Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe)



Abstract
:1. Introduction
2. Results and Discussion
2.1. Plant Biomass, Photosynthesis Rate, Water Use Efficiency
2.2. Total Soluble Carbohydrate (TSC) and Starch Content
2.3. Total Phenolic and Flavonoids
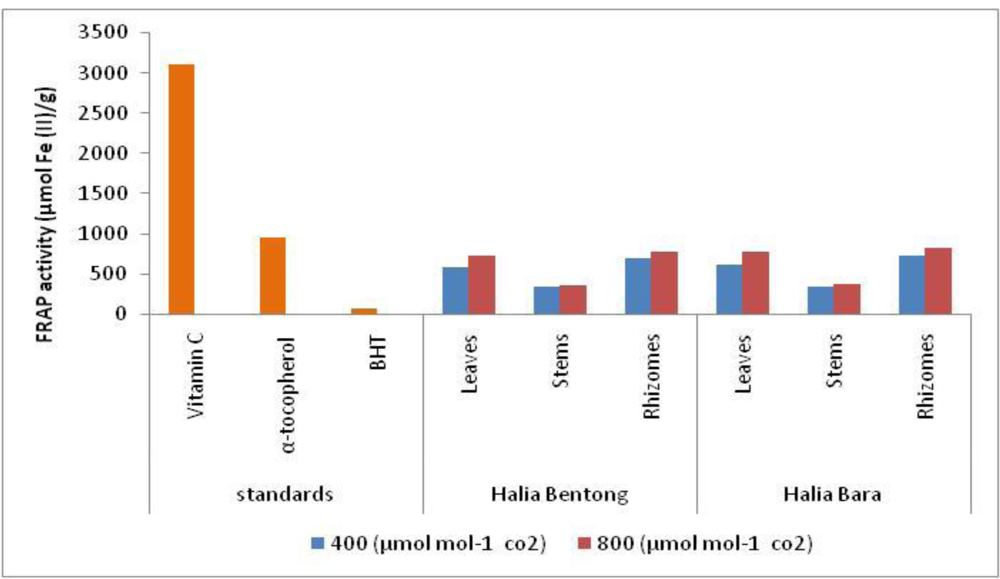
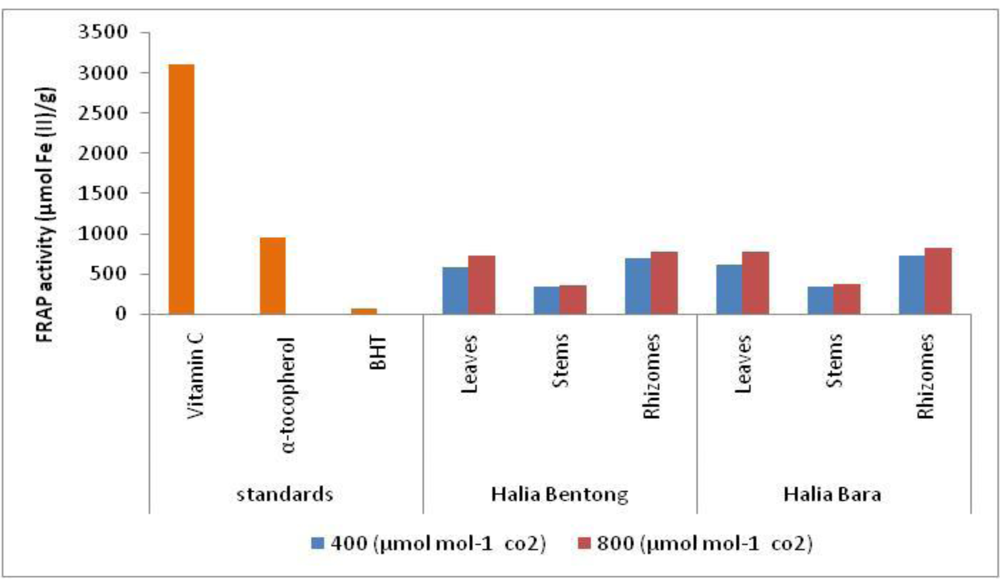
2.4. Ferric Reducing/Antioxidant Potential (FRAP)
3. Experimental
3.1. Plants Material
3.2. Extract Preparation
3.3. Determination of Plant Biomass and Photosynthesis Rate
3.4. Determination of Total Soluble Carbohydrate (TSC)
3.5. Determination of Starch Content
3.6. Determination of Total Phenolic Content
3.7. Determination of Total Flavonoids
3.8. Determination of Antioxidant Activities
Reducing Ability (FRAP Assay)
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Bazzaz, FA. The response of natural ecosystems to the rising global CO2 levels. Annu. Rev. Ecol. Syst 1990, 21, 167–196. [Google Scholar]
- Mark, SJ. Growth response of Quercuse petraea, Fraxinus excelsior and Pinus sylvestris to elevated carbon dioxide, ozone and water supply. New Phytol 2000, 146, 437–451. [Google Scholar]
- Mattson, WJ; Julkunen-Tiitto, R; Herms, DA. CO2 enrichment and carbon partitioning to phenolics: Do plant responses accord better with the protein competition or the growth-differentiation balance models? Oikos 2005, 111, 337–347. [Google Scholar]
- Wang, YSH; Bunce, AJ; Maas, LJ. Elevated carbon dioxide increases contents of antioxidant compounds in field-grown strawberries. J. Agric. Food Chem 2003, 51, 4315–4320. [Google Scholar]
- Pearson, M; Davies, WJ; Mansfield, TA. Asymmetric responses of adaxial and abaxial stomata to elevated CO2: Impacts on the control of gas-exchange by leaves. Plant Cell Environ 1995, 18, 837–843. [Google Scholar]
- Niklaus, PA; Spinnler, D; Kornerb, C. Soil moisture dynamics of calcareous grassland under elevated CO2. Oecologia 1998, 17, 201–208. [Google Scholar]
- Saxe, H; Ellsworth, DS; Heath, J. Tree and forest functioning in an enriched CO2 atmosphere. New Phytol 1998, 139, 395–436. [Google Scholar]
- Eamus, D. The interaction of rising atmospheric CO2 and temperatures with water use efficiency. Plant Cell Environ 1991, 14, 843–852. [Google Scholar]
- Bunce, JA. Carbon dioxide effects on stomatal responses to the environment and water use by crops under field conditions. Oecologia 2004, 140, 1–10. [Google Scholar]
- Liu, JX; Zhang, DQ; Zhou, GY; Faivre-Vuillin, B; Deng, Q; Wang, CL. CO2 enrichment increases nutrient leaching from model forest ecosystems in subtropical China. Biog. Dis 2008, 5, 2679–2706. [Google Scholar]
- Eamus, D. The interaction of rising atmospheric CO2 and temperatures with water use efficiency. Plant Cell Environ 1991, 14, 843–852. [Google Scholar]
- Booker, FL. Influence of carbon dioxide enrichment, ozone and nitrogen fertilization on cotton (Gossypium hirsutum L.) leaf and root composition. Plant Cell Environ 2000, 23, 573–583. [Google Scholar]
- Kaufman, PB; Cseke, LJ; Warber, S; Duke, JA; Brielmann, HL. Natural Products from Plants; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Wink, M. Introduction: Biochemistry, Role and Biotechnology of Secondary Products Biochemistry of Secondary Product Metabolism; CRC Press: Boca Raton, FL, USA, 1999; pp. 1–16. [Google Scholar]
- Malencic, D; Popovic, M; Miladinovic, J. Phenolic content and antioxidant properties of soybean (Glycine max (L.)) Merr. Seeds. Molecules 2007, 12, 576–581. [Google Scholar]
- Andlauer, W; Fürst, P. Antioxidative power of phytochemicals with special reference to cereals. Cereal Food World 1998, 43, 356–360. [Google Scholar]
- Amin, I; Tan, SH. Antioxidant activity of selected commercial seaweeds. Malays. J. Nut 2002, 8, 167–177. [Google Scholar]
- Atoui, K; Mansouri, A; Bosku, G; Kefalas, P. Tea and herbal infusions: Their antioxidant activity and phenolic profile. Food Chem 2005, 89, 27–36. [Google Scholar]
- Chan, EWC; Lim, YY; Wong, LF; Lianto, FS; Wong, SK. Antioxidant and tyrosinase inhibition properties of leaves and rhizomes of ginger species. Food Chem 2008, 109, 477–483. [Google Scholar]
- Malikov, VM; Yuledashev, MP. Phenolic compounds of plants of the Scutellaria L. genus: Distribution, structure, and properties. Chem. Nat. Compd 2002, 38, 358–406. [Google Scholar]
- Ibrahim, MH; Jaafar, HZ; Rahmat, A; Rahman, ZA. The Relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules 2011, 16, 162–174. [Google Scholar]
- Lindroth, RL; Kinney, KK; Platz, CL. Responses of deciduous trees to elevated atmospheric CO2: Productivity, phytochemistry, and insect performance. Ecology 1993, 74, 763–777. [Google Scholar]
- Tisserat, B; Vaughn, SF. Essential oils enhanced by ultra-high carbon dioxide levels from Lamiaceae species grown in vitro and in vivo. Plant Cell Rep 2001, 20, 361–368. [Google Scholar]
- Rozanida, AR; Nurul Izza, N; Mohd Helme, MH; Zanariah, H. Xanwhite TM—A Cosmeceutical Product from Species in the Family Zingiberaceae; Forest Research Institute: Selangor, Malaysia, 2005; pp. 31–36. [Google Scholar]
- Devasagayam, PA; Tilak, JC; Boloor, KK; Sane, KS; Ghaskadbi, SS; Lele, RD. Free radicals and antioxidants in human health: current status and future prospects. J. Assoc. Physic India 2004, 52, 794–804. [Google Scholar]
- Serafini, M; Bellocco, R; Wolk, A; Ekstrom, AM. Total antioxidant potential of fruit and vegetables and risk of gastric cancer. Gastroenterology 2002, 123, 985–991. [Google Scholar]
- Madsen, HL; Nielsen, BR; Bertelsen, G; Skibsted, LH. Screening of antioxidative activity of spices: Comparisons between assays based on ESR spin trapping and electrochemical measurement of oxygen consumption. Food Chem 1996, 57, 331–337. [Google Scholar]
- Reddy, AC; Lokesh, BR. Studies on spice principles as antioxidants in the inhibition of lipid peroxidation of rat liver microsomes. Mol. Cell. Biochem 1992, 111, 117–124. [Google Scholar]
- Hasna, O; Afidah, A. Antioxidant activity and phenolic content of Paederia foetida and Syzygium aqueum. Molecules 2009, 14, 970–978. [Google Scholar]
- Praven, K; Ramamoorty, A; Awang, B. Anti oxidant activity, total phenolic and flavonoid content in Morinda citrifolia fruit. J. Eng. Sci 2007, 2, 70–80. [Google Scholar]
- Lavola, A; Julkunen, TR. The effect of elevated carbon dioxide and fertilization on primary and secondary metabolites in Befula pendula (Roth). Oecologia 1994, 99, 315–321. [Google Scholar]
- Reitz, SR; Karowe, D; Diawara, MM; Tumble, JT. Effects of elevated atmospheric carbon dioxide on the growth and linear furanocoumarin content of celery. J. Agric. Food Chem 1997, 45, 3642–3646. [Google Scholar]
- Srivastava, K; Misra, A; Sharma, S. Effect of Zn deficiency on net photosynthetic rate, 14C partitioning, and oil accumulation in leaves of peppermint. Photosynthetica 1997, 33, 71–79. [Google Scholar]
- Buddendorf-Joosten, JMC; Woltering, EJ. Components of the gaseous environment and their effects on plant growth and development in vitro. J. Plant Growth Regul 1994, 15, 1–16. [Google Scholar]
- Gielen, B; Ceulemans, R. The likely impact of rising atmospheric CO2 on natural and managed Populus: A literature review. Environ. Pollut 2001, 115, 335–358. [Google Scholar]
- Norby, RJ; Wullschleger, SD; Gunderson, CA; Johnson, DW; Ceulemans, R. Tree responses to rising CO2 in field experiments: Implications for the future forest. Plant Cell Environ 1999, 22, 683–714. [Google Scholar]
- Reekie, EG; Bazzaz, FA. Competition and patterns of resource use among seedlings of tropical trees grown at ambient and elevated CO2. Oecologia 1989, 79, 212–222. [Google Scholar]
- Yusuke, O; Tadaki, H; Kouki, H. Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three natural CO2 springs in Japan. Ecol. Res 2007, 22, 475–484. [Google Scholar]
- Morison, JIL. Intercellular carbon dioxide concentration and stomatal responses to carbon dioxide. In Stomatal Function; Stanford University Press: Stanford, CA, USA, 1987; pp. 229–251. [Google Scholar]
- Jackson, RB; Sala, OE; Field, CB; Mooney, HA. Carbon dioxide alters water use, carbon gain, and yield of dominant species in a natural grassland. Oecologia 1994, 98, 257–262. [Google Scholar]
- Jeffrey, QC; Whendee, LS. Some aspect of ecophysiological and biogeochemical responses of tropical forest to atmospheric changes. Philos. Trans. R. Soc. Lond. B 2004, 359, 549–555. [Google Scholar]
- Moore, BD; Cheng, SH; Rice, J; Seemann, JR. Sucrose cycling, Rubisco expression, and prediction of photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ 1998, 21, 905–916. [Google Scholar]
- Wolfe, DW; Gifford, RM; Hilbert, D; Luo, Y. Integration of photosynthetic acclimation to CO2 at the wholeplant level. Global Change Biol 1998, 4, 879–893. [Google Scholar]
- Landolt, W; Pfenninger, I. The effect of elevated CO2 and soil type on non-structural carbohydrates in beech leaves and norway spruce needles growing in model ecosystems. Acta Oecol 1997, 18, 351–359. [Google Scholar]
- Herrmann, KM; Weaver, LM. The shikimate pathway. Annu. Rev. Plant Phys. Plant Mol. Biol 1999, 50, 473–503. [Google Scholar]
- Penuelas, J; Estiarte, M. Can elevated CO2 affect secondary metabolism and ecosystem function? Trees 1998, 13, 20–24. [Google Scholar]
- Ramawat, KG. Herbal Drugs: Ethnomedicine to Modern Medicine; Springer: New York, NY, USA, 2009; p. 414. [Google Scholar]
- Stutte, GW; Eraso, I. Carbon dioxide enrichment enhances growth and flavonoid content of two Scutellaria species. J. Am. Soc. Hortic. Sci 2008, 133, 631–638. [Google Scholar]
- Baumann, J; Wurn, G; Bruchlausen, FV. Prostaglandin synthetase inhibiting O2 radical scavenging properties of some flavonoids and related phenolic compounds. Deutsche Pharmakologische Gesellschaft Abstracts of the 20th spring meeting. Naunyn-Schmiedeberg’s Arch. Pharmacol 1979, 307, 1–77. [Google Scholar]
- Miliauskas, G; Venskutonis, PR; van Beek, TA. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem 2004, 85, 231–237. [Google Scholar]
- Mohd Zin, Z; Abdul-Hamid, A; Osman, A. Antioxidative activity of extracts from mengkudu (Morinda citrifolia L.) root, fruit and leaf. Food Chem 2002, 78, 227–231. [Google Scholar]
- Luximon-Ramma, A; Bahorun, T; Soobrattee, AM; Aruoma, OI. Antioxidant activities of phenolic, proanthocyanidin and flavonoid components in extracts of Acacia fistula. J. Agric. Food Chem 2005, 50, 5042–5047. [Google Scholar]
- Oktay, M; Gulcin, I; Kufrevioglu, OI. Determination of in vitro antioxidant activity of fennel (Foeniculum vulgare) seed extracts. Lebensm. Wiss. Technol 2003, 36, 263–271. [Google Scholar]
- Sivaci, A. Seasonal changes of total carbohydrate contents in three varieties of apple (Malus sylvestris Miller) stem cuttings. Sci. Hortic 2006, 109, 234–237. [Google Scholar]
- McCready, MR; Guggolz, J; Silviera, V; Owens, SH. Determination of starch and amylose in vegetables. Anal. Chem 1950, 22, 1156–1158. [Google Scholar]
- Kim, D; Jeond, S; Lee, CY. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem 2003, 81, 321–326. [Google Scholar]
- Bushra, S; Farooq, A; Muhammad, A. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar]
- Benzie, IFF; Strain, JJ. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: the FRAP assay. Anal. Biochem 1996, 239, 70–76. [Google Scholar]
- Penuelas, J; Estiarte, M; Liusia, J. Carbon based secondary compounds at elevated CO2. Photosynthetica 1997, 33, 313–316. [Google Scholar]

{kind=link}
{kind=link}
| Parameters | Halia Bentong | Halia Bara | ||
|---|---|---|---|---|
| 400 | 800 | 400 | 800 | |
| Leaves biomass | 22.83 ±0.91c | 35.3 ±0.46b | 20.79 ±0.37d | 38.5 ± 1.18a |
| Stems biomass | 19.1 ±1.23a | 23.8 ±0.47a | 20 ±1.26a | 24.4 ± 0.046a |
| Rhizomes biomass | 14.5 ±0.29b | 24.1 ±1.005a | 6.91 ±0.049c | 14.05 ± 0.61b |
| Total Biomass | 56.5 ±1.85c | 83.4 ±1.93a | 47.7 ±0.84d | 77.05 ± 1.76b |
| Photosynthesis | 5.58 ±0.24d | 9.22 ±0.35b | 6.86 ±0.028c | 10.05 ±0.14a |
| Stomatal conductance | 0.182 ±0.005a | 0.126 ±0.03ab | 0.106 ±0.015ab | 0.08 ± 0.009b |
| Water use efficiency | 1.52 ±0.056b | 1.85 ±0.035a | 0.99 ±0.042c | 1.48 ± 0.007b |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Photosynthesis | 1 | ||||||||
| 2 | Stomatal conductance | −0.56n.s | 1 | |||||||
| 3 | WUE | 0.87** | −0.81** | 1 | ||||||
| 4 | Biomass | 0.85** | −0.90** | 0.86** | 1 | |||||
| 5 | TSC | 0.96** | −0.72* | 0.60n.s | 0.93** | 1 | ||||
| 6 | Starch | 0.92** | −0.74* | 0.71* | 0.92** | 0.94** | 1 | |||
| 7 | TP | 0.83** | −0.24n.s | 0.007n.s | 0.49n.s | 0.71* | 0.72* | 1 | ||
| 8 | TF | 0.72* | −0.18n.s | 0.07n.s | 0.5n.s | 0.70* | 0.63n.s | 0.9** | 1 | |
| 9 | FRAP | 0.71* | −0.43n.s | 0.22n.s | 0.51n.s | 0.72* | 0.49n.s | 0.76* | 0.71* | 1 |
| Varieties | Parts | CO2 Concentration (μmol mol−1) | |
|---|---|---|---|
| 400 | 800 | ||
| Halia Bentong | Leaves | 14.05 ± 0.953e | 38.22 ± 1.98a |
| Stems | 11.8 ± 0.455f | 17.26 ± 0.385d | |
| Rhizomes | 10.9 ± 0.481f | 27.63 ± 1.69c | |
| Halia Bara | Leaves | 14.46 ± 0.98e | 38.43 ± 0.935a |
| Stems | 11.59 ± 1.8f | 18.83 ± 0.895d | |
| Rhizomes | 11.46 ± 0.63f | 30.16 ± 2.004b | |
| Varieties | Parts | CO2 Concentration (μmol mol−1) | |
|---|---|---|---|
| 400 | 800 | ||
| Halia Bentong | Leaves | 311.67 ± 14.4c | 385.4 ± 10.9b |
| Rhizome | 311.9 ± 28.4c | 583.5 ± 24.9a | |
| Halia Bara | Leaves | 317.2 ± 8.2c | 402.7 ± 15.6b |
| Rhizome | 315.9 ± 15.2c | 553.3 ± 24.6a | |
| Varieties | Plant Parts | TF (mg Quercetin/g dry weight) | TP (mg Gallic acid/g dry weight) | ||
|---|---|---|---|---|---|
| CO2 Concentration (μmol mol−1) | CO2 Concentration (μmol mol−1) | ||||
| 400 | 800 | 400 | 800 | ||
| Halia Bentong | Leaves | 5.44 ±0.45de | 6.04 ±0.79d | 31.22 ±2.41d | 39.68 ±5.61c |
| Stems | 1.61 ±0.22g | 1.96 ±0.17g | 6.14 ±0.8f | 7.6 ± 0.66ef | |
| Rhizomes | 4.03 ±0.081f | 7.35 ±1.99c | 11.33 ±0.27e | 28.76 ±7.74d | |
| Halia Bara | Leaves | 8.66 ±0.42bc | 9.23 ±0.36ab | 43.22 ±2.15b | 60.69 ±2.6a |
| Stems | 1.74 ±0.37g | 2.04 ±0.31g | 7.1 ±1.04ef | 7.89 ± 1.17ef | |
| Rhizomes | 4.48 ±0.08ef | 9.78 ±0.77a | 13.5 ±0.26e | 38.16 ± 1.55c | |
| Varieties | Extraction source | CO2 Concentration (μmol mol−1) | |
|---|---|---|---|
| 400 | 800 | ||
| Halia Bentong | Leaves | 577.21 ± 14.21e | 722.31 ± 11.45d |
| Stems | 341.2 ± 40.76f | 356.31 ± 41.6f | |
| Rhizomes | 671.3 ± 21.2d | 774.62 ± 26.35bc | |
| Halia Bara | Leaves | 620.1 ± 18.7e | 783.09 ± 21.95b |
| Stems | 350.2 ± 27.6f | 367.4 ± 20.43f | |
| Rhizomes | 740.6 ± 40.15cd | 831.16 ± 39.08a | |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ghasemzadeh, A.; Jaafar, H.Z.E. Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe). Int. J. Mol. Sci. 2011, 12, 1101-1114. https://doi.org/10.3390/ijms12021101
Ghasemzadeh A, Jaafar HZE. Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe). International Journal of Molecular Sciences. 2011; 12(2):1101-1114. https://doi.org/10.3390/ijms12021101
Chicago/Turabian StyleGhasemzadeh, Ali, and Hawa Z. E. Jaafar. 2011. "Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe)" International Journal of Molecular Sciences 12, no. 2: 1101-1114. https://doi.org/10.3390/ijms12021101
APA StyleGhasemzadeh, A., & Jaafar, H. Z. E. (2011). Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe). International Journal of Molecular Sciences, 12(2), 1101-1114. https://doi.org/10.3390/ijms12021101