Role of Dehydrodiferulates in Maize Resistance to Pests and Diseases

{kind=link}
{kind=link}
Abstract
:1. Introduction
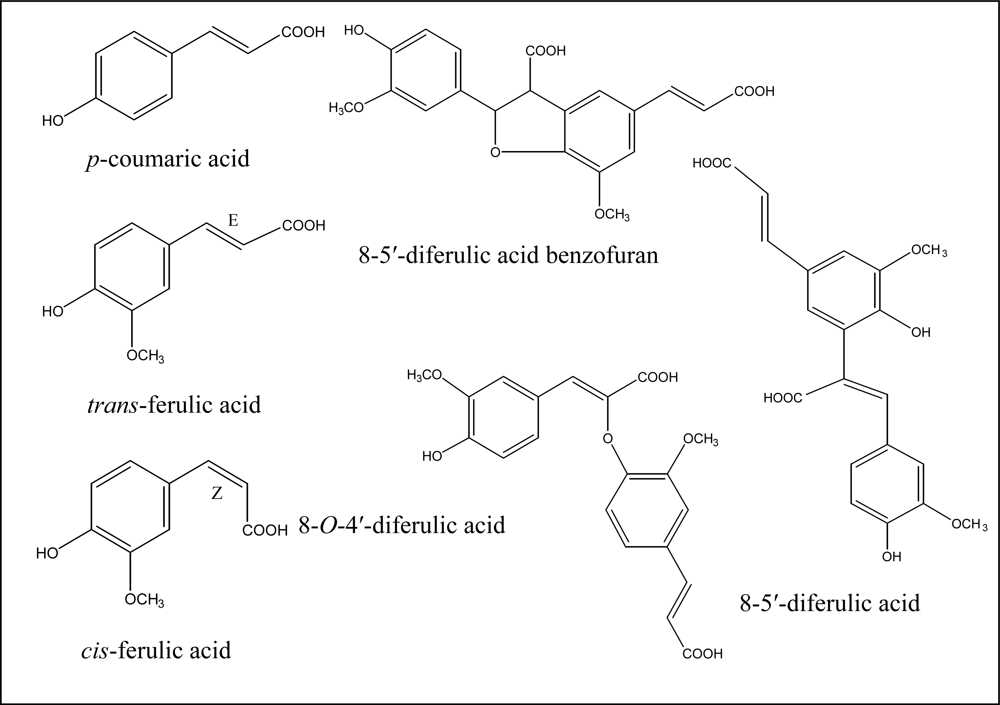
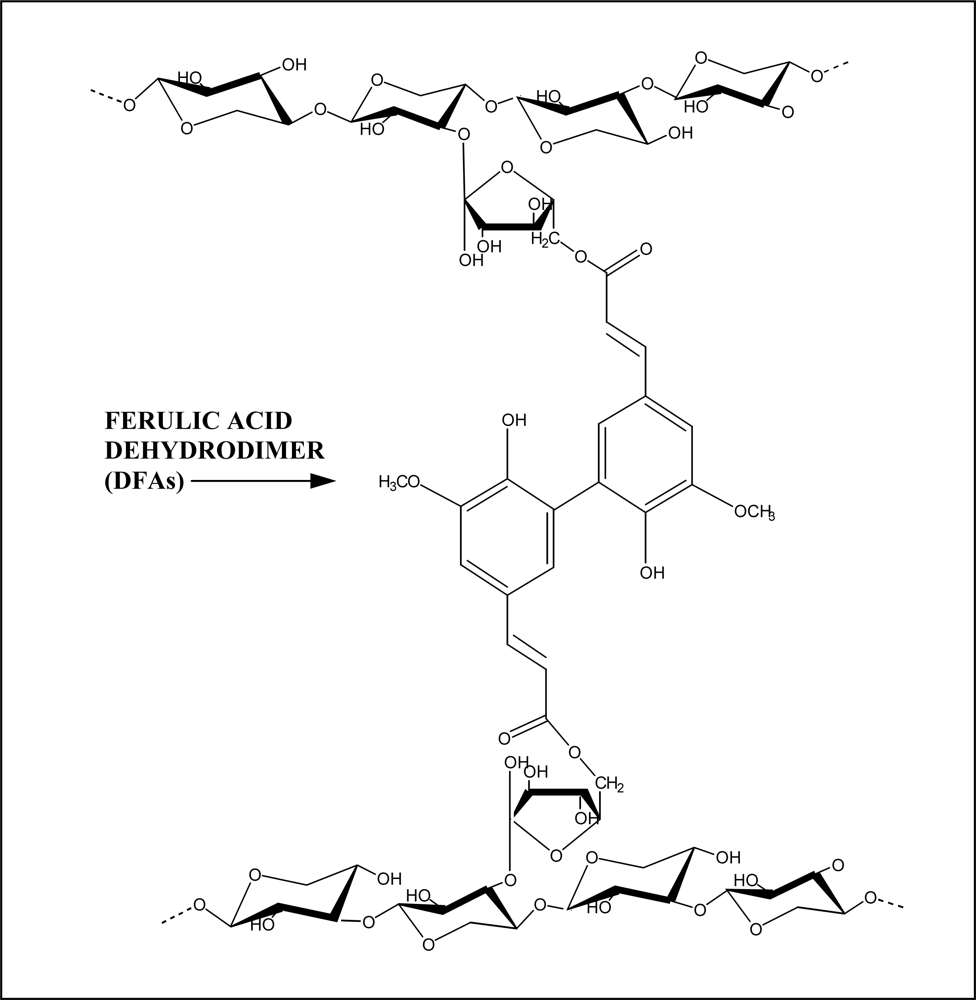
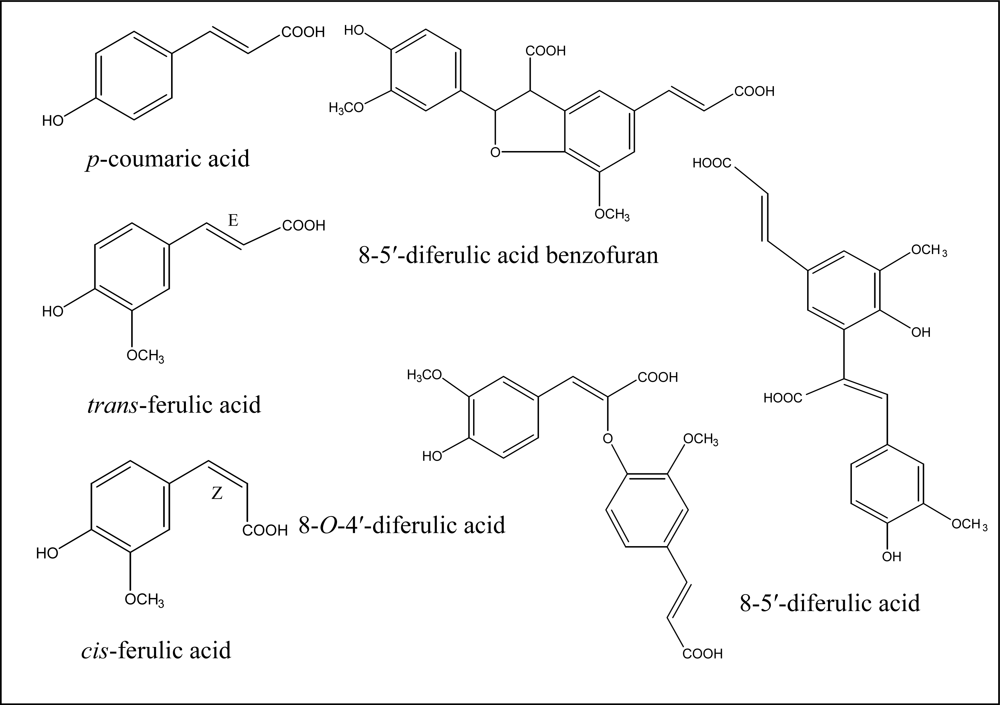
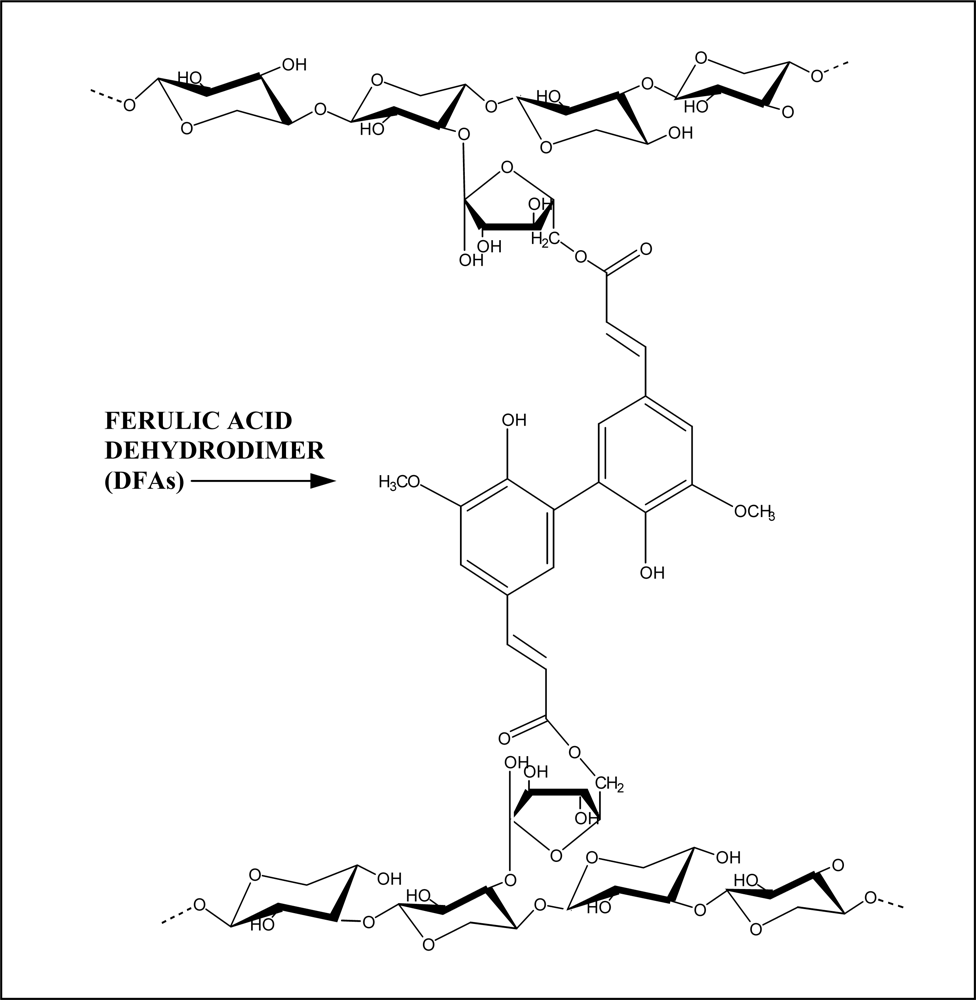
2. Cell Wall Cross-Linking by Dehydrodiferulates
3. Role of Dehydrodiferulates Compounds in Maize Resistance to Pests
3.1. Corn Borer Resistance
3.2. Maize Weevil Resistance
4. Role of Dehydrodiferulates Compounds in Maize Resistance to Fungal Diseases
4.1. Gibberella Ear Rot Resistance
4.2. Gibberella Stalk Rot Resistance
5. Conclusions
Abbreviations:
| DFA | dehydrodiferulic acid |
| ECB | european corn borer |
| MCB | mediterranean corn borer |
| MW | maize weevil |
| DON | deoxynivalenol |
Acknowledgments
References and Notes
- Carpita, NC; Defernez, M; Findlay, K; Wells, B; Shoue, DA; Catchpole, G; Wilson, RH; McCann, MC. Cell wall architecture of the elongating maize coleoptile. Plant Physiol 2001, 127, 551–565. [Google Scholar]
- Ishii, T. Structure and functions of feruloylated polysaccharides. Plant Sci 1997, 127, 111–127. [Google Scholar]
- Hatfield, RD; Wilson, JR; Mertens, DR. Composition of cell walls isolated from cell types of grain sorghum stems. J. Sci. Food Agric 1999, 79, 891–899. [Google Scholar]
- Saulnier, L; Thibault, JF. Ferulic acid and diferulic acids as components of sugar-beet pectins and maize bran heteroxylans. J. Sci. Food Agric 1999, 79, 396–402. [Google Scholar]
- Ralph, J; Hatfield, RD; Quideau, S; Helm, RF; Grabber, JH; Jung, HJG. Pathway of p-coumaric acid incorporation into maize lignin as revealed by NMR. J. Am. Chem. Soc 1994, 116, 9448–9456. [Google Scholar]
- Grabber, JH; Quideau, S; Ralph, J. p-Coumaroylated syringyl units in maize lignin; implications for β-ether cleavage by thioacidolysis. Phytochemistry 1996, 43, 1189–1194. [Google Scholar]
- Lu, F; Ralph, J. Detection and determination of p-coumaroylated units in lignins. J. Agric. Food Chem 1999, 47, 1988–1992. [Google Scholar]
- Morrison, TA; Jung, HJG; Buxton, DR; Hatfield, RD. Cell-wall composition of maize internodes of varying maturity. Crop Sci 1998, 38, 455–460. [Google Scholar]
- Vailhe, MAB; Provan, GJ; Scobbie, L; Chesson, A; Maillot, MP; Cornu, A; Besle, JM. Effect of phenolic structures on the degradability of cell walls isolated from newly extended apical internode of tall fescue (Festuca arundinacea Schreb.). J. Agric. Food Chem 2000, 48, 618–623. [Google Scholar]
- Migne, C; Prensier, G; Utille, JP; Angibeaud, P; Cornu, A; Grenet, E. Immunocytochemical localisation of p-coumaric acid and feruloyl-arabinose in the cell walls of maize stem. J. Sci. Food Agric 1998, 78, 373–381. [Google Scholar]
- MacAdam, JW; Grabber, JH. Relationship of growth cessation with the formation of diferulate cross-links and p-coumaroylated lignins in tall fescue. Planta 2002, 215, 785–793. [Google Scholar]
- Jung, HJG. Maize stem tissues: Ferulate deposition in developing internode cell walls. Phytochemistry 2003, 63, 543–549. [Google Scholar]
- Grabber, JH; Ralph, J; Lapierre, C; Barrière, Y. Genetic and molecular basis of grass cell wall degradability. I. Lignin-cell wall matrix interactions. Comptes Rendus Biol 2004, 327, 455–465. [Google Scholar]
- Biggs, KJ; Fry, SC. Phenolic cross-linking in the plant cell wall. In Physiology of Expansion During Plant Growth: Proceedings of the Second Annual Pennsylvania State Symposium in Plant Physiology; Cosgrove, DJ, Knieval, DP, Eds.; American Society of Plant Physiologists: Rockville, PA, USA, 1987; pp. 46–57. [Google Scholar]
- Brett, CT; Waldron, KW. Cell walls in diet and health. In Physiology and Biochemistry of Plant Cell Walls; Black, M, Charlwood, BV, Eds.; Chapman and Hall: London, UK, 1996; pp. 222–238. [Google Scholar]
- Bunzel, M. Chemistry and occurrence of hydroxycinnamate oligomers. Phytochem Rev 2009. doi 10.1007/s11101-009-9139-3.. [Google Scholar]
- Allerdings, E; Ralph, J; Schatz, PF; Gniechwitz, D; Steinhart, H; Bunzel, M. Isolation and structural identification of diarabinosyl 8-O-4-dehydrodiferulate from maize bran insoluble fibre. Phytochemistry 2005, 66, 113–124. [Google Scholar]
- Ralph, J; Grabber, JH; Hatfield, RD. Lignin-ferulate crosslinks in grasses: Active incorporation of ferulate polysaccharide esters into ryegrass lignins. Carbohydr. Res 1995, 275, 167–178. [Google Scholar]
- Grabber, JH; Ralph, J; Hatfield, RD. Cross-linking of maize walls by ferulate dimerization and incorporation into lignin. J. Agric. Food Chem 2000, 48, 6106–6113. [Google Scholar]
- Piber, M; Koehler, P. Identification of dehydro-ferulic acidtyrosine in rye and wheat: Evidence for a covalent cross-link between arabinoxylans and proteins. J. Agric. Food Chem 2005, 53, 5276–5284. [Google Scholar]
- Geissmann, T; Neukom, H. Vernetzung von Phenolcarbonsäureestern von Polysacchariden durch oxydative phenolische Kupplung. Helv. Chim. Acta 1971, 54, 1108–1112. [Google Scholar]
- Markwalder, HU; Neukom, H. Diferulic acid as a possible cross-link in hemicelluloses from wheat germ. Phytochemistry 1976, 15, 836–837. [Google Scholar]
- Nordkvist, E; Salomonsson, AC; Aman, P. Distribution of insoluble bound phenolic acids in barley grain. J. Sci. Food Agric 1984, 35, 657–661. [Google Scholar]
- Kamisaka, S; Takeda, S; Takahashi, K; Shibata, K. Diferulic and ferulic acids in the cell wall of oat coleoptiles—their relationships to mechanical properties of the cell wall. Physiol. Plant 1990, 78, 1–7. [Google Scholar]
- Ralph, J; Quideau, S; Grabber, JH; Hatfield, RD. Identification and synthesis of new ferulic acid dehydrodimers present in grass cell walls. J. Chem. Soc. Perkin Trans 1994, 1, 3485–3498. [Google Scholar]
- Parr, AJ; Waldron, KW; Ng, A; Parker, ML. The wall-bound phenolics of Chinese water chestnut (Eleocharis dulcis). J. Sci. Food Agric 1996, 71, 501–507. [Google Scholar]
- Parr, AJ; Ng, A; Waldron, KW. Ester-linked phenolic components of carrot cell walls. J. Agric. Food Chem 1997, 45, 2468–2471. [Google Scholar]
- Saulnier, L; Thibault, JF. Ferulic acid and diferulic acids as components of sugar-beet pectins and maize bran heteroxylans. J. Sci. Food Agric 1999, 79, 396–402. [Google Scholar]
- Bunzel, M; Ralph, J; Marita, J; Steinhart, H. Identification of 4-O-5-coupled diferulic acid from insoluble cereal fiber. J. Agric. Food Chem 2000, 48, 3166–3169. [Google Scholar]
- Bunzel, M; Ralph, J; Marita, JM; Hatfield, RD; Steinhart, H. Diferulates as structural components in soluble and insoluble cereal dietary fibre. J. Sci. Food Agric 2001, 81, 653–660. [Google Scholar]
- Bunzel, M; Funk, C; Steinhart, H. Semipreparative isolation of dehydrodiferulic and dehydrotriferulic acids as standard substances from maize bran. J. Sep. Sci 2004, 27, 1080–1086. [Google Scholar]
- Bunzel, M; Ralph, J; Funk, C; Steinhart, H. Structural elucidation of new ferulic acid-containing phenolic dimers and trimers isolated from maize bran. Tetrahedron Lett 2005, 46, 5845–5850. [Google Scholar]
- Bunzel, M; Ralph, J; Brüning, P; Steinhart, H. Structural identification of dehydrotriferulic and dehydrotetraferulic acids from insoluble maize bran fiber. J. Agric. Food Chem 2006, 54, 6409–6418. [Google Scholar]
- Rouau, X; Cheynier, V; Surget, A; Gloux, D; Barron, C; Meudec, E; Louis-Montero, J; Criton, M. A dehydrotrimer of ferulic acid from maize bran. Phytochemistry 2003, 63, 899–903. [Google Scholar]
- Funk, C; Ralph, J; Steinhart, H; Bunzel, M. Isolation and structural characterisation of 8–O–4/8–O–4- and 8–8/8–O–4-coupled dehydrotriferulic acids from maize bran. Phytochemistry 2005, 66, 363–371. [Google Scholar]
- Ralph, J; Bunzel, M; Marita, JM; Hatfield, RD; Lu, F; Kim, H; Schatz, PF; Grabber, JH; Steinhart, H. Peroxidase-dependent cross-linking reactions of p-hydroxycinnamates in plant cell walls. Phytochem. Rev 2004, 3, 79–96. [Google Scholar]
- Zarra, I; Sanchez, M; Queijeiro, E; Pena, MJ; Revilla, G. The cell wall stiffening mechanism in Pinus pinaster Aiton: Regulation by apoplastic levels of ascorbate and hydrogen peroxide. J. Sci. Food Agric 1999, 79, 416–420. [Google Scholar]
- Waldron, KW; Smith, AC; Parr, AJ; Ng, A; Parker, ML. New approaches to understanding and controlling cell separation in relation to fruit and vegetable texture. Trends Food Sci. Technol 1997, 8, 213–221. [Google Scholar]
- Ng, A; Harvey, AJ; Parker, ML; Smith, AC; Waldron, KW. Effect of oxidative coupling on the thermal stability of texture and cell wall chemistry of beet boot (Beta vulgaris). J. Agric. Food Chem 1998, 46, 3365–3370. [Google Scholar]
- Parker, CC; Parker, ML; Smith, AC; Waldron, KW. Thermal stability of texture in Chinese water chestnut may be dependent on 8, 80-diferulic acid (aryltetralyn form). J. Agric. Food Chem 2003, 51, 2034–2039. [Google Scholar]
- Waldron, KW; Parker, ML; Smith, AC. Plant cell walls and food quality. Comp. Rev. Food Sci. Food Safety 2003, 2, 128–146. [Google Scholar]
- Robertson, JA; Faulds, CB; Smith, AC; Waldron, KW. Peroxidase-mediated oxidative cross-linking and its potential to modify mechanical properties in water-soluble polysaccharide extracts and cereal grain residues. J. Agric. Food Chem 2008, 56, 1720–1726. [Google Scholar]
- Oosterveld, A; Beldman, G; Voragen, AGJ. Oxidative cross-linking of pectic polysaccharides from sugar beet pulp. Carbohydr. Res 2000, 328, 199–207. [Google Scholar]
- Grabber, JH. How do lignin composition, structure, and cross-linking affect degradability? A review of cell wall model studies. Crop Sci 2005, 45, 820–831. [Google Scholar]
- Jung, HJG; Casler, MD. Maize stem tissues: Impact of development on cell wall degradability. Crop Sci 2006, 46, 1801–1809. [Google Scholar]
- Gianessi, L; Sankula, S; Reigner, N. Plant Biotechnology: Potential impact for improving pest management in european agriculture, maize case study. The National Center for Food and Agricultural Policy. 2003. Full report available: http://www.ncfap.org/documents/ExecutiveSummaryJune.pdf (accessed November 12, 2009).
- Bergvinson, DJ; Arnason, JT; Hamilton, RI; Mihm, JA; Jewell, DC. Determining leaf toughness and its role in maize resistance to the European corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol 1994, 87, 1743–1748. [Google Scholar]
- Bergvinson, DJ; Hamilton, RI; Arnason, JT. Leaf profile of maize resistance factors to European corn borer, Ostrinia nubilalis. J. Chem. Ecol 1995, 21, 343–354. [Google Scholar]
- Bergvinson, DJ; Arnason, JT; Hamilton, RI. Phytochemical changes during recurrent selection for resistance to the European corn borer. Crop Sci 1997, 37, 1567–1572. [Google Scholar]
- Ramputh, AI. Soluble and cell wall Bound phenolic-mediated insect resistance in corn and sorghum; . Ph.D. Dissertation, Ottawa-Carleton Institute of Biology: Ottawa, ON, Canada, 2002. [Google Scholar]
- Brisson, LF; Tenhaken, R; Lamb, C. Function of oxidative cross-linking of cell wall structural proteins in plant disease resistance. Plant Cell 1994, 6, 1703–1712. [Google Scholar]
- Richardson, A; Stewart, D; McDougall, GJ. Identification and partial characterization of a coniferyl alcohol oxidase from lignifying xylem of Sitka spruce (Picea sitchensis). Planta 1997, 203, 35–43. [Google Scholar]
- Fry, SC. Cross-linking of matrix polymers in the growing cell walls of angiosperms. Ann. Rev. Plant Physiol 1986, 37, 165–186. [Google Scholar]
- Ramputh, AI; Arnason, JT; Cass, L; Simmonds, JA. Reduced herbivory of the European corn borer (Ostrinia nubilalis) on corn transformed with germin, a wheat oxalate oxidase gene. Plant Sci 2002, 162, 431–440. [Google Scholar]
- Mao, J; Burt, AJ; Ramputh, A-I; Simmonds, J; Cass, L; Hubbard, K; Miller, S; Altosaar, I; Arnason, JT. Diverted secondary metabolism and improved resistance to European corn borer (Ostrinia nubilalis) in maize (Zea mays L.) transformed with wheat oxalate oxidase. J. Agric. Food Chem 2007, 55, 2582–2589. [Google Scholar]
- Santiago, R; Butrón, A; Arnason, JT; Reid, LM; Souto, XC; Malvar, RA. Putative role of pith cell wall phenylpropanoids in Sesamia nonagrioides (Lepidoptera: Noctuidae) resistance. J. Agric. Food Chem 2006, 54, 2274–2279. [Google Scholar]
- Santiago, R; Butrón, A; Reid, LM; Arnason, JT; Sandoya, G; Souto, XC; Malvar, RA. Diferulate content of maize sheaths is associated with resistance to the Mediterranean corn borer Sesamia nonagrioides (Lepidoptera: Noctuidae). J. Agric. Food Chem 2006, 54, 9140–9144. [Google Scholar]
- Santiago, R; Sandoya, G; Butrón, A; Barros, J; Malvar, RA. Changes in phenolic concentrations during recurrent selection for resistance to the Mediterranean corn borer (Sesamia nonagrioides Lef.). J. Agric. Food Chem 2008, 56, 8017–8022. [Google Scholar]
- Markham, RH; Bosque-Pérez, NA; Borgemeister, C; Meikle, WG. Developing pest management strategies for the maize weevil, Sitophilus zeamais, and the large grain borer, Prostephanus truncatus, in the humid and sub-humid tropics. FAO Plant Prot. Bull 1994, 42, 125–136. [Google Scholar]
- Tigar, BJ; Osborne, PE; Key, GE; Flores-S, ME; Vazquez, AM. Insect pest associated with rural maize stores in Mexico with particular reference to Prostephanus truncatus (Coleoptera: Bostrichidae). J. Stored Prod. Res 1994, 30, 267–281. [Google Scholar]
- Dobie, P. The contribution of the Tropical Stored Products Centre to the study of insect resistance in stored maize. Trop. Stored Prod. Inf 1977, 34, 7–22. [Google Scholar]
- Serratos, A; Arnason, JT; Nozzolillo, C; Lambert, JDH; Philogène, BJR; Fulcher, G; Davidson, K; Peacock, L; Atkinson, J; Morand, P. Factors contributing to resistance of exotic maize populations to maize weevil, Sitophilus zeamais. J. Chem. Ecol 1987, 13, 751–762. [Google Scholar]
- Tipping, PW; Legg, DE; Rodriguez, JG; Poneleit, CG. Influence of maize pericarp surface relief on the resistance to the maize weevil. J. Kans. Entomol. Soc 1988, 61, 237–241. [Google Scholar]
- Arnason, JT; Baum, B; Gale, J; Lambert, JDH; Bergvinson, D; Philogène, BJR; Serratos, JA; Mihm, JA; Jewell, DC. Variation in resistance of Mexican landraces of maize to maize weevil Sitophilus zeamais, in relation to taxonomic and biochemical parameters. Euphytica 1994, 74, 227–236. [Google Scholar]
- Arnason, JT; Conilh de Beyssac, B; Philogène, BJR; Bergvinson, D; Serratos, JA; Mihm, JA. Mechanism of resistance in maize grain to the maize weevil and the larger grain borer. In Insect Resistance Maize: Recent Advances and Utilization; Proceedings of an International Symposium; Mexico, DF, Ed.; Mexico; 23 November–3 December, 1997; pp. 91–95. [Google Scholar]
- Classen, D; Arnason, JT; Serratos, JA; Lambert, JDH; Nozzolillo, C; Philogène, BJR. Correlation of phenolic acid content of maize to resistance to Sitophilus zeamais, the maize weevil, in CIMMYT’s collections. J. Chem. Ecol 1990, 16, 301–315. [Google Scholar]
- García-Lara, S; Bergvinson, D; Burt, AJ; Ramputh, A-I; Díaz-Pontones, DM; Arnason, JT. The role of pericarp cell wall components in maize weevil resistance. Crop Sci 2004, 44, 1546–1552. [Google Scholar]
- Sutton, JC. Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Can. J. Plant Pathol 1982, 4, 195–209. [Google Scholar]
- Mirocha, CJ; Pathre, SY; Schauerhamer, B; Christensen, CM. Natural occurrence of Fusarium toxins in feedstuff. Appl. Environ. Microbiol 1976, 32, 553–556. [Google Scholar]
- Pitt, JI. Toxigenic fungi and mycotoxins. Br. Med. Bull 2000, 56, 184–192. [Google Scholar]
- Steyn, PS; Stander, MA. Mycotoxins as causal factors of diseases in humans. J. Toxicol. Toxin Rev 1999, 18, 229–243. [Google Scholar]
- Reid, LM; Mather, DE; Arnason, JT; Hamilton, RI; Bolton, AT. Changes in phenolic constituents of maize silk infected with Fusarium graminearum. Can. J. Bot 1992, 70, 1697–1702. [Google Scholar]
- Assabgui, RA; Reid, LM; Hamilton, RI; Arnason, JT. Correlation of kernel (E)-ferulic acid content of maize with resistance to Fusarium graminearum. Phytopathology 1993, 83, 949–953. [Google Scholar]
- Miller, JD; Miles, M; Davis, AF. Kernel concentrations of 4-acetylbenzoxazolin-2-one and diferuloylputrescine in maize genotypes and Gibberella ear rot. J. Agric. Food Chem 1997, 45, 4456–4459. [Google Scholar]
- Bily, AC; Reid, LM; Taylor, JH; Johnston, D; Malouin, C; Buró, AJ; Bakan, B; Regnault-Roger, C; Pauls, KP; Arnason, JT; Philogène, BJR. Dehydrodimers of ferulic acid in maize grain pericarp and aleurone: Resistance factors to Fusarium graminearum. Phytopathology 2003, 93, 712–719. [Google Scholar]
- Grabber, JH; Hatfield, RH; Ralph, J. Diferulate cross-links impede the enzymatic degradation of non-lignified maize walls. J. Sci. Food Agric 1998, 77, 193–200. [Google Scholar]
- Obel, N; Porchia, AC; Scheller, HV. Dynamic changes in cell wall polysaccharides during wheat seedling development. Phytochemistry 2002, 60, 603–610. [Google Scholar]
- Manka, M. Cellulolytic and pectolytic activity of Fusarium isolates pathogenic to corn seedlings. Acta Microbiol. Pol 1980, 30, 25–32. [Google Scholar]
- Huang, Z; White, DG; Payne, GA. Corn seed proteins inhibitory to Aspergillus flavus and aflatoxin biosynthesis. Phytopathology 1997, 87, 622–627. [Google Scholar]
- Bakan, B; Bily, AC; Melcion, D; Cahagnier, B; Regnault-Roger, C; Philogène, BJR; Richard-Molard, D. Possible role of plant phenolics in the production of trichothecenes by Fusarium graminearum strains on different fractions of maize kernels. J. Agric. Food Chem 2003, 51, 2826–2831. [Google Scholar]
- Snijders, CH. A breeding for resistance to Fusarium in wheat and maize. In Mycotoxins in Grain: Compounds Other than Aflatoxin; Miller, JD, Trenholm, HL, Eds.; Eagan Press: St. Paul, MN, USA, 1994. [Google Scholar]
- Gilberton, RL; Brown, WM; Ruppel, EG. Prevalence and virulence of Fusarium spp. associated with stalk rot of corn in Colorado. Plant Dis 1985, 69, 1065–1068. [Google Scholar]
- Marasas, WFO; Nelson, PE; Tousson, TA. Toxigenic Fusarium Species: Identity and Mycotoxicology; The Pennsylvania State University Press: Pennsylvania, USA, 1984. [Google Scholar]
- Miller, JD; Young, JC; Trenholm, HL. Fusarium toxins in field corn. I. Time course of fungal growth and production of deoxynivalenol and other mycotoxins. Can. J. Bot 1983, 61, 3080–3087. [Google Scholar]
- Young, JC; Miller, JD. Appearance of fungus, ergosterol and Fusarium mycotoxins in the husk, axial stem and stalk after ear inoculation of field corn. Can. J. Plant Sci 1985, 65, 47–53. [Google Scholar]
- Purushothaman, D. Changes in phenolic compounds in rice cultivars as influenced by Xanthomonas oryzae. Il Riso 1976, 25, 88–91. [Google Scholar]
- Baranowski, JD; Davidson, PM; Nagel, CW; Branen, AL. Inhibition of Saccharomyces cerevisiae by naturally occurring hydroxycinnamates. J. Food Sci 1980, 45, 592–594. [Google Scholar]
- Kasenberg, TR; Traquair, JA. Effects of phenolics on growth of Fusarium oxysporum f. spradicis-lycopersici in vitro. Can. J. Bot 1988, 66, 1174–1177. [Google Scholar]
- McKeehen, JD; Busch, RH; Fulcher, RG. Evaluation of wheat (Triticum aestivum L.) phenolic acids during grain development and their contribution to Fusarium resistance. J. Agric. Food Chem 1999, 47, 1476–1482. [Google Scholar]
- Fry, SC. Intracellular feruloylation of pectic polysaccharides. Planta 1987, 171, 205–211. [Google Scholar]
- Santiago, R; Reid, LM; Arnason, JT; Zhu, XY; Martinez, N; Malvar, RA. Phenolics in maize genotypes differing in susceptibility to Gibberella stalk rot (Fusarium graminearum Schwabe). J. Agric. Food Chem 2007, 55, 5186–5193. [Google Scholar]
- Ikegawa, T; Mayama, S; Nakayashiki, H; Kato, H. Accumulation of diferulic acid during the hypersensitive response of oat leaves to Puccinia coronata f. sp. avena and its role in the resistance of oat tissues to cell wall degrading enzymes. Physiol. Mol. Plant Pathol 1996, 48, 245–256. [Google Scholar]
- Matern, U; Grimmig, B. Polyphenols in plant pathology. In Polyphenolic Phenomena; Scalbert, A, Ed.; INRA Editions: Paris, France, 1993. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Santiago, R.; Malvar, R.A. Role of Dehydrodiferulates in Maize Resistance to Pests and Diseases. Int. J. Mol. Sci. 2010, 11, 691-703. https://doi.org/10.3390/ijms11020691
Santiago R, Malvar RA. Role of Dehydrodiferulates in Maize Resistance to Pests and Diseases. International Journal of Molecular Sciences. 2010; 11(2):691-703. https://doi.org/10.3390/ijms11020691
Chicago/Turabian StyleSantiago, Rogelio, and Rosa A. Malvar. 2010. "Role of Dehydrodiferulates in Maize Resistance to Pests and Diseases" International Journal of Molecular Sciences 11, no. 2: 691-703. https://doi.org/10.3390/ijms11020691
APA StyleSantiago, R., & Malvar, R. A. (2010). Role of Dehydrodiferulates in Maize Resistance to Pests and Diseases. International Journal of Molecular Sciences, 11(2), 691-703. https://doi.org/10.3390/ijms11020691