Mechanisms Regulating Skin Pigmentation: The Rise and Fall of Complexion Coloration

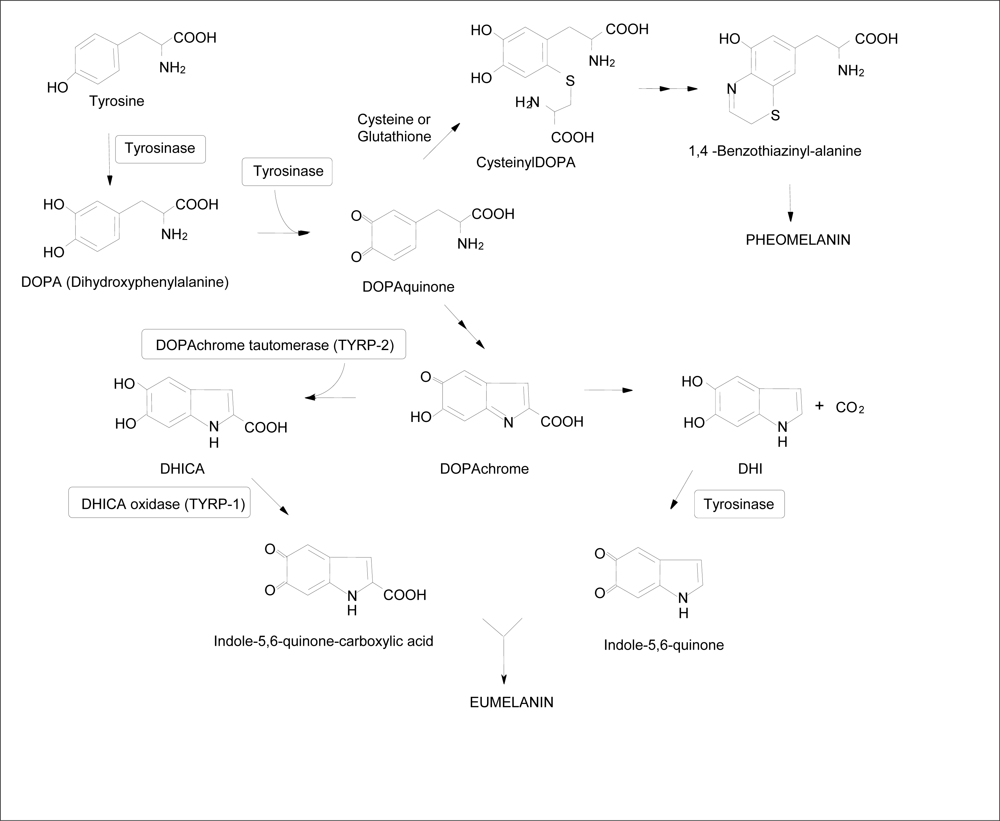
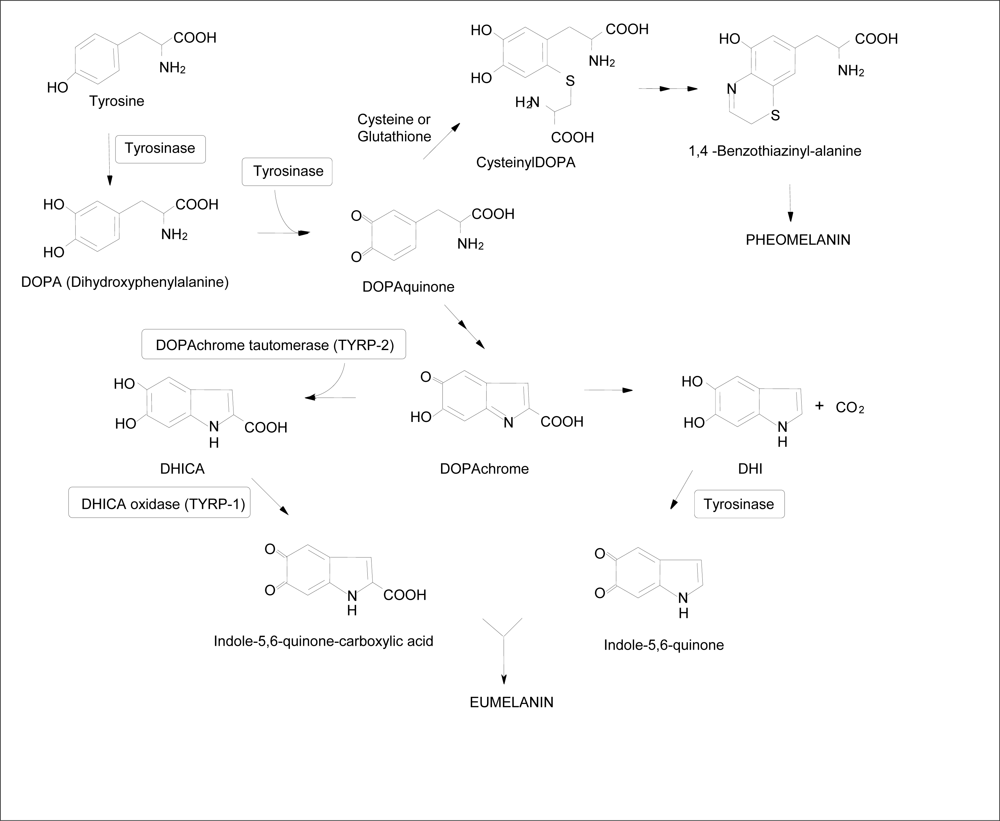{kind=link}
Abstract
:1. Introduction
2. Transcriptional Regulation of Melanogenic Enzymes
3. Post-Translational Modification of Melanogenic Enzymes
4. Attenuation of Tyrosinase and Related Melanogenic Enzymes Catalytic Activity
4.1. Hydroquinone
4.2. Monobenzylether
4.3. Arbutin and Deoxyarbutin
4.4. Mequinol
4.5. N-Acetyl-4-S-Cysteaminylphenol
4.6. Kojic Acid
4.7. Azelaic Acid
4.8. Gentisic Acid
4.9. Flavonoid-like Agents
4.9.1. Aloesin
4.9.2. Hydroxystilbene
4.9.3. Licorice Extract
4.10. Antioxidants and Redox Agents
4.10.1. l-Ascorbic Acid and Magnesium-l-Ascorbyl-2-Phosphate
4.10.2. Alpha Tocopherol and Alpha Tocopherol Ferulate
5. Interruption of Melanosome Transfer
5.1. Centaureidin and Methylophiopogonanone B
5.2. Niacinamide
5.3. PAR-2 Inhibitors
5.4. Lectins and Neoglycoproteins
6. Acceleration of Epidermal Turnover and Desquamation
6.1. α-Hydroxyacids
6.2. Salicylic Acid
6.3. Linoleic Acid
6.4. Retinoids
7. Conclusions
References and Notes
- Turner, W; Taylor, J; Tchen, T. Melanosome formation in the goldfish: The role of multivesicular bodies. J. Ultra. Res 1975, 51, 16–31. [Google Scholar]
- Boissy, R; Huizing, M; Gahl, W. Biogenesis of melanosomes. In The Pigmentary System: Physiology and Pathophysiology, 2nd ed; Nordlund, JJ, Boissy, RE, Hearing, VJ, King, RA, Oetting, WS, Ortonne, JP, Eds.; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 155–170. [Google Scholar]
- Boissy, R. Melanosome transfer to and translocation in the keratinocyte. Exp. Dermatol 2003, 12, 5–12. [Google Scholar]
- Urabe, K; Nakayama, J; Hori, Y. Mixed epidermal and dermal hypermelanoses. In The Pigmentary System: Physiology and Pathophysiology; Nordlund, JJ, Boissy, RE, Hearing, VJ, King, RA, Oetting, WS, Ortonne, JP, Eds.; Oxford University Press: New York, NY, USA, 1998; pp. 909–913. [Google Scholar]
- Virador, V; Matsunaga, N; Matsunaga, J; Valencia, J; Oldham, R; Kameyama, K; Peck, G; Ferrans, V; Vieira, W; Abdel-Malek, Z; Hearing, V. Production of melanocyte-specific antibodies to human melanosomal proteins: Expression patterns in normal human skin and in cutaneous pigmented lesions. Pigment Cell Res 2001, 14, 289–297. [Google Scholar]
- Parvez, S; Kang, M; Chung, H-S; Cho, C; Hong, M-C; Shin, M-K; Bae, H. Survey and mechanism of skin depigmentation and lightening agents. Phytother. Res 2006, 20, 921–934. [Google Scholar]
- Ando, H; Kondoh, H; Ichihashi, M; Hearing, V. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Invest. Dermatol 2007, 127, 751–761. [Google Scholar]
- Sturm, R; Teasdale, R; Box, N. Human pigmentation genes: Identification, structure and consequences of polymorphic variation. Gene 2001, 277, 49–62. [Google Scholar]
- García-Borrón, J; Solano, F. Molecular anatomy of tyrosinase and its related proteins: Beyond the histidine-bound metal catalytic center. Pigment Cell Res 2002, 15. [Google Scholar]
- Briganti, S; Camera, E; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res 2003, 16, 101–110. [Google Scholar]
- Yamaguchi, Y; Brenner, M; Hearing, V. The regulation of skin pigmentation. J. Biol. Chem 2007, 282, 27557–27561. [Google Scholar]
- Ito, S. A chemist’s view of melanogenesis. Pigment Cell Res 2003, 16, 230–236. [Google Scholar]
- Shibahara, S; Takeda, K; Yasumoto, K; Udono, T; Watanabe, K; Saito, H; Takahashi, K. Microphthalmia-associated transcription factor (MITF): Multiplicity in structure, function, and regulation. J. Investig. Dermatol. Symp. Proc 2001, 6, 99–104. [Google Scholar]
- Tachibana, M; Takeda, K; Nobukuni, Y; Urabe, K; Long, J; Meyers, K; Aaronson, S; Miki, T. Ectopic expression of MITF, a gene for Waardenburg syndrome type 2, converts fibroblasts to cells with melanocyte characteristics. Nat. Genet 1996, 14, 50–54. [Google Scholar]
- Levy, C; Khaled, M; Fisher, D. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol. Med 2006, 12, 406–414. [Google Scholar]
- Takeda, K; Shibahara, S. Transcriptional regulation of melanocyte function. In The Pigmentary System: Physiology and Pathophysiology, 2nd ed; Nordlund, JJ, Boissy, RE, Hearing, VJ, King, RA, Oetting, WS, Ortonne, JP, Eds.; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 242–260. [Google Scholar]
- Steingrímsson, E; Copeland, N; Jenkins, N. Melanocytes and the microphthalmia transcription factor network. Annu. Rev. Genet 2004, 38, 365–411. [Google Scholar]
- Lin, C; Babiarz, L; Liebel, F; Roydon Price, E; Kizoulis, M; Gendimenico, G; Fisher, D; Seiberg, M. Modulation of microphthalmia-associated transcription factor gene expression alters skin pigmentation. J. Invest. Dermatol 2002, 119, 1330–1340. [Google Scholar]
- Yasumoto, K; Yokoyama, K; Shibata, K; Tomita, Y; Shibahara, S. Microphthalmia-associated transcription factor as a regulator for melanocyte-specific transcription of the human tyrosinase gene. Mol. Cell. Biol 1994, 14, 8058–8070. [Google Scholar]
- Solano, F; Briganti, S; Picardo, M; Ghanem, G. Hypopigmenting agents: An updated review on biological, chemical and clinical aspects. Pigment Cell Res 2006, 90, 550–571. [Google Scholar]
- Saha, B; Sing, S; Sarkar, C; Bera, R; Ratha, J; Tobin, D; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signalling to CREB. Pigment Cell Res 2006, 19, 595–605. [Google Scholar]
- Kim, D; Park, S; Park, K. Transforming growth factor-β1 decreases melanin synthesis via delayed extracellular signal-regulated kinase activation. Int. J. Biochem. Cell Biol 2004, 36, 1482–1491. [Google Scholar]
- Yang, G; Li, Y; Nishimura, E; Xin, H; Zhou, A; Guo, Y; Dong, L; Denning, M; Nickoloff, B; Cui, R. Inhibition of PAX3 by TGF-β modulates melanocyte viability. Mol. Cell 2008, 32, 554–563. [Google Scholar]
- Englaro, W; Bertolotto, C; Busca, R; Brunet, A; Pages, G; Ortonne, J; Ballotti, R. Inhibition of the mitogen-activated protein kinase pathway triggers B16 melanoma cell differentiation. J. Biol. Chem 1998, 273, 9966–9970. [Google Scholar]
- Kim, D; Park, S; Kwon, S; Park, E; Huh, C; Youn, SW; Park, K. Sphingosylphosphorylcholine-induced ERK activation inhibits melanin synthesis in human melanocytes. Pigment Cell Res 2006, 19, 146–153. [Google Scholar]
- Kim, D; Hwang, E; Lee, J; Kim, S; Kwon, S; Park, K. Sphingosine-1-phosphate decreases melanin synthesis via sustained ERK activation and subsequent MITF degradation. J. Cell Sci 2003, 116, 1699–1706. [Google Scholar]
- Kim, D; Kim, S; Chung, J; Kim, K; Eun, H; Park, K. Delayed ERK activation by ceramide reduces melanin synthesis in human melanocytes. Cell. Signal 2002, 14, 779–785. [Google Scholar]
- Hemesath, T; Price, E; Takemoto, C; Badalian, T; Fisher, D. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar]
- Wu, M; Hemesath, T; Takemoto, C; Horstmann, M; Wells, A; Price, E; Fisher, DZ; Fisher, DE. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev 2000, 14, 301–312. [Google Scholar]
- Xu, W; Gong, L; Haddad, M; Bischof, O; Campisi, J; Yeh, E; Medrano, E. Regulation of microphthalmia-associated transcription factor MITF protein levels by association with the ubiquitin-conjugating enzyme hUBC9. Exp. Cell Res 2000, 255, 135–143. [Google Scholar]
- Kim, D; Park, S; Kwon, S; Youn, S; Park, K. Effects of lysophosphatidic acid on melanogenesis. Chem. Phys. Lipids 2004, 127, 199–206. [Google Scholar]
- Widlund, H; Fisher, D. Microphthalamia-associated transcription factor: A critical regulator of pigment cell development and survival. Oncogene 2003, 22, 3035–3041. [Google Scholar]
- McGill, G; Horstmann, M; Widlund, H; Du, J; Motyckova, G; Nishimura, E; Lin, YL; Ramaswamy, S; Avery, W; Ding, H; Jordan, S; Jackson, I; Korsmeyer, S; Golub, T; Fisher, D. Bcl2 regulation by the melanocyte master regulator Mitf modulates lineage survival and melanoma cell viability. Cell 2002, 109, 707–718. [Google Scholar]
- Buscà, R; Berra, E; Gaggioli, C; Khaled, M; Bille, K; Marchetti, B; Thyss, R; Fitsialos, G; Larribère, L; Bertolotto, C; Virolle, T; Barbry, P; Pouysségur, J; Ponzio, G; Ballotti, R. Hypoxia-inducible factor 1α is a new target of microphthalmia-associated transcription factor (MITF) in melanoma cells. J. Cell Biol 2005, 170, 49–59. [Google Scholar]
- Hornyak, T; Jiang, S; Guzman, E; Scissors, B; Tuchinda, C; He, H; Neville, J; Strickland, F. Mitf dosage as a primary determinant of melanocyte survival after ultraviolet irradiation. Pigment Cell Melanoma Res 2009, 22, 307–318. [Google Scholar]
- Negriou, G; Branza-Nichita, N; Petrescu, A; Dwek, R; Petrescu, S. Protein specific N-glycosylation of tyrosinase and tyrosinase related protein-1 in B16 mouse melanoma cells. Biochem. J 1999, 344, 659–665. [Google Scholar]
- Branza-Nichita, N; Petrescu, A; Negroiu, G; Dwek, R; Petrescu, S. N-Glycosylation processing and glycoprotein folding Lessons from the tyrosinase-related proteins. Chem. Rev 2000, 100, 4697–4711. [Google Scholar]
- Imokawa, G; Mishima, Y. Functional analysis of tyrosinase isozymes of cultured malignant melanoma cells during the recovery period following interrupted melanogenesis induced by glycosylation inhibitors. J. Invest. Dermatol 1984, 83, 196–201. [Google Scholar]
- Mishima, Y; Imokawa, G. Selective aberration and pigment loss in melanosomes of malignant melanoma cells in vitro by glycosylation inhibitors: Premelanosomes as glycoproteins. J. Invest. Dermatol 1984, 81, 106–114. [Google Scholar]
- Terao, M; Tomita, K; Oki, T; Tabe, L; Gianni, M; Garattini, E. Inhibition of melanogenesis by BMY-28565, a novel compound depressing tyrosinase activity in B16 melanoma cells. Biochem. Pharmacol 1992, 43, 183–189. [Google Scholar]
- Land, E; Ramsden, C; Riley, P. Toxicological aspects of melanin and melanogenesis. In The Pigmentary System: Physiology and Pathophysiology, 2nd ed; Nordlund, JJ, Boissy, RE, Hearing, VJ, King, RA, Oetting, WS, Ortonne, JP, Eds.; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 354–394. [Google Scholar]
- Choi, H; Ahn, S; Chang, H; Cho, N; Joo, K; Lee, B; Chang, I; Hwang, J. Influence of N-glycan processing disruption on tyrosinase and melanin synthesis in HM3KO melanoma cells. Exp. Dermatol 2006, 16, 110–117. [Google Scholar]
- Franchi, J; Coutadeur, M; Marteau, C; Mersel, M; Kupferberg, A. Depigmenting effects of calcium d-pantetheine-s-sulfonate on human melanocytes. Pigment Cell Res 2000, 35, 165–171. [Google Scholar]
- Maresca, V; Flori, E; Cardinali, G; Briganti, S; Lombardi, D; Mileo, A; Paggi, M; Picardo, M. Ferritin light chain down-modulation generates depigmentation in human metastatic melanoma cells by influencing tyrosinase maturation. J. Cell. Physiol 2006, 206, 843–848. [Google Scholar]
- Imokawa, G. Analysis of initial melanogenesis including tyrosinase transfer and melanosome differentiation through interrupted melanization by glutathione. J. Invest. Dermatol 1989, 93, 100–107. [Google Scholar]
- Villarama, C; Maibach, H. Glutathione as a depigmenting agent: An overview. Int. J. Cosmet. Sci 2005, 27, 147–153. [Google Scholar]
- Nordlund, J; Grimes, P; Ortonne, J. The safety of hydroquinone. J. Eur. Acad. Dermatol. Venereol 2006, 20, 781–787. [Google Scholar]
- Halder, R; Nordlund, J. Topical treatment of pigmentary disorders. In The Pigmentary System: Physiology and Pathophysiology, 2nd ed; Nordlund, JJ, Boissy, RE, Hearing, VJ, King, RA, Oetting, WS, Ortonne, JP, Eds.; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 1165–1174. [Google Scholar]
- Draelos, Z. Skin lightening preparations and the hydroquinone controversy. Dermatol. Ther 2007, 20, 308–313. [Google Scholar]
- Picardo, M; Carrera, M. New and experimental treatments of cloasma and other hypermelanoses. Dermatol. Clin 2007, 25, 353–362. [Google Scholar]
- Badreshia-Bansal, S; Draelos, Z. Insight into skin lightening cosmeceuticals for women of color. J. Drugs Dermatol 2007, 6, 32–39. [Google Scholar]
- Ni-Komatsu, L; Tong, C; Chen, G; Brindzei, N; Orlow, S. Identification of quinolines that inhibit melanogenesis by altering tyrosinase family trafficking. Mol. Pharmacol 2008, 74, 1576–1586. [Google Scholar]
- Chakraborty, A; Funasaka, Y; Komoto, M; Ichihashi, M. Effect of arbutin on melanogenic proteins in human melanocytes. Pigment Cell Res 1998, 11, 206–212. [Google Scholar]
- Chawla, S; deLong, M; Visscher, M; Wickett, R; Manga, P; Boissy, R. Mechanism of tyrosinase inhibition by deoxyArbutin and its second-generation derivatives. Br. J. Dermatol 2008, 159, 1267–1274. [Google Scholar]
- Boissy, R; Visscher, M; DeLong, M. DeoxyArbutin: A novel reversible tyrosinase inhibitor with effective in vivo skin lightening potency. Exp. Dermatol 2005, 14, 601–608. [Google Scholar]
- Hamed, S; Sriwiriyanont, P; deLong, M; Visscher, M; Wickett, R; Boissy, R. Comparative efficacy and safety of deoxyarbutin, a new tyrosinase-inhibiting agent. J. Cos. Sci 2006, 57, 291–308. [Google Scholar]
- Petit, L; Piérard, G. Skin-lightening products revisited. Int. J. Cosmet. Sci 2003, 25, 169–181. [Google Scholar]
- Colby, S; Schwartzel, E; Huber, F; Highton, A; Altman, D; Epinette, W; Lyon, E. A promising new treatment for solar lentigines. J. Drugs Dermatol 2003, 2, 147–152. [Google Scholar]
- Keeling, J; Cardona, L; Benitez, A; Epstein, R; Rendon, M. Mequinol 2%/tretinoin 0.01% topical solution for the treatment of melasma in men: A case series and review of the literature. Cutis 2008, 81, 179–183. [Google Scholar]
- Jimbow, K. N-Acetyl-4-S-Cysteaminylphenol as a new type of depigmenting agent for the melanoderma of patients with melasma. Arch. Dermatol 1991, 127, 1528–1534. [Google Scholar]
- Ferguson, J; Rogers, PM; Kelland, LR; Robins, DJ. Synthesis and antimelanoma activity of sterically congested tertiary amide analogues of N-acetyl-4-S-cysteaminylphenol. Oncol. Res 2005, 15, 87–94. [Google Scholar]
- Kim, D; Hwang, J; Baek, H; Kim, K; Lee, B; Chang, I; Kang, H; Lee, O. Development of 5-[(3-aminopropyl)phosphinooxy]-2-(hydroxymethyl)-4H-pyran-4-one as a novel whitening agent. Chem. Pharm. Bull 2003, 51, 113–116. [Google Scholar]
- Kim, Y; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell. Mol. Life Sci 2005, 62, 1707–1723. [Google Scholar]
- Parvez, S; Kang, M; Chung, H; Bae, H. Naturally occuring tyrosinase inhibitors: Mechanism and application in skin health, cosmetics and agriculture industries. Phytother. Res 2007, 21, 805–816. [Google Scholar]
- Grimes, P. Melasma: Etiologic and therapeutic considerations. Arch. Dermatol 1995, 131, 1453–1457. [Google Scholar]
- Baliña, L; Graupe, K. The treatment of melasma 20% azeiaic acid versus 4% hydroquinone cream. Int. J. Dermatol 1991, 30, 893–895. [Google Scholar]
- Fitton, A; Goa, K. Azelaic acid: A review of its pharmacological properties and therapeutic efficacy in acne and hyperpigmentary skin disorders. Drugs 1991, 41, 780–798. [Google Scholar]
- Kubo, I; Kinst-Hori, I; Kubo, Y; Yamagiwa, Y; Kamikawa, T; Haraguchi, H. Molecular design of antibrowing agents. J. Agric. Food Chem 2000, 48, 1393–1399. [Google Scholar]
- Jones, K; Hughes, J; Hong, M; Jia, Q; Orndorff, S. Modulation of melanogenesis by aloesin: A competitive inhibitor of tyrosinase. Pigment Cell Res 2002, 15, 335–340. [Google Scholar]
- Zhu, W; Gao, J. The use of botanical extracts as topical skin-lightening agents for the improvement of skin pigmentation disorders. J. Investig. Dermatol. Symp. Proc 2008, 13, 20–24. [Google Scholar]
- Jin, Y; Lee, S; Chung, M; Park, J; Park, YI; Cho, T; Lee, S. Aloesin and arbutin inhibit tyrosinase activity in a synergistic manner via a different action mechanism. Arch. Pharm. Res 1999, 22, 232–236. [Google Scholar]
- Kim, D; Kim, J; Baek, D; Seo, J; Kho, Y; Oh, T; Lee, C. Enhancement of tyrosinase inhibition of the extract of veratrum patulum using cellulase. Biotechnol. Bioeng 2004, 87, 849–854. [Google Scholar]
- Frémont, L. Biological effects of resveratrol. Life Sci 2000, 66, 663–673. [Google Scholar]
- Kim, Y; Yun, J; Lee, C; Lee, H; Min, K; Kim, Y. Oxyresveratrol and hydroxystilbene compounds inhibitory effect on tyrosinase and mechanisms of action. J. Biol. Chem 2002, 227, 16340–16344. [Google Scholar]
- Newton, R; Cook, A; Roberts, D; Leonard, J; Sturm, R. Post-transcriptional regulation of melanin biosynthetic enzymes by cAMP and resveratrol in human melanocytes. J. Invest. Dermatol 2007, 127, 2216–2227. [Google Scholar]
- Yamakoshi, J; Otsuka, F; Sano, A; Tokutake, S; Saito, M. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a proanthocyanidin-rich extract from grape seeds. Pigment Cell Res 2003, 16, 629–638. [Google Scholar]
- Hachiya, A; Kobayashi, A; Yoshida, Y; Kitahara, T; Takema, Y; Imokawa, G. Biphasic expression of two paracrine melanogenic cytokines, stem cell factor and endothelin-1, in ultraviolet b-induced human melanogenesis. Am. J. Pathol 2004, 165, 2099–2109. [Google Scholar]
- Berson, D. Natural antioxidants. J. Drugs Dermatol 2008, 7, s7–s12. [Google Scholar]
- Karg, E; Odh, G; Wittbjer, A; Rosengren, E; Rorsman, H. Hydrogen peroxide as inducer of elevated tyrosinase level in melanoma cells. J. Invest. Dermatol 1993, 100, 209s–213s. [Google Scholar]
- Farris, P. Topical vitamin C: A useful agent for treating photoaging and other dermatologic conditions. Dermatol. Surg 2005, 31, 814–818. [Google Scholar]
- Kameyama, K; Sakai, C; Kondoh, S; Yonemoto, K; Nishiyama, S; Tagawa, M; Murata, T; Ohnuma, T; Quigley, J; Dorsky, A; Bucks, D; Blanock, K. Inhibitory effect of magnesium l-ascorbyl-2-phosphate (VC-PMG) on melanogenesis in vitro and in vivo. J. Am. Acad. Dermatol 1996, 34, 29–33. [Google Scholar]
- Hakozaki, T; Takiwaki, H; Miyamoto, K; Sato, Y; Arase, S. Ultrasound enhanced skin-lightening effect of vitamin C and niacinamide. Skin Res. Technol 2006, 12, 105–113. [Google Scholar]
- Shimizu, K; Kondo, R; Sakai, K; Takeda, N; Nagahata, T; Oniki, T. Novel vitamin E derivative with 4-substituted resorcinol moiety has both antioxidant and tyrosinase inhibitory properties. Lipids 2001, 36, 1321–1326. [Google Scholar]
- Fukuzawa, K; Gebicki, J. Oxidation of α-tocopherol in micelles and liposomes by hydroxy, perhydroxy and superoxide free radicals. Arch. Biochem. Biophys 1983, 226, 242–251. [Google Scholar]
- Choi, C; Berson, D. Cosmeceuticals. Semin. Cutan. Med. Surg 2006, 25, 163–168. [Google Scholar]
- Funasaka, Y; Chakraborty, A; Komoto, M; Ohashi, A; Ichihashi, M. The depigmenting effect of α-tocopheryl ferulate on human melanoma cells. Br. J. Dermatol 1999, 141, 20–29. [Google Scholar]
- Minwalla, L; Zhao, Y; Cornelius, J; Babcock, G; Wickett, R; Le Poole, I; Boissy, R. Inhibition of melanosome transfer from melanocytes to keratinocytes by lectins and neoglycoproteins in an in vitro model system. Pigment Cell Res 2001, 14, 185–194. [Google Scholar]
- Ito, Y; Kanamaru, A; Tada, A. Centaureidin promotes dendrite retraction of melanocytes by activating Rho. Biochim. Biophys. Acta 2006, 1760, 487–494. [Google Scholar]
- Ito, Y; Kanamaru, A; Tada, A. Effects of methylophiopogonanone B on melanosome transfer and dendrite retraction. J. Dermatol. Sci 2006, 42, 68–70. [Google Scholar]
- Ito, Y; Kanamaru, A; Tada, A. A novel agent, methylophiopogonanone B, promotes Rho activation and tubulin depolymerization. Mol. Cell Biochem 2007, 297, 121–129. [Google Scholar]
- Lin, J; Chiang, H; Lin, Y; Wen, K. Natural products with skin–whitening effects. J.Food Drug Anal 2008, 16, 1–10. [Google Scholar]
- Hakozaki, T; Minwalla, L; Zhuang, J; Chhoa, M; Matsubara, A; Miyamoto, K; Greatens, A; Hillebrand, G; Bissett, D; Boissy, R. The effect of niacinamide on reducing cutaneous pigmentation and suppression of melanosome transfer. Br. J. Dermatol 2002, 147, 20–31. [Google Scholar]
- Greatens, A; Hakozaki, T; Koshoffer, A; Epstein, H; Schwemberger, S; Babcock, G; Bissett, D; Takiwaki, H; Arase, S; Wickett, R; Boissy, R. Effective inhibition of melanosome transfer to keratinocytes by lectins and niacinamide is reversible. Exp. Dermatol 2005, 14, 498–508. [Google Scholar]
- Bissett, D; Miyamoto, K; Sun, P; Li, J; Berge, C. Topical niacinamide reduces yellowing, wrinkling, red blotchiness, and hyperpigmented spots in aging facial skin. Int. J. Cosmet. Sci 2004, 26, 231–238. [Google Scholar]
- Bissett, D; Oblong, J; Berge, C. Niacinamide: A B vitamin that improves aging facial skin appearance. Dermatol. Surg 2005, 31, 860–865. [Google Scholar]
- Bissett, D. Topical niacinamide and barrier enhancement. Cutis 2002, 70S, 8–12. [Google Scholar]
- Bissett, D; Oblong, J; Saud, A; Berge, C; Trejo, A; Biedermann, K. Topical niacinamide provides skin aging appearance benefits while enhancing barrier function. J. Clin. Dermatol 2003, 32S, 9–18. [Google Scholar]
- Van Den Bossche, K; Naeyaert, J; Lambert, J. The quest for the mechanism of melanin transfer. Traffic 2006, 7, 769–778. [Google Scholar]
- Seiberg, M; Paine, C; Sharlow, E; Andrade-Gordon, P; Costanzo, M; Eisinger, M; Shapiro, S. The protease-activated receptor 2 regulates pigmentation via keratinocyte-melanocyte interactions. Exp. Cell Res 2000, 254, 25–32. [Google Scholar]
- Lin, C; Chen, N; Scarpa, R; Guan, F; Babiarz-Magee, L; Liebel, F; Li, W; Kizoulis, M; Shapiro, S; Seiberg, M. LIGR, a protease-activated receptor-2-derived peptide, enhances skin pigmentation without inducing inflammatory processes. Pigment Cell Melanoma Res 2008, 21, 172–183. [Google Scholar]
- Derian, C; Eckardt, A; Andrade-Gordon, P. Differential regulation of human keratinocyte growth and differentiation by a novel family of protease-activated receptors. Cell Growth Diff 1997, 8, 743–749. [Google Scholar]
- Marthinuss, J; Andrade-Gordon, P; Seiberg, M. A secreted serine protease can induce apoptosis in Pam212 keratinocytes. Cell Growth Diff 1995, 6, 807–816. [Google Scholar]
- Seiberg, M; Paine, C; Sharlow, E; Andrade-Gordon, P; Constanzo, M; Eisinger, M; Shapiro, S. Inhibition of melanosome transfer results in skin lightening. J. Invest. Dermatol 2000, 115, 162–167. [Google Scholar]
- Babiarz-Magee, L; Chen, N; Seiberg, M; Lin, C. The expression and activation of protease-activated receptor-2 correlate with skin color. Pigment Cell Res 2004, 17, 241–251. [Google Scholar]
- Sharlow, E; Paine, C; Babiarz, L; Eisinger, M; Shapiro, S; Seiberg, M. The protease-activated receptor-2 upregulates keratinocyte phagocytosis. J. Cell Sci 2000, 113, 3093–3101. [Google Scholar]
- Paine, C; Sharlow, E; Liebel, F; Eisinger, M; Shapiro, S; Seiberg, M. An alternative approach to depigmentation by soybean extracts via the inhibition of the PAR-2 pathway. J. Invest. Dermatol 2001, 116, 587–595. [Google Scholar]
- Scott, G; Deng, A; Rodriguez-Burford, C; Seiberg, M; Han, R; Babiarz, L; Grizzle, W; Bell, W; Pentland, A. Protease-activated receptor 2, a receptor involved in melanosome transfer, is upregulated in human skin by ultraviolet irradiation. J. Invest. Dermatol 2001, 117, 1412–1420. [Google Scholar]
- Birk, Y. The Bowman-Birk inhibitor. Trypsin- and chymotrypsin-inhibitor from soybeans. Int. J. Pept. Protein Res 1985, 25, 113–131. [Google Scholar]
- Brenner, M; Hearing, V. Modifying skin pigmentation–approaches through intrinsic biochemistry and exogenous agents. Drug Discov. Today Dis. Mech 2008, 5, 189–199. [Google Scholar]
- Zhu, W; Zhang, R. Skin lightening agents. In Cosmetic Formulation of Skin Care Products Cosmetic Science and Technology Series; Draelos, ZD, Thaman, LA, Eds.; Taylor and Francis Group LLC: New York, NY, USA, 2006; Volume 30, pp. 205–218. [Google Scholar]
- Bowe, W; Shalita, A. Effective over-the-counter acne treatments. Semin. Cutan. Med. Surg 2008, 27, 170–176. [Google Scholar]
- Yamamoto, Y; Uede, K; Yonei, N; Kishioka, A; Ohtani, T; Furukawa, F. Effects of alpha-hydroxy acids on the human skin of Japanese subjects: The rationale for chemical peeling. J. Dermatol 2006, 1, 16–22. [Google Scholar]
- Ando, H; Funasaka, Y; Oka, M; Ohashi, A; Furumura, M; Matsunaga, J; Matsunaga, N; Hearing, V; Ichihashi, M. Possible involvement of proteolytic degradation of tyrosinase in the regulatory effect of fatty acids on melanogenesis. J. Lipid Res 1999, 40, 1312–1316. [Google Scholar]
- Halaban, R; Cheng, E; Zhang, Y; Moellmann, G; Hanlon, D; Michalak, M; Setaluri, V; Hebert, D. Aberrant retention of tyrosinase in the endoplasmic reticulum mediates accelerated degradation of the enzyme and contributes to the dedifferentiated phenotype of amelanotic melanoma cells. Proc. Natl. Acad. Sci. USA 1997, 94, 6210–6215. [Google Scholar]
- Ando, H; Ryu, A; Hashimoto, A; Oka, M; Ichihashi, M. Linoleic acid and α-linolenic acid lightens ultraviolet-induced hyperpigmentation of the skin. Arch. Dermatol. Res 1998, 290, 375–381. [Google Scholar]
- Nair, X; Parah, P; Suhr, L; Tramposch, K. Combination of 4-hydroxyanisole and all trans retinoic acid produces synergistic skin depigmentation in swine. J. Invest. Dermatol 1993, 101, 145–149. [Google Scholar]
- Berardesca, E; Ardigò, M; Berardesca, M; Cameli, N. Melasma: Current and future treatments. Expert Rev. Dermatol 2008, 3, 187–193. [Google Scholar]
- Lei, T; Virador, V; Vieira, W; Hearing, V. A melanocyte–keratinocyte coculture model to assess regulators of pigmentation in vitro. Anal. Biochem 2002, 305, 260–268. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ebanks, J.P.; Wickett, R.R.; Boissy, R.E. Mechanisms Regulating Skin Pigmentation: The Rise and Fall of Complexion Coloration. Int. J. Mol. Sci. 2009, 10, 4066-4087. https://doi.org/10.3390/ijms10094066
Ebanks JP, Wickett RR, Boissy RE. Mechanisms Regulating Skin Pigmentation: The Rise and Fall of Complexion Coloration. International Journal of Molecular Sciences. 2009; 10(9):4066-4087. https://doi.org/10.3390/ijms10094066
Chicago/Turabian StyleEbanks, Jody P., R. Randall Wickett, and Raymond E. Boissy. 2009. "Mechanisms Regulating Skin Pigmentation: The Rise and Fall of Complexion Coloration" International Journal of Molecular Sciences 10, no. 9: 4066-4087. https://doi.org/10.3390/ijms10094066
APA StyleEbanks, J. P., Wickett, R. R., & Boissy, R. E. (2009). Mechanisms Regulating Skin Pigmentation: The Rise and Fall of Complexion Coloration. International Journal of Molecular Sciences, 10(9), 4066-4087. https://doi.org/10.3390/ijms10094066