, C.Y.; Chien, H.S.; Lu, P.L.; Lin, C.M.; Chen, Y.H. Zingiber officinale (ginger) compounds with tetracycline have synergistic effects against clinical extensively-drug resistant Acinetobacter baumannii. Phytother. Res 2010, 24, 1825–1830. [Google Scholar]
- Chen, B.H.; Wu, P.Y.; Chen, K.M.; Fu, T.Z.; Wang, H.M.; Chen, C.Y. Antiallergic potential on RBL-2H3 cells of some phenolic constituents of Zingiber officinale (Ginger). J. Nat. Prod 2009, 72, 950–953. [Google Scholar]
- Waddington, R.J.; Alraies, A.; Colombo, J.S.; Sloan, A.J.; Okazaki, J.; Moseley, R. Characterization of oxidative stress status during diabetic bone healing. Cells Tissues Organs 2011, 194, 307–312. [Google Scholar]
- Mladenović, M.; Mihailović, M.; Bogojević, D.; Matić, S.; Nićiforović, N.; Mihailović, V.; Vuković, N.; Sukdolak, S.; Solujić, S. In vitro antioxidant activity of selected 4-hydroxy-chromene-2-one derivatives—SAR, QSAR and DFT studies. Int. J. Mol. Sci 2011, 12, 2822–2841. [Google Scholar]
- Zhang, H.; Wang, Z.Y.; Yang, L.; Yang, X.; Wang, X.; Zhang, Z. In vitro antioxidant activities of sulfated derivatives of polysaccharides extracted from Auricularia auricular. Int. J. Mol. Sci 2011, 12, 3288–3302. [Google Scholar]
- Fu, L.; Xu, B.T.; Gan, R.Y.; Zhang, Y.; Xu, X.R.; Xia, E.Q.; Li, H.B. Total phenolic contents and antioxidant capacities of herbal and tea infusions. Int. J. Mol. Sci 2011, 12, 2112–2124. [Google Scholar]
- Wei, S.D.; Zhou, H.C.; Lin, Y.M. Antioxidant activities of extract and fractions from the hypocotyls of the mangrove plant Kandelia candel. Int. J. Mol. Sci 2010, 11, 4080–4093. [Google Scholar]
- Amjad, S.B.; Carachi, R.; Edward, M. Keratinocyte regulation of TGF-beta and connective tissue growth factor expression: A role in suppression of scar tissue formation. Wound Repair Regen 2007, 15, 748–755. [Google Scholar]
- Rolfe, K.J.; Richardson, J.; Vigor, C.; Irvine, L.M.; Grobbelaar, A.O.; Linge, C. A role for TGF-β1-induced cellular responses during wound healing of the non-scarring early human fetus? J. Invest. Dermatol 2007, 127, 2656–2667. [Google Scholar]
- Trompezinski, S.; Berthier-Vergnes, O.; Denis, A.; Schmitt, D.; Viac, J. Comparative expression of vascular endothelial growth factor family members, VEGF-B, -C and -D, by normal human keratinocytes and fibroblasts. Exp. Dermatol 2004, 13, 98–105. [Google Scholar]
- Stavri, G.T.; Hong, Y.; Zachary, I.C.; Breier, G.; Baskerville, P.A.; Yla-Herttuala, S.; Risau, W.; Martin, J.F.; Erusalimsky, J.D. Hypoxia and platelet-derived growth factor-BB synergistically upregulate the expression of vascular endothelial growth factor in vascular smooth muscle cells. FEBS Lett 1995, 358, 311–315. [Google Scholar]
- Afzal, M.; Al-Hadidi, D.; Menon, M.; Pesek, J.; Dhami, M.S. Ginger: An ethnomedical, chemical and pharmacological review. Drug Metabol. Drug Interact 2001, 18, 159–190. [Google Scholar]
- Wohlmuth, H.; Smith, M.K.; Brooks, L.O.; Myers, S.P.; Leach, D.N. Essential oil composition of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe) grown in Australia. J. Agric. Food Chem 2006, 54, 1414–1419. [Google Scholar]
- Shao, X.; Lv, L.; Parks, T.; Wu, H.; Ho, C.T.; Sang, S. Quantitative analysis of ginger components in commercial products using liquid chromatography with electrochemical array detection. J. Agric. Food Chem 2010, 58, 12608–12614. [Google Scholar]
- Ling, H.; Yang, H.; Tan, S.H.; Chui, W.K.; Chew, E.H. 6-Shogaol, an active constituent of ginger, inhibits breast cancer cell invasion by reducing matrix metalloproteinase-9 expression via blockade of nuclear factor-κB activation. Br. J. Pharmacol 2010, 161, 1763–1777. [Google Scholar]
- Gan, F.F.; Nagle, A.A.; Ang, X.; Ho, O.H.; Tan, S.H.; Yang, H.; Chui, W.K.; Chew, E.H. Shogaols at proapoptotic concentrations induce G2/M arrest and aberrant mitotic cell death associated with tubulin aggregation. Apoptosis 2011, 16, 856–867. [Google Scholar]
- Iwasaki, Y.; Morita, A.; Iwasawa, T.; Kobata, K.; Sekiwa, Y.; Morimitsu, Y.; Kubota, K.; Watanabe, T. A nonpungent component of steamed ginger—[10]-shogaol—increases adrenaline secretion via the activation of TRPV1. Nutr. Neurosci 2006, 9, 169–178. [Google Scholar]
- Wu, L.; Wang, G.; Tang, S.; Long, G.; Yin, T. Protection of endothelial cells, inhibition of neointimal hyperplasia by β-elemene in an injured artery. Cardiovasc. Drugs Ther 2011, 25, 233–242. [Google Scholar]
- Panchatcharam, M.; Miriyala, S.; Gayathri, V.S.; Suguna, L. Curcumin improves wound healing by modulating collagen and decreasing reactive oxygen species. Mol. Cell Biochem 2006, 290, 87–96. [Google Scholar]
- Bhagavathula, N.; Warner, R.L.; DaSilva, M.; McClintock, S.D.; Barron, A.; Aslam, M.N.; Johnson, K.J.; Varani, J. A combination of curcumin and ginger extract improves abrasion wound healing in corticosteroid-impaired hairless rat skin. Wound Repair Regen 2009, 17, 360–366. [Google Scholar]
- Yoshimura, K.; Tsukamoto, K.; Okazaki, M.; Virador, V.M.; Lei, T.C.; Suzuki, Y.; Uchida, G.; Kitano, Y.; Harii, K. Effects of all-trans retinoic acid on melanogenesis in pigmented skin equivalents and monolayer culture of melanocytes. 2001, S68–75. [Google Scholar]
- Chen, C.Y.; Tai, C.J.; Cheng, J.T.; Zheng, J.J.; Chen, Y.Z.; Liu, T.Z.; Yiin, S.J.; Chern, C.L. 6-Dehydrogingerdione sensitizes human hepatoblastoma Hep G2 cells to TRAIL-induced apoptosis via reactive oxygen species-mediated increase of DR5. J. Agric. Food Chem 2010, 58, 5604–5611. [Google Scholar]
- Hsu, Y.L.; Chen, C.Y.; Hou, M.F.; Tsai, E.M.; Jong, Y.J.; Hung, C.H.; Kuo, P.L. 6-Dehydrogingerdione, an active constituent of dietary ginger, induces cell cycle arrest and apoptosis through reactive oxygen species/c-Jun N-terminal kinase pathways in human breast cancer cells. Mol. Nutr. Food Res 2010, 54, 1307–1317. [Google Scholar]
- Riedel, K.; Riedel, F.; Goessler, U.R.; Germann, G.; Sauerbier, M. TGF-β antisense therapy increases angiogenic potential in human keratinocytes in vitro. Arch. Med. Res 2007, 38, 45–51. [Google Scholar]
- Lindahl, P.; Johansson, B.R.; Leveen, P.; Betsholtz, C. Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science 1997, 277, 242–245. [Google Scholar]
- Gerber, H.P.; Dixit, V.; Ferrara, N. Vascular endothelial growth factor induces expression of the antiapoptotic proteins Bcl-2 and A1 in vascular endothelial cells. J. Biol. Chem 1998, 273, 13313–13316. [Google Scholar]
- Panchatcharam, M.; Miriyala, S.; Gayathri, V.S.; Suguna, L. Curcumin improves wound healing by modulating collagen and decreasing reactive oxygen species. Mol. Cell Biochem 2006, 290, 87–96. [Google Scholar]
- Lin, R.J.; Chen, C.Y.; Lee, J.D.; Lu, C.M.; Chung, L.Y.; Yen, C.M. Larvicidal constituents of Zingiber officinale (Ginger) against Anisakis simplex. Planta Med 2010, 76, 1852–1858. [Google Scholar]
- Shoji, N.; Iwasa, A.; Takemoto, T.; Ishida, Y.; Ohizumi, Y. Cardiotonic principles of ginger (Zingiber officinale Roscoe). Pharm. Sci 1982, 71, 1174–1175. [Google Scholar]
- Wang, H.M.; Chen, C.Y.; Chen, C.Y.; Ho, M.L.; Chou, Y.T.; Chang, H.C.; Lee, C.H.; Wang, C.Z.; Chu, I.M. (−)-N-Formylanonaine from Michelia alba as human tyrosinase inhibitor and antioxidant. Bioorg. Med. Chem. 2010, 18, 5241–5247. [Google Scholar]
- Wang, H.M.; Chen, C.Y.; Wen, Z.H. Identifying melanogenesis inhibitors from Cinnamomum subavenium with in vitro and in vivo screening systems by targeting the human tyrosinase. Exp. Dermatol 2011, 20, 242–248. [Google Scholar]
- Wang, H.M.; Chiu, C.C.; Wu, P.F.; Chen, C.Y. Subamolide E from Cinnamomum subavenium induces sub G1 cell cycle arrest, caspase-dependent apoptosis, and reduces migration ability of human melanoma cells. J. Agric. Food Chem 2011, 59, 8187–8192. [Google Scholar]
- Geback, T.; Schulz, M.M.; Koumoutsakos, P.; Detmar, M. TScratch: a novel and simple software tool for automated analysis of monolayer wound healing assays. Biotechniques 2009, 46, 265–274. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | DPPH. Scavenging (%) | Metal Chelating (%) | Reducing Power (100 μM, OD700) |
|---|---|---|---|
| 10-Shogaol | 34.54 ± 0.02 | <10.00 | 0.60 ± 0.02 |
| Vitamin C a | 90.02 ± 0.40 | - | - |
| EDTA b | - | 94.78 ± 0.60 | - |
| BHA c | - | - | 0.98 ± 0.11 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, C.-Y.; Cheng, K.-C.; Chang, A.Y.; Lin, Y.-T.; Hseu, Y.-C.; Wang, H.-M. 10-Shogaol, an Antioxidant from Zingiber officinale for Skin Cell Proliferation and Migration Enhancer. Int. J. Mol. Sci. 2012, 13, 1762-1777. https://doi.org/10.3390/ijms13021762
Chen C-Y, Cheng K-C, Chang AY, Lin Y-T, Hseu Y-C, Wang H-M. 10-Shogaol, an Antioxidant from Zingiber officinale for Skin Cell Proliferation and Migration Enhancer. International Journal of Molecular Sciences. 2012; 13(2):1762-1777. https://doi.org/10.3390/ijms13021762
Chicago/Turabian StyleChen, Chung-Yi, Kuo-Chen Cheng, Andy Y Chang, Ying-Ting Lin, You-Cheng Hseu, and Hui-Min Wang. 2012. "10-Shogaol, an Antioxidant from Zingiber officinale for Skin Cell Proliferation and Migration Enhancer" International Journal of Molecular Sciences 13, no. 2: 1762-1777. https://doi.org/10.3390/ijms13021762
APA StyleChen, C.-Y., Cheng, K.-C., Chang, A. Y., Lin, Y.-T., Hseu, Y.-C., & Wang, H.-M. (2012). 10-Shogaol, an Antioxidant from Zingiber officinale for Skin Cell Proliferation and Migration Enhancer. International Journal of Molecular Sciences, 13(2), 1762-1777. https://doi.org/10.3390/ijms13021762