Proliferation-Attenuating and Apoptosis-Inducing Effects of Tryptanthrin on Human Chronic Myeloid Leukemia K562 Cell Line in Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
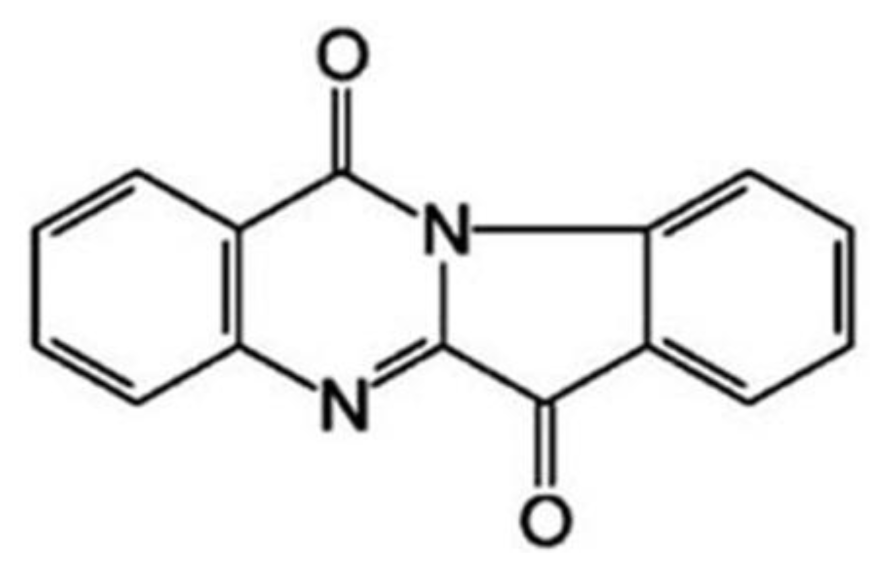
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Treatment
2.3. MTT Assay
2.4. Hoechst 33258 Fluorescent Staining
2.5. Transmission Electron Microscopy
2.6. Annexin-V/PI Staining, Apoptosis and Cell Cycle Determination by Flow Cytometry
2.7. Measurement of Mitochondrial Membrane Potential (Δψm)
2.8. Assay of Bcl-2, Bax, Cyt-c, Procaspase-3 and Caspase-3
2.9. Statistical Analysis
3. Results
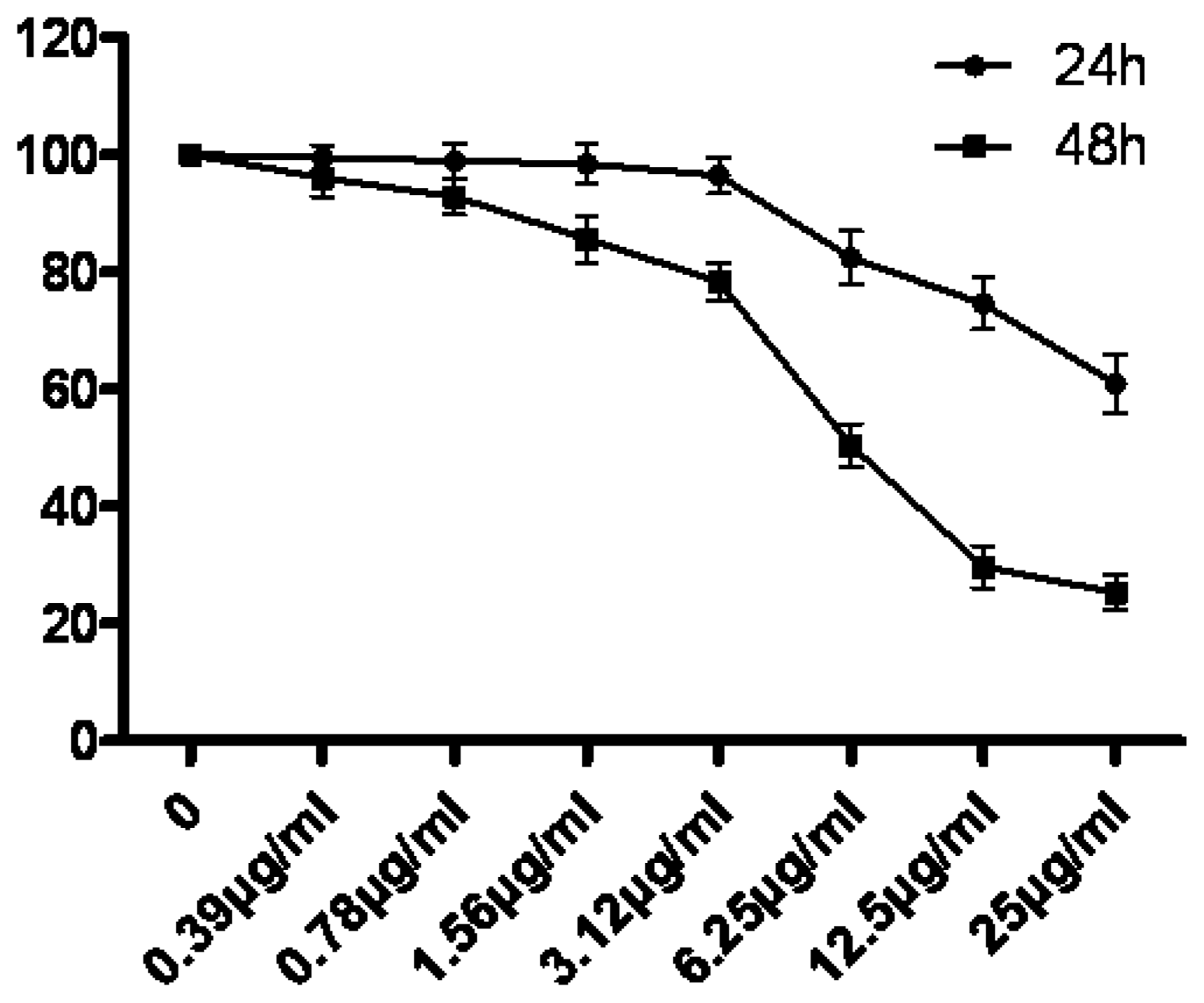
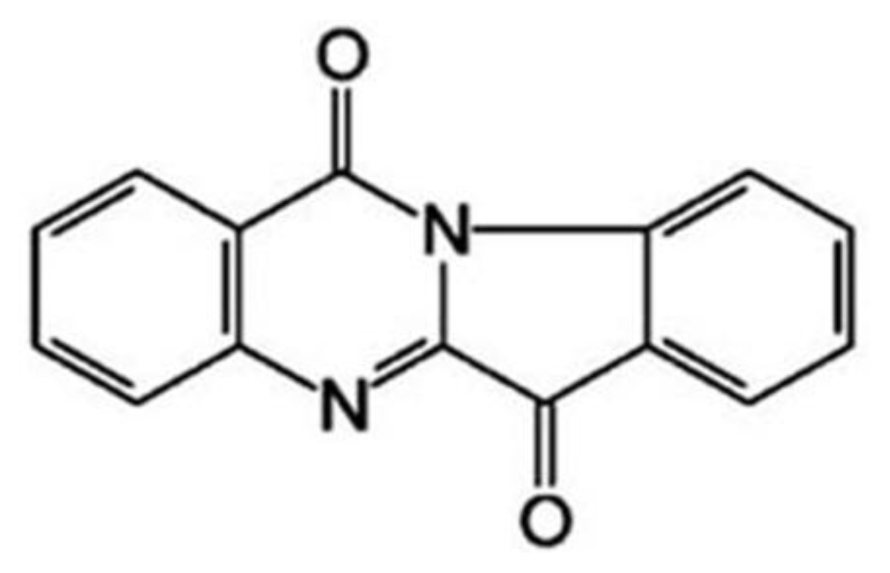
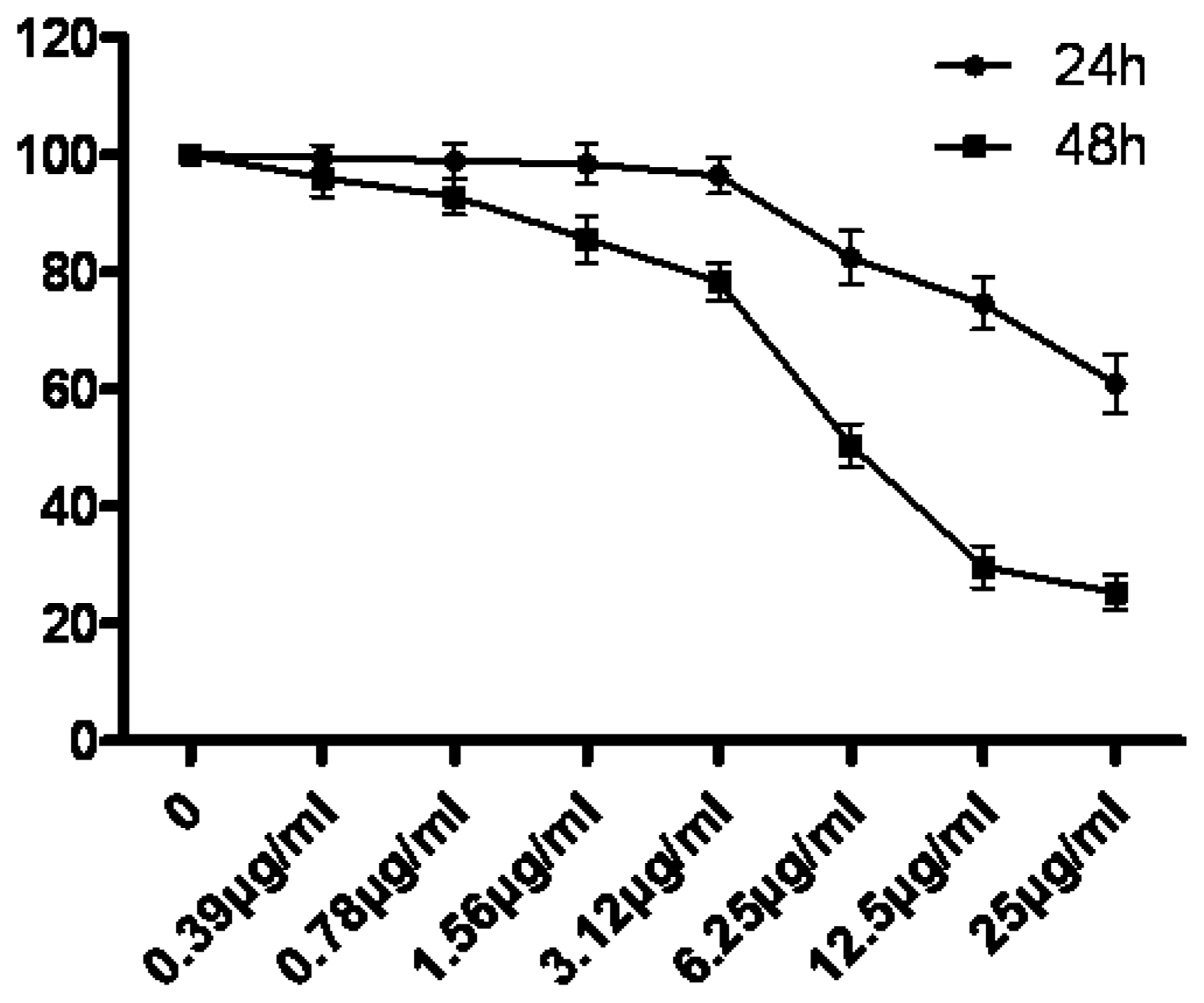
3.1. The Growth Suppression Effect of Tryptanthrin on K562 Cells
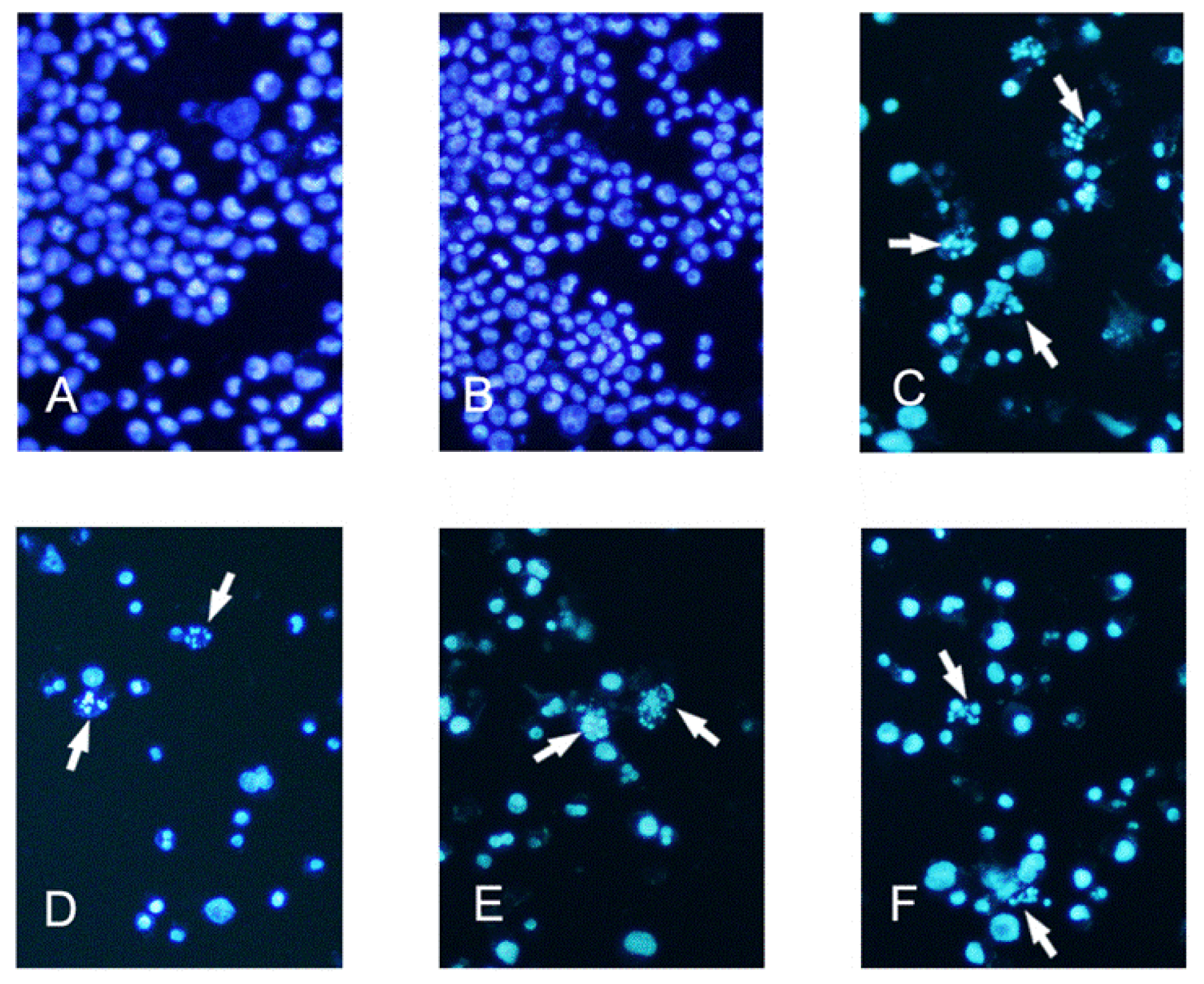
3.2. Determination of Apoptotic K562 Cells by Hoechst 33258 Staining
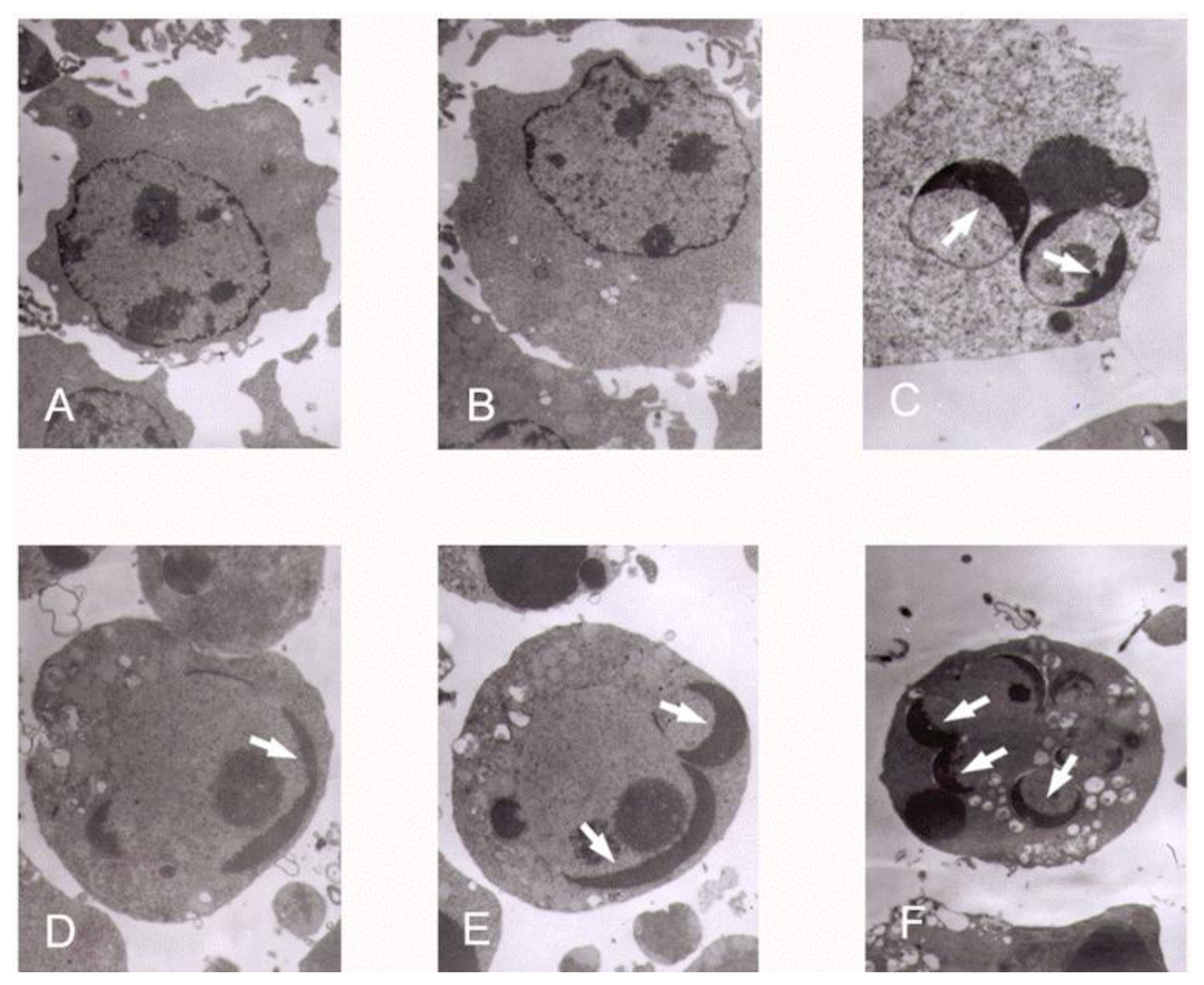
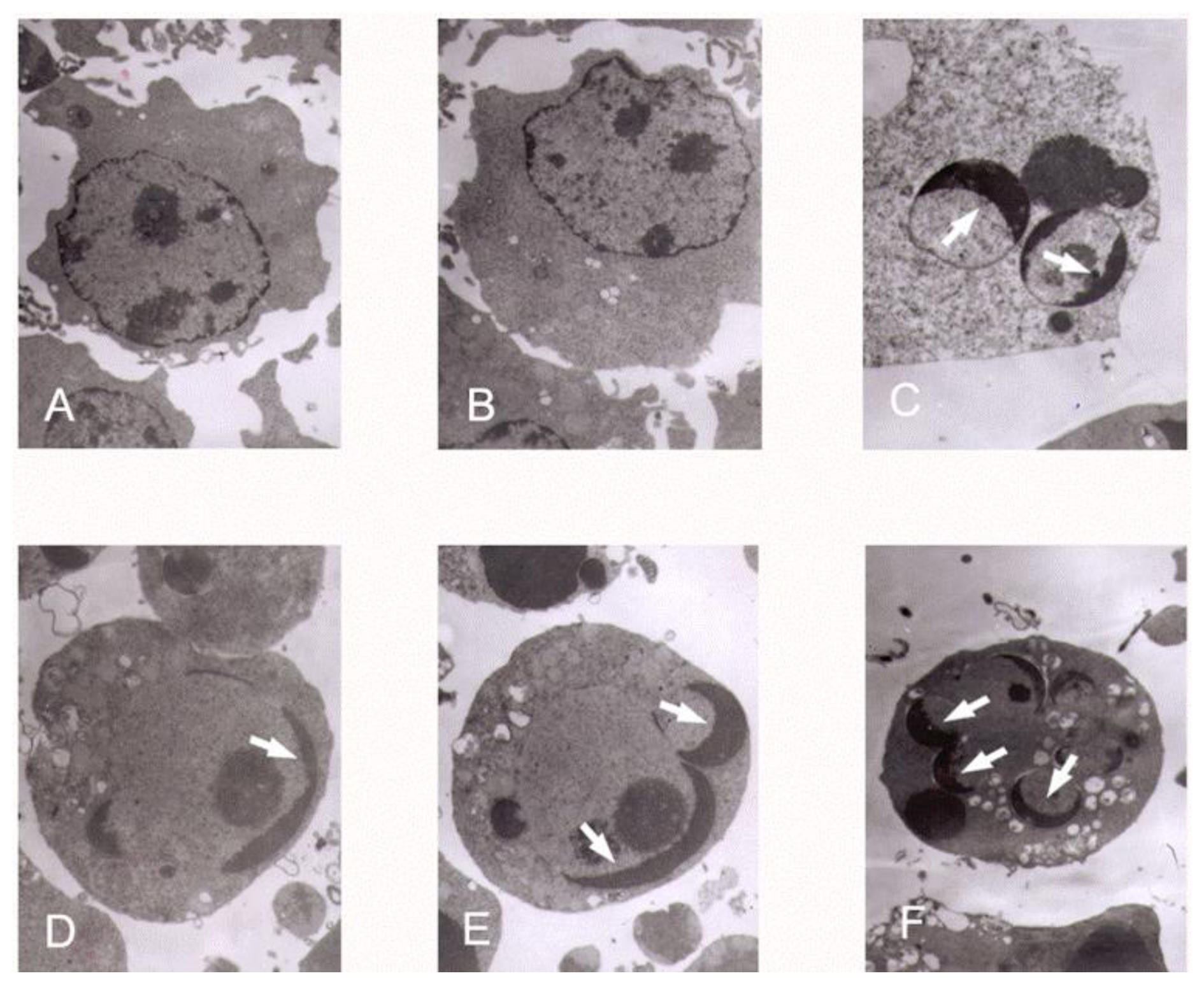
3.3. K562 Cells Ultra Structure
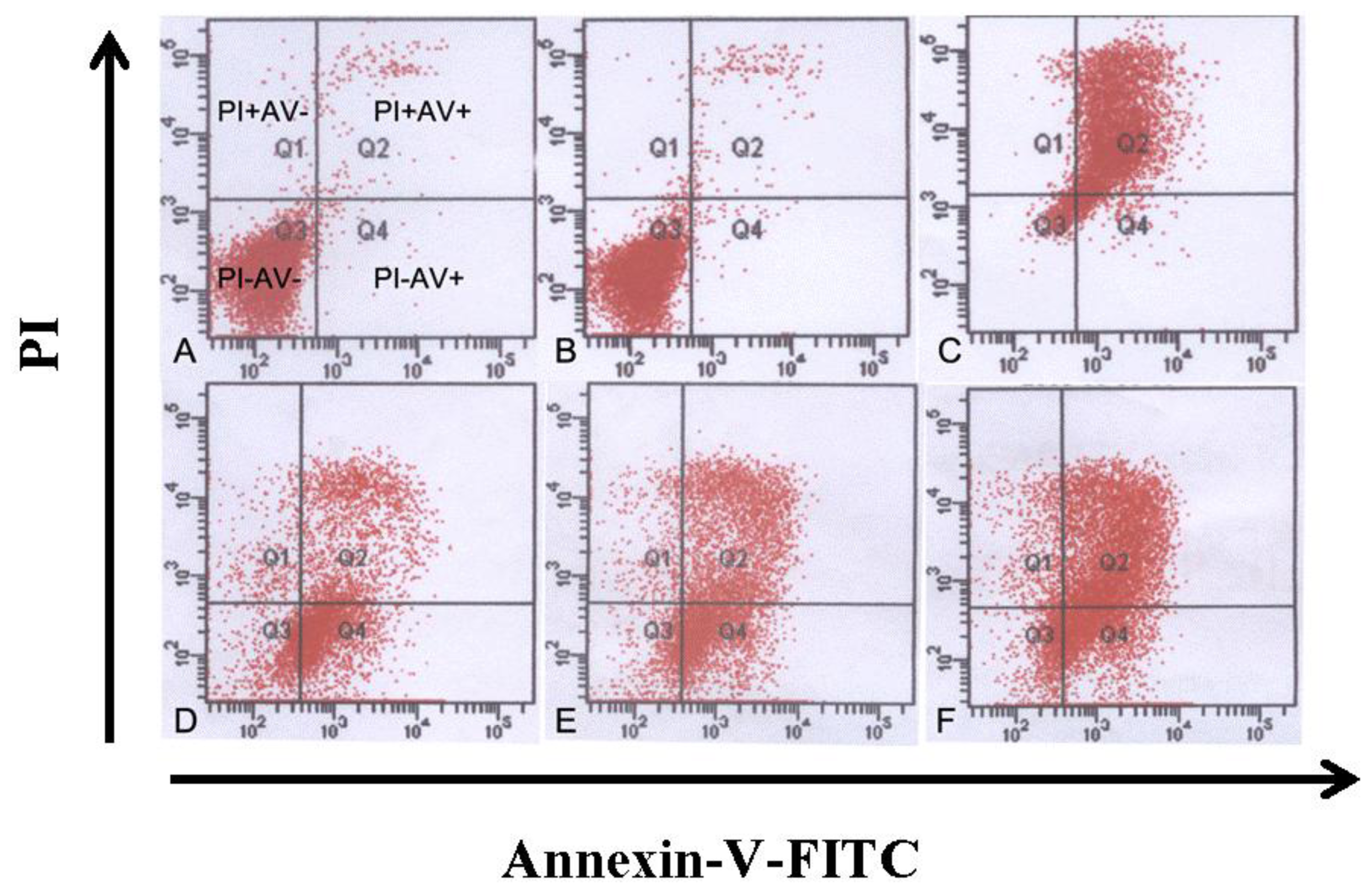
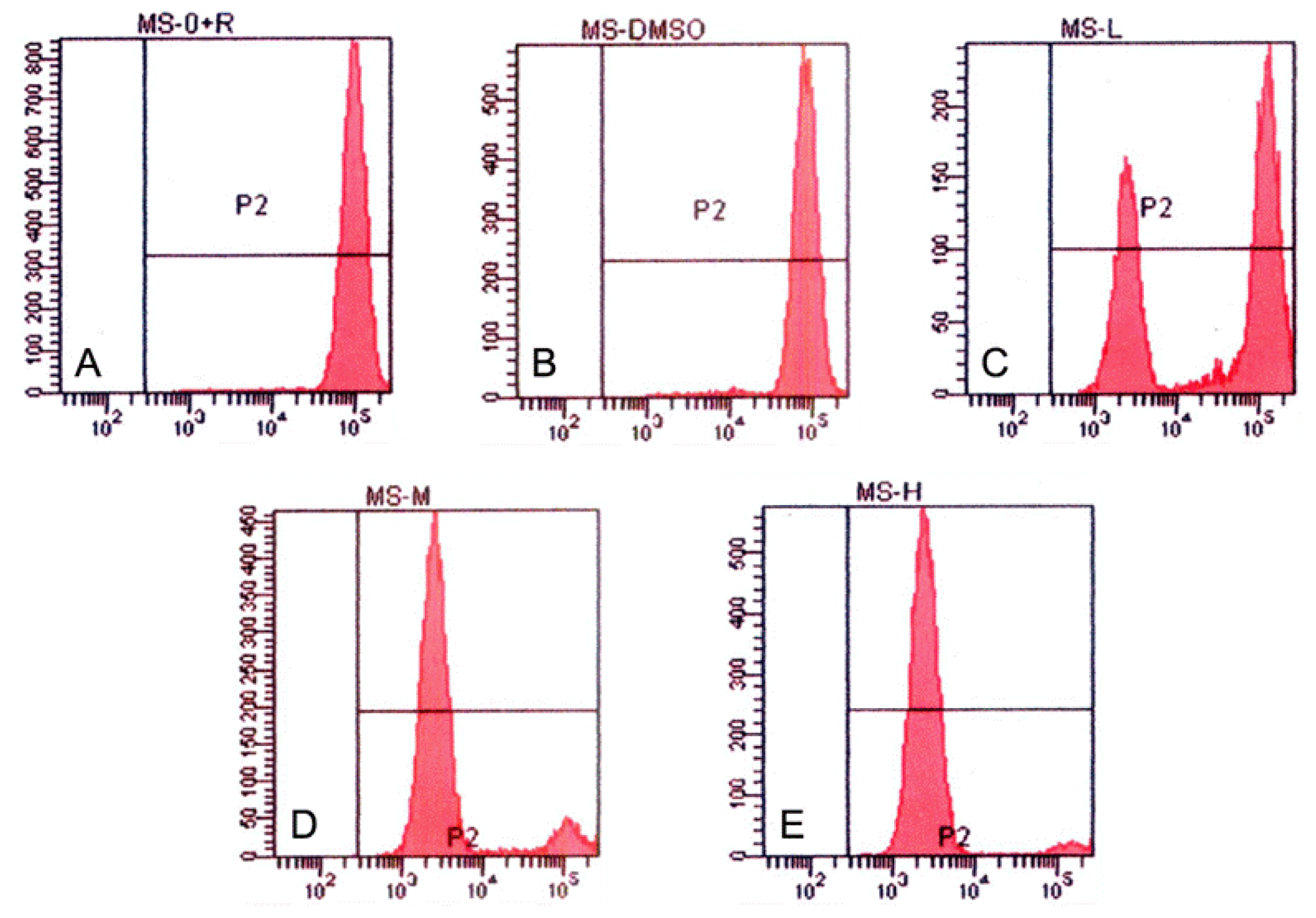
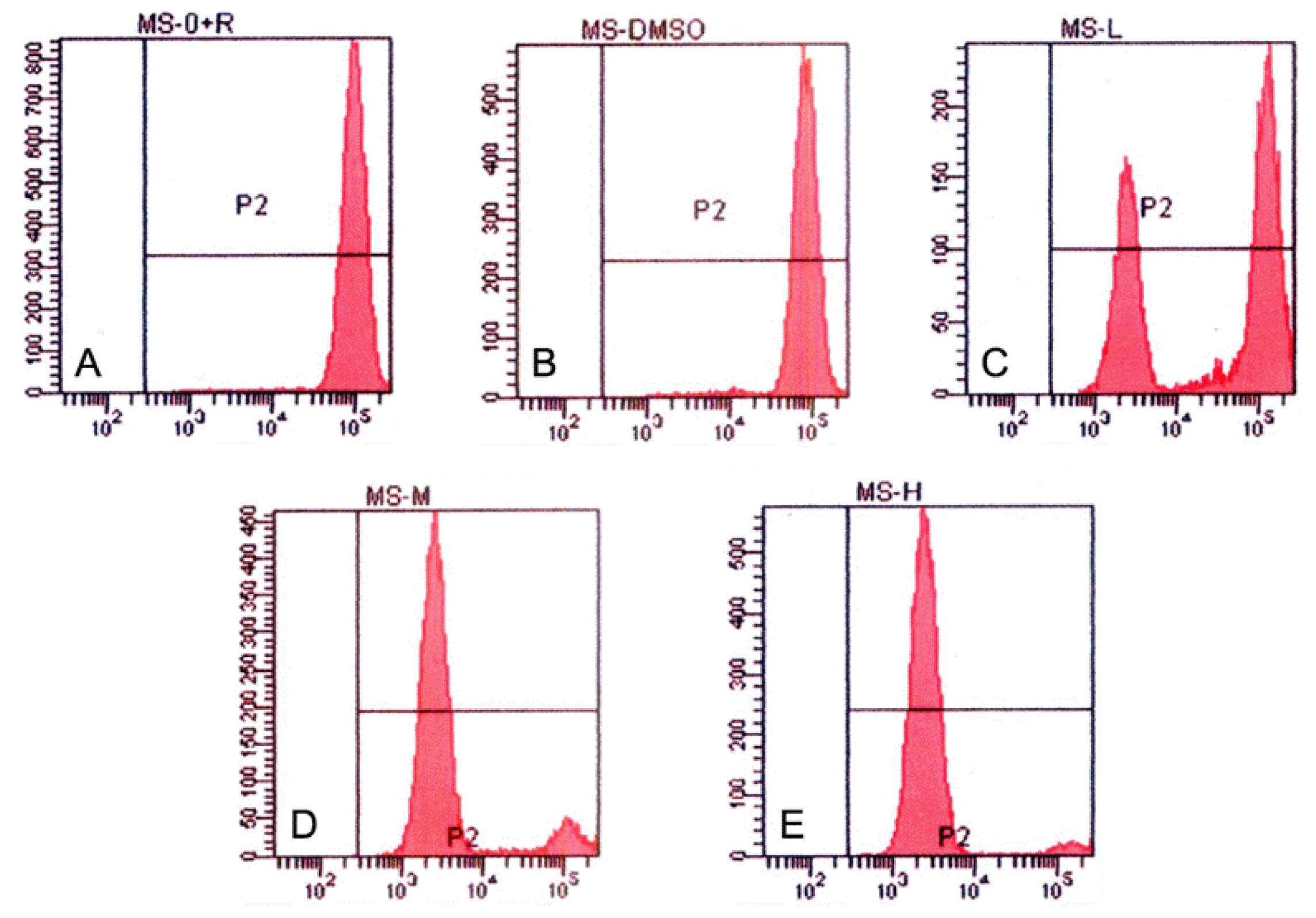
3.4. Tryptanthrin-Induced K562 Cell Apoptosis and FCM Analysis
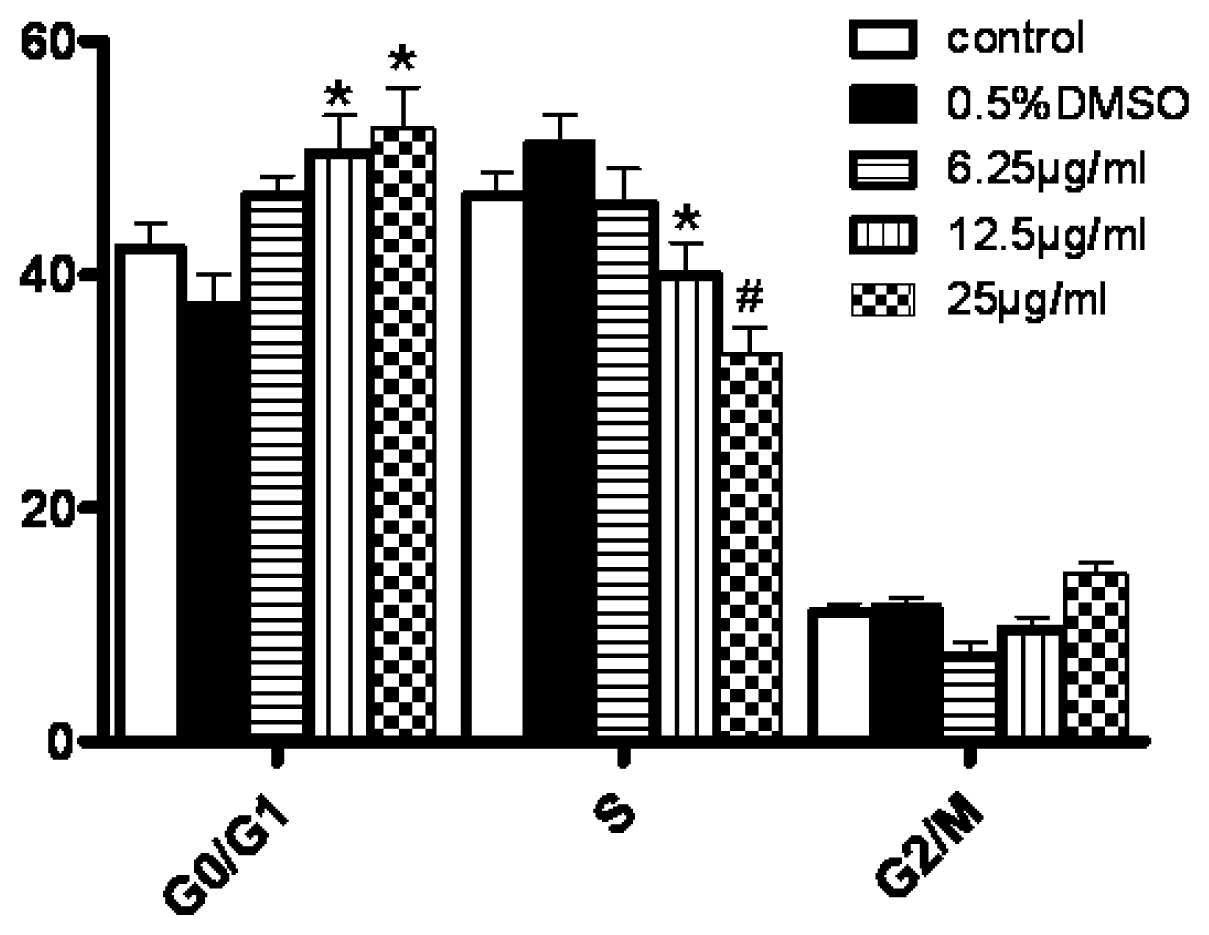
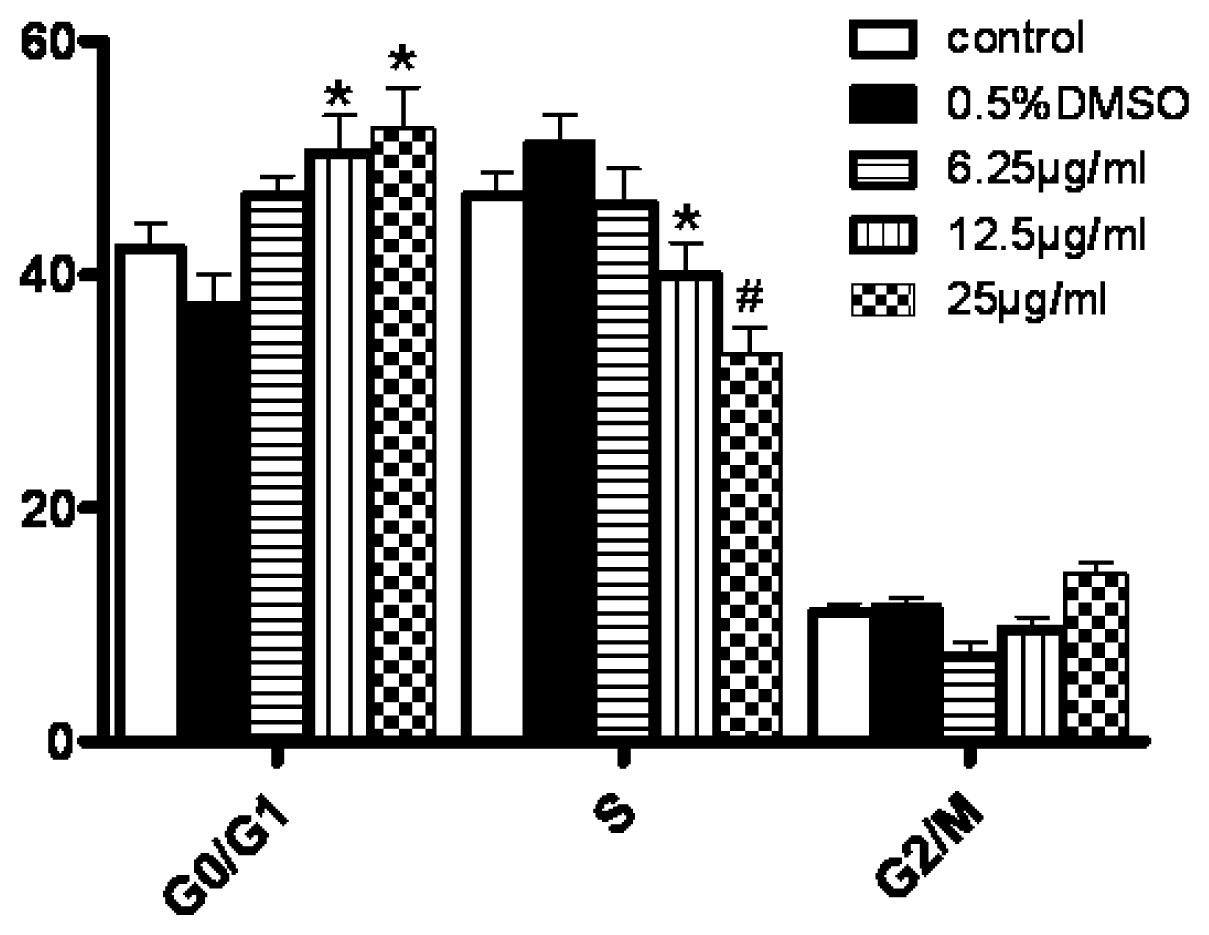
3.5. Changes in Cell Cycle Progression
3.6. Changes in Mitochondrial Membrane Potentials
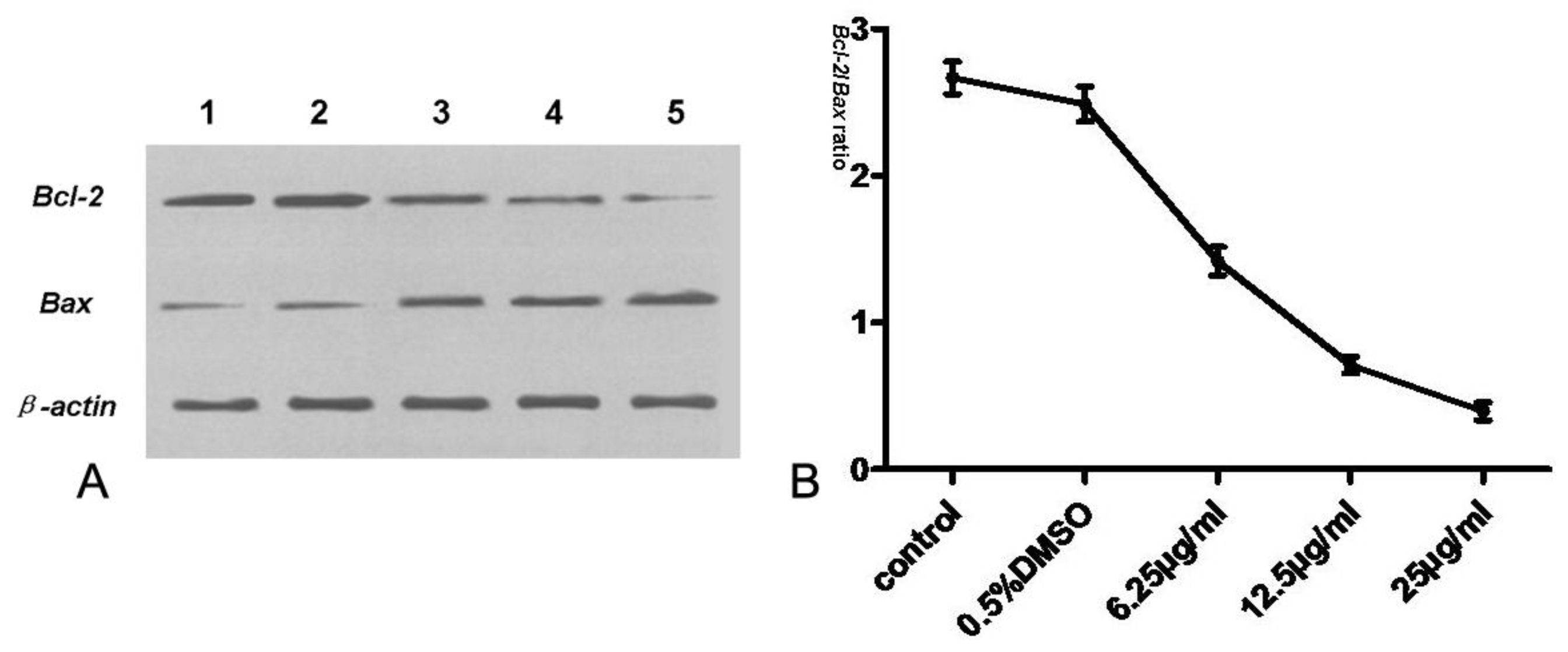
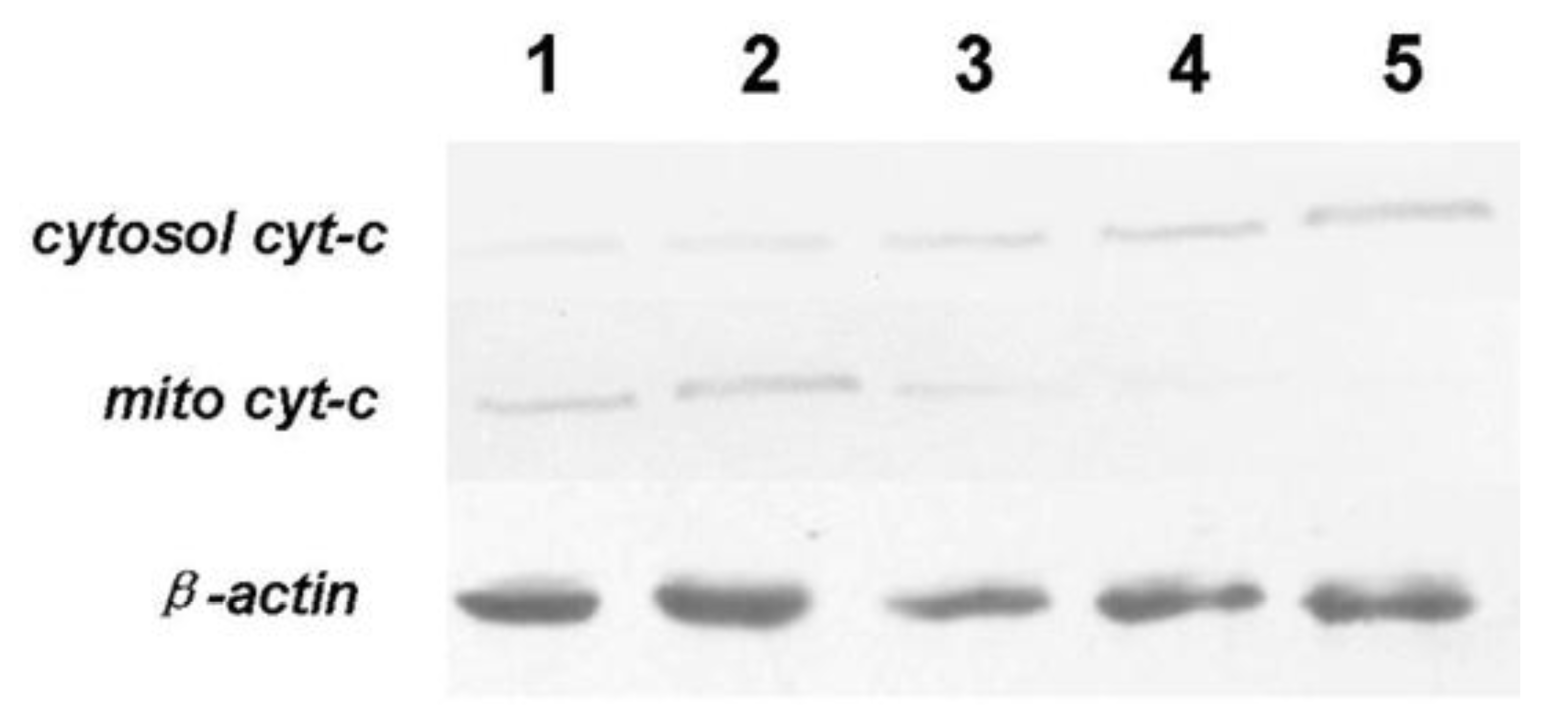
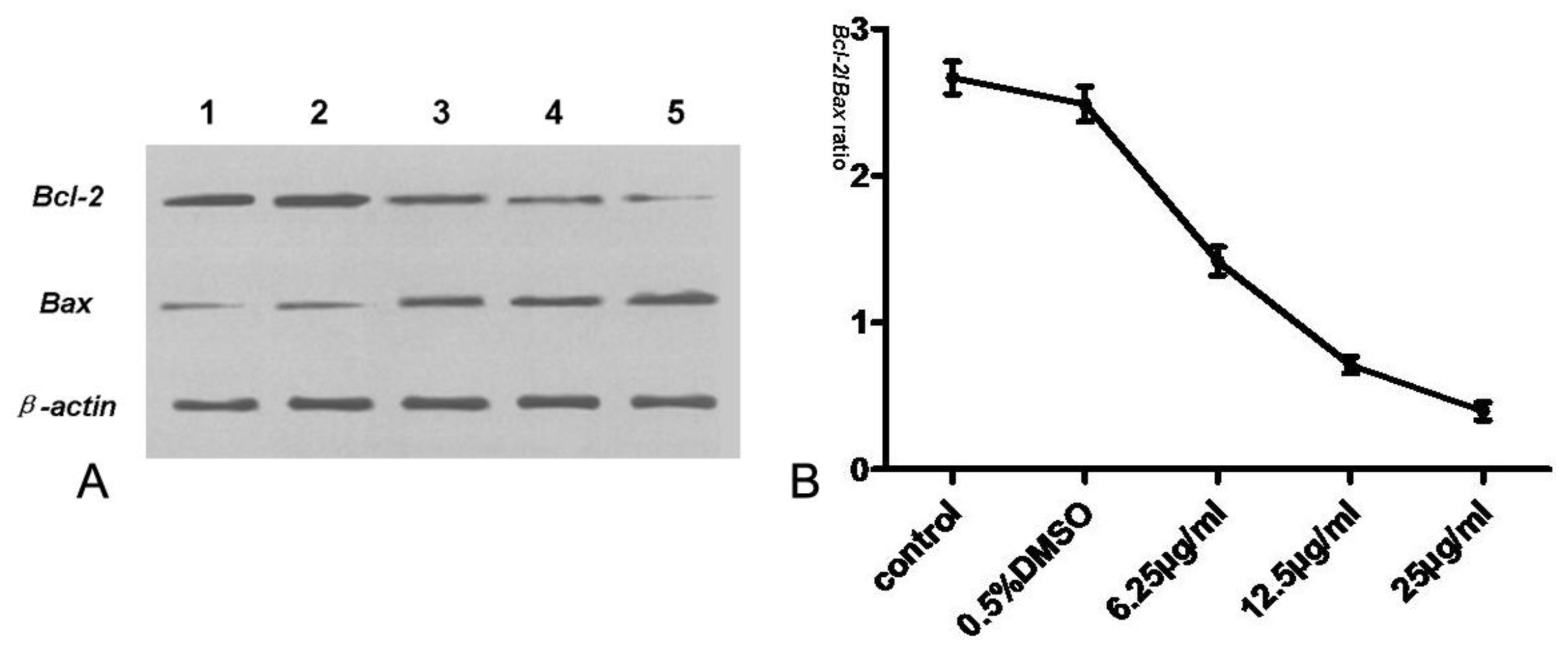
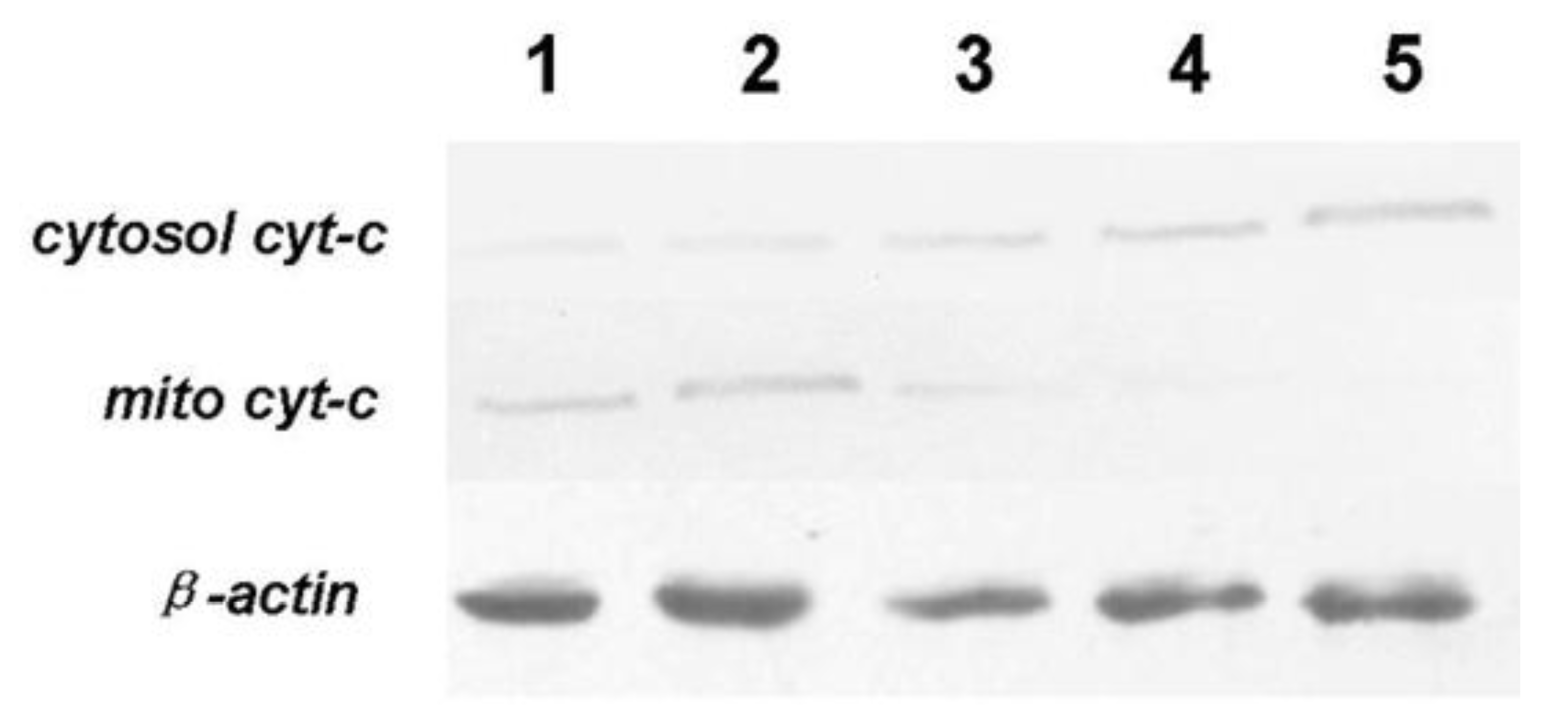
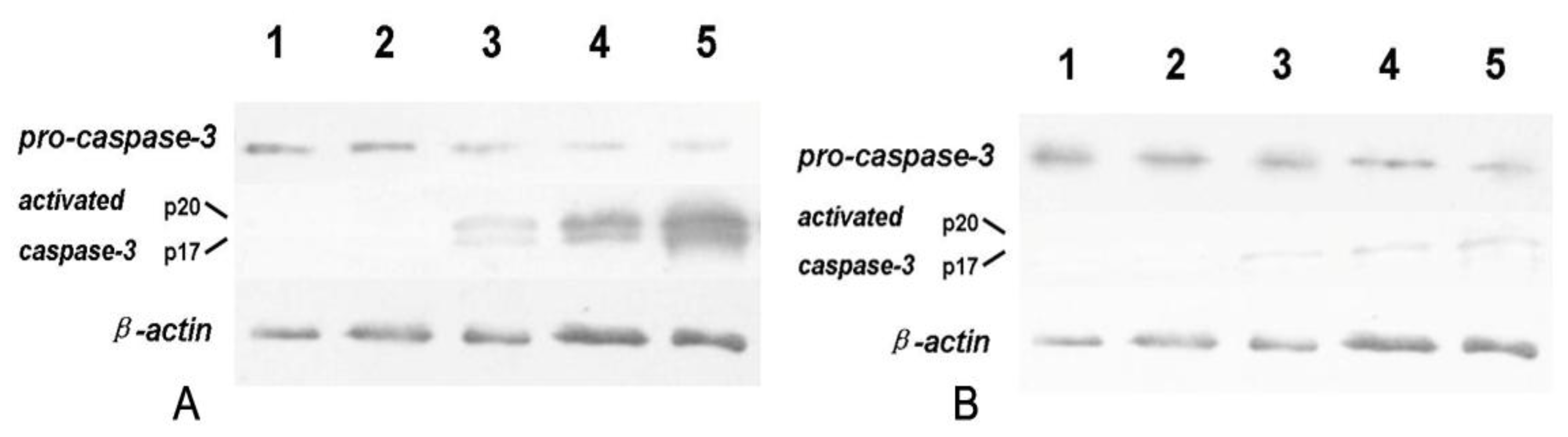
3.7. Expression of Bcl-2, Bax, Cyt-c and Caspase-3
4. Discussion
5. Conclusions
Acknowledgements
References
- Liu, JJ; Zhang, Y; Guang, WB; Yang, HZ; Lin, DJ; Xiao, RZ. Ponicidin inhibits monocytic leukemia cell growth by induction of apoptosis. Int. J. Mol. Sci 2008, 9, 2265–2277. [Google Scholar]
- Shim, MJ; Kim, HJ; Yang, SJ; Lee, IS; Choi, HI; Kim, T. Arsenic trioxide induces apoptosis in chronic myelogenous leukemia K562 cells: Possible involvement of p38 MAP kinase. J. Biochem. Mol. Biol 2002, 35, 377–383. [Google Scholar]
- Zhang, Y; Dawson, MI; Mohammad, R; Rishi, AK; Farhana, L; Feng, KC; Leid, M; Peterson, V; Zhang, XK; Edelstein, M; et al. Induction of apoptosis of human B-CLL and ALL cells by a novel retinoid and its nonretinoidal analog. Blood 2002, 100, 2917–2925. [Google Scholar]
- Chowdhury, I; Tharakan, B; Bhat, GK. Current concepts in apoptosis: The physiological suicide program revisited. Cell. Mol. Biol. Lett 2006, 11, 506–525. [Google Scholar]
- Darzynkiewicz, Z; Juan, G; Li, X; Gorczyca, W; Murakami, T; Traganos, F. Cytometry in cell necrobiology: Analysis of apoptosis and accidental cell death (necrosis). Cytometry 1997, 27, 1–20. [Google Scholar]
- Kataoka, M; Hirata, K; Kunikata, T; Ushio, S; Iwaki, K; Ohashi, K; Ikeda, M; Kurimoto, M. Antibacterial action of tryptanthrin and kaempferol, isolated from the indigo plant (Polygonum tinctorium Lour.), against Helicobacter pylori-infected Mongolian gerbils. J. Gastroenterol 2001, 36, 5–9. [Google Scholar]
- Danial, NN; Korsmeyer, SJ. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar]
- Li, P; He, QP; Ouyang, YB; Liu, CL; Hu, BR; Siesjö, BK. Early release of cytochrome C and activation of caspase-3 in hyperglycemic rats subjected to transient forebrain ischemia. Brain Res 2001, 896, 69–76. [Google Scholar]
- Yan, Q; Li, Y; Jiang, Z; Sun, Y; Zhu, L; Ding, Z. Antiproliferation and apoptosis of human tumor cell lines by a lectin (AMML) of Astragalus mongholicus. Phytomedicine 2009, 16, 586–593. [Google Scholar]
- Honda, G; Tosirisuk, V; Tabata, M. Isolation of an antidermatophytic, Tryptanthrin, from Indigo plants, Polygonum tinctorium and Isatis tinctoria. Planta Med 1980, 38, 275–276. [Google Scholar]
- Ishihara, T; Kohno, K; Ushio, S; Iwaki, K; Ikeda, M; Kurimoto, M. Tryptanthrin inhibits nitric oxide and prostaglandin E2 synthesis by murine macrophages. Eur. J. Pharmacol 2000, 407, 197–204. [Google Scholar]
- Recio, MC; Cerdá-Nicolás, M; Potterat, O; Hamburger, M; Ríos, JL. Anti-inflammatory antiallergic activity in vivo of lipophilic Isatis tinctoria extracts and tryptanthrin. Planta Med 2006, 72, 539–546. [Google Scholar]
- Yu, ST; Chen, TM; Tseng, SY; Chen, YH. Tryptanthrin inhibits MDR1 and reverses doxorubicin resistance in breast cancer cells. Biochem. Biophys. Res. Commun 2007, 358, 79–84. [Google Scholar]
- Arokium, H; Camougrand, N; Vallette, FM; Manon, S. Studies of the interaction of substituted mutants of Bax with yeast mitochondria reveal that the C-terminal hydrophobic α-helix is a second ART sequence and plays a role in the interaction with anti-apoptotic Bcl-xL. J. Biol. Chem 2004, 279, 52566–52573. [Google Scholar]
- Tolomeo, M; Grimaudo, S; Di Cristina, A; Pipitone, RM; Dusonchet, L; Meli, M; Crosta, L; Gebbia, N; Invidiata, FP; Titone, L; et al. Galangin increases the cytotoxic activity of imatinib mesylate in imatinib-sensitive and imatinib-resistant Bcr-Abl expressing leukemia cells. Cancer Lett 2008, 265, 289–297. [Google Scholar]
- Deininger, MW; Goldman, JM; Lydon, N; Melo, JV. The tyrosine kinase inhibitor CGP57148B selectively inhibits the growth of Bcr-Abl-positive cells. Blood 1997, 90, 3691–3698. [Google Scholar]
- Horita, M; Andreu, EJ; Benito, A; Arbona, C; Sanz, C; Benet, I; Prosper, F; Fernandez-Luna, JL. Blockade of the Bcr-Abl kinase activity induces apoptosis of chronic myelogenous leukemia cells by suppressing signal transducer and activator of transcription 5-dependent expression of Bcl-xL. J. Exp. Med 2000, 191, 977–984. [Google Scholar]
- Chu, S; Li, L; Singh, H; Bhatia, R. BCR-tyrosine 177 plays an essential role in Ras and Akt activation and in human hematopoietic progenitor transformation in chronic myelogenous leukaemia. Cancer Res 2007, 67, 7045–7053. [Google Scholar]
- Gangemi, RM; Santamaria, B; Bargellesi, A; Cosulich, E; Fabbi, M. Late apoptotic effects of taxanes on K562 erythroleukemia cells: Apoptosis is delayed upstream of caspase-3 activation. Int. J. Cancer 2000, 85, 527–533. [Google Scholar]
- Liu, XD; Fan, RF; Zhang, Y; Yang, HZ; Fang, ZG; Guan, WB; Lin, DJ; Xiao, RZ; Huang, RW; Huang, HQ; et al. Down-regulation of telomerase activity and activation of caspase-3 are responsible for tanshinone I-induced apoptosis in monocyte leukemia cells in vitro. Int. J. Mol. Sci 2010, 11, 2267–2280. [Google Scholar]
- Micallef, MJ; Iwaki, K; Ishihara, T; Ushio, S; Aga, M; Kunikata, T; Koya-Miyata, S; Kimito, T; Ikeda, M; Kurimoto, M. The natural plant product tryptanthrin ameliorates dextran sodium sulfate-induced colitis in mice. Int. Immunopharmacol 2002, 2, 565–578. [Google Scholar]
- Henning, D; Stefka, S; Olivier, ART; Simon, HU; Dannhardt, G; Ulbrich, H; Hamburger, M. Inhibitory activity of tryptanthrin on prostaglandin and leukotriene syntheses. Planta Med 2002, 68, 875–880. [Google Scholar]
- Takei, Y; Kunikata, T; Aga, M; Inoue, S; Ushio, S; Iwaki, K; Ikeda, M; Kurimoto, M. Tryptanthrin inhibits interferon-γ production by Peyer’s patch lymphocytes derived from mice that had been orally administered staphylococcal enterotoxin. Biol. Pharm. Bull 2003, 26, 365–367. [Google Scholar]
- Lim, ML; Lum, MG; Hansen, TM; Roucou, X; Nagley, P. On the release of cytochrome c from mitochondria during cell death signaling. J. Biomed. Sci 2002, 9, 488–506. [Google Scholar]
- Fei, XF; Wang, BX; Li, TJ; Tashiro, S; Minami, M; Xing, DJ; Ikejima, T. Evodiamine, a constituent of Evodiae Fructus, induces anti-proliferating effects in tumor cells. Cancer Sci 2003, 94, 92–98. [Google Scholar]

© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miao, S.; Shi, X.; Zhang, H.; Wang, S.; Sun, J.; Hua, W.; Miao, Q.; Zhao, Y.; Zhang, C. Proliferation-Attenuating and Apoptosis-Inducing Effects of Tryptanthrin on Human Chronic Myeloid Leukemia K562 Cell Line in Vitro. Int. J. Mol. Sci. 2011, 12, 3831-3845. https://doi.org/10.3390/ijms12063831
Miao S, Shi X, Zhang H, Wang S, Sun J, Hua W, Miao Q, Zhao Y, Zhang C. Proliferation-Attenuating and Apoptosis-Inducing Effects of Tryptanthrin on Human Chronic Myeloid Leukemia K562 Cell Line in Vitro. International Journal of Molecular Sciences. 2011; 12(6):3831-3845. https://doi.org/10.3390/ijms12063831
Chicago/Turabian StyleMiao, Shan, Xiaopeng Shi, Hai Zhang, Siwang Wang, Jiyuan Sun, Wei Hua, Qing Miao, Yong Zhao, and Caiqin Zhang. 2011. "Proliferation-Attenuating and Apoptosis-Inducing Effects of Tryptanthrin on Human Chronic Myeloid Leukemia K562 Cell Line in Vitro" International Journal of Molecular Sciences 12, no. 6: 3831-3845. https://doi.org/10.3390/ijms12063831
APA StyleMiao, S., Shi, X., Zhang, H., Wang, S., Sun, J., Hua, W., Miao, Q., Zhao, Y., & Zhang, C. (2011). Proliferation-Attenuating and Apoptosis-Inducing Effects of Tryptanthrin on Human Chronic Myeloid Leukemia K562 Cell Line in Vitro. International Journal of Molecular Sciences, 12(6), 3831-3845. https://doi.org/10.3390/ijms12063831