Variations in IC50 Values with Purity of Mushroom Tyrosinase

Abstract
:1. Introduction
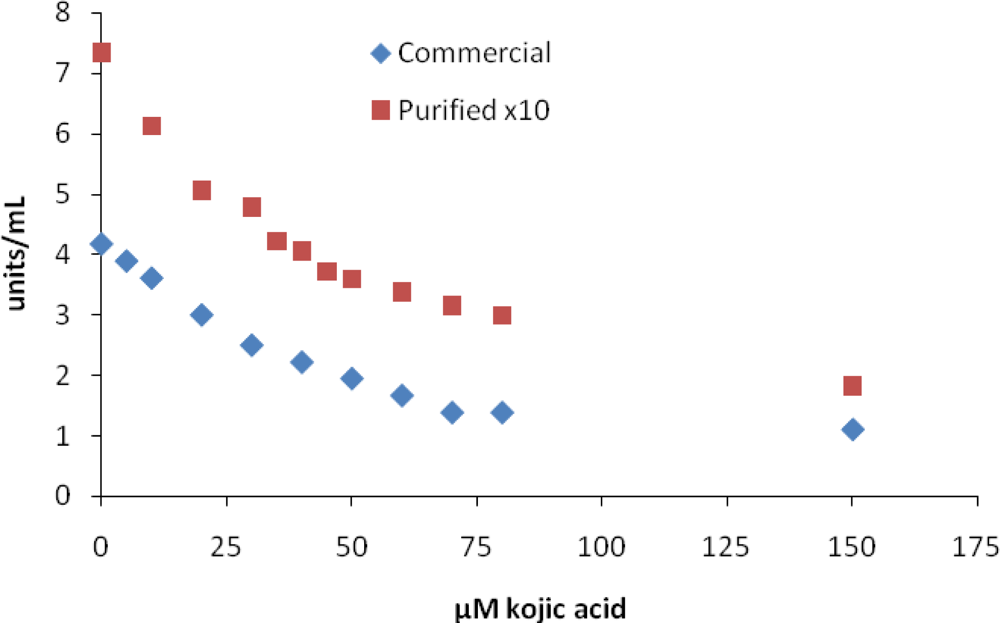
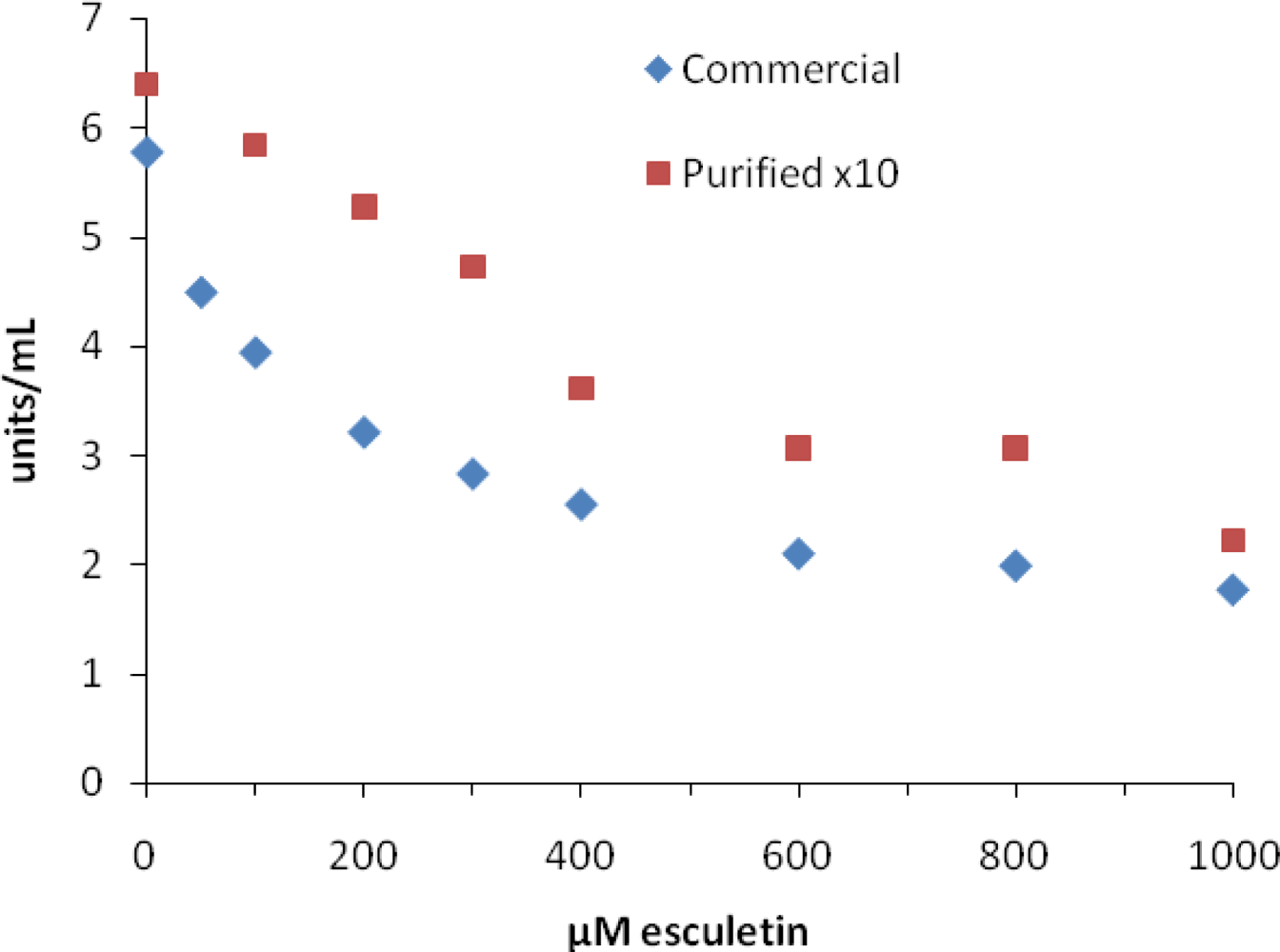
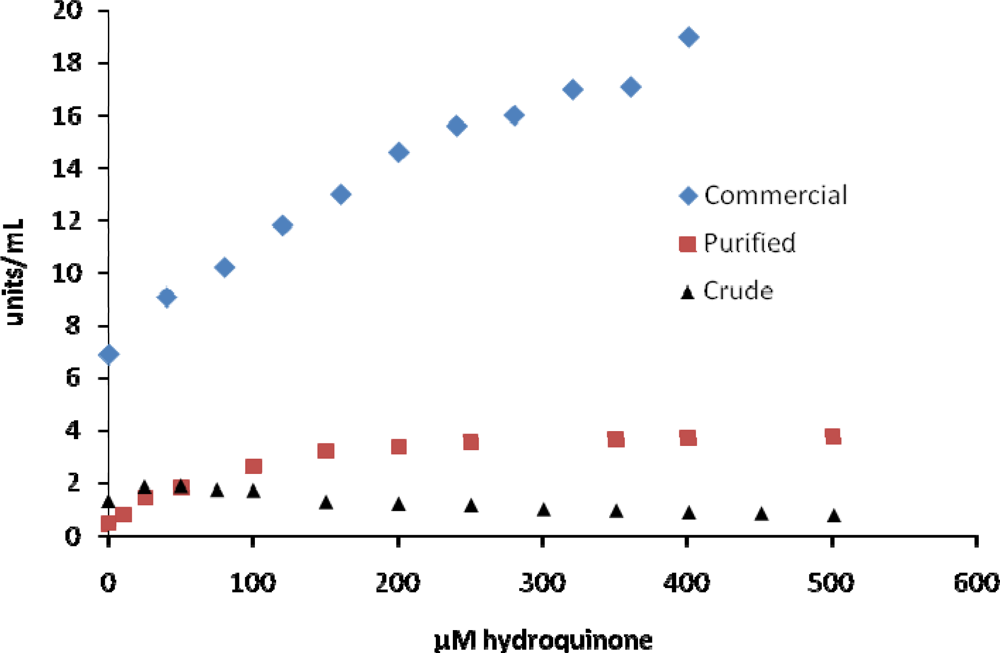
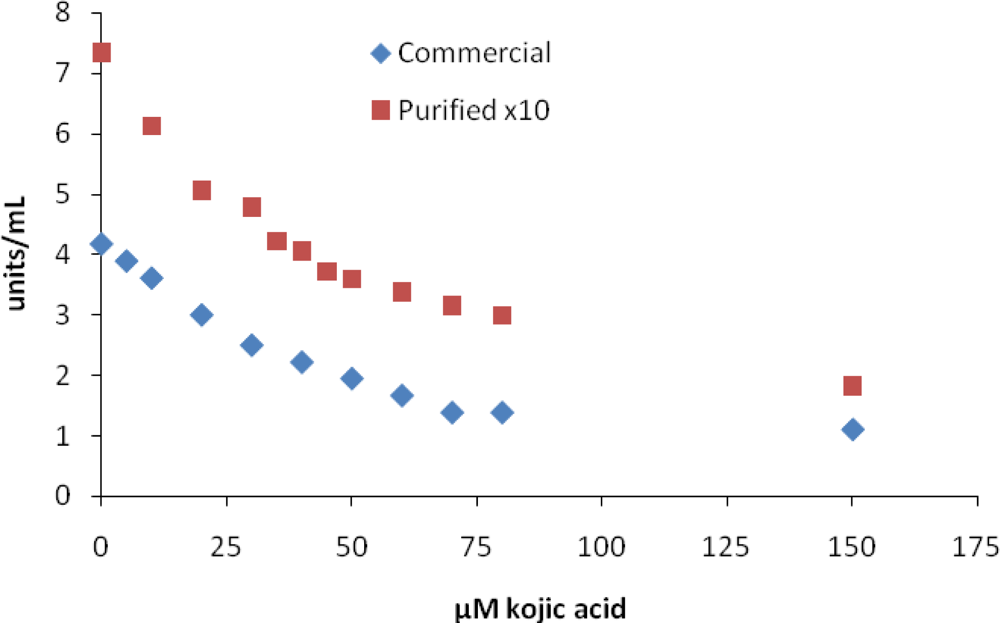
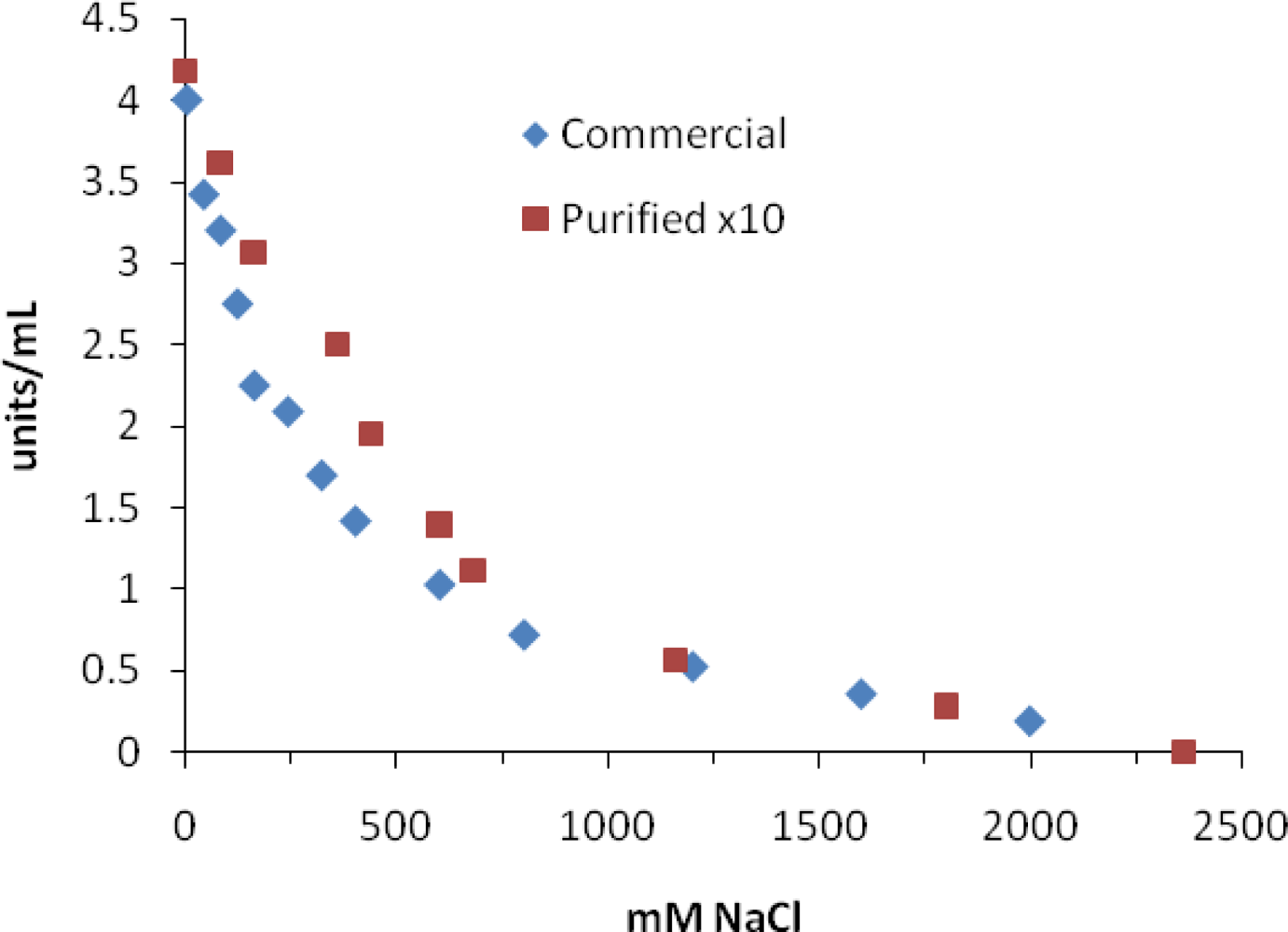
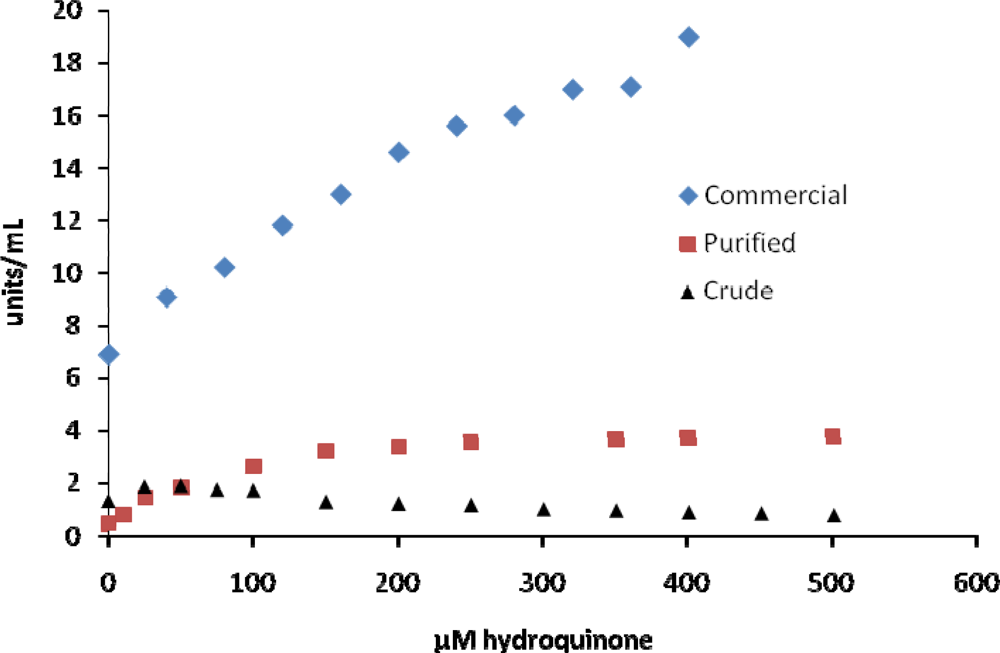
2. Results and Discussion
3. Experimental Section
3.1. Materials
3.2. Methods
3.2.1. Crude Extract Tyrosinase Preparation
3.2.2. Enzyme Purification
3.2.3. Enzyme Assays
3.2.4. IC50 Determinations
4. Conclusions
Acknowledgments
Abbreviations:
| ATTM | ammonium tetrathiomolybdate |
| biphenol | 4,4’-dihydroxy-biphenyl |
| DOPA | dihydroxyphenylalanine |
| DMSO | dimethyl sulfoxide |
| EDTA | ethylenediaminetetraacetic acid |
| 4HR | 4-hexylresorcinol |
| MT | mushroom tyrosinase |
| PMSF | phenylmethylsulfonyl fluoride |
| SDS | sodium dodecylsulfate |
| SHAM | salicyl-hydroxamic acid |
References and Notes
- Flurkey, WH; Inlow, JK. Proteolytic processing of polyphenol oxidase from plants and fungi. J. Inorg. Biochem 2008, 102, 2160–2170. [Google Scholar]
- Ando, H; Kondoh, H; Ichihashi, M; Hearing, VJ. Approaches to identify inhibitors of melanin biosynthesis via quality control of tyrosinase. J. Invest. Dermatol 2007, 127, 751–761. [Google Scholar]
- Mayer, AM. Polyphenol oxidases in plants and fungi: Going places? Phytochemistry 2006, 67, 2318–2331. [Google Scholar]
- Selinheimo, E; NiEidhin, D; Steffenson, C; Nielson, J; Lomascolo, A; Halaouli, S; Record, E; O’Beirne, D; Buchert, J; Kruus, K. Comparison of the characteristics of fungal and plant tyrosinases. J. Biotechnol 2007, 130, 471–480. [Google Scholar]
- Gandia-Herrero, F; Escribano, J; Garcia-Carmona, F. Betaxanthins as substrates for tyrosinase. An approach to the role of tyrosinase in the biosynthetic pathway of betalains. Plant Physiol 2005, 138, 421–432. [Google Scholar]
- Yoruk, R; Marshall, M. Physicochemical properties and function of plant polyphenol oxidase: A review. J. Food Biochem 2003, 27, 361–422. [Google Scholar]
- Walker, JRL; Ferrar, PH. The control of enzymic browning in foods. Chem. Indust 1995, 16, 836–839. [Google Scholar]
- Kim, J; Marshall, MR; Wei, C. Polyphenoloxidase. In Sea Food Enzymes; Haard, NF, Simpson, BK, Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 271–315. [Google Scholar]
- Rescigno, A; Zucca, P; Flurkey, A; Inlow, J; Flurkey, WH. Identification and discrimination between some contaminant enzyme activities in commercial preparations of mushroom tyrosinase. Enzyme Micro. Tech 2007, 41, 620–627. [Google Scholar]
- Flurkey, A; Cooksey, J; Reddy, A; Spoonmore, K; Rescigno, A; Inlow, J; Flurkey, WH. Enzyme, protein, carbohydrate, and phenolic contaminants in commercial tyrosinase preparations; potential problems affecting tyrosinase activity and inhibition studies. J. Agric. Food Chem 2008, 56, 4760–4768. [Google Scholar]
- Flurkey, WH; Ratcliff, B; Lopez, L; Kuglin, J; Dawley, RM. Differentiation of fungal tyrosinase and laccases using selective inhibitors and substrates. In Enzymatic Browning and Its Prevention; Lee, CY, Whitaker, JR, Eds.; American Chemical Society: Washington, DC, USA, 1995; Series 600, Chapter 6, pp. 81–90. [Google Scholar]
- Sugumaran, M; Bolton, JL. Laccase-and not tyrosinase- is the enzyme responsible for quinone methide production from 2,6-dimethoxy-4-allyl-phenol. Arch. Biochem. Biophys 1998, 353, 207–212. [Google Scholar]
- Jacobsohn, GM; Jacobsohn, MK. Incorporation and binding of estrogens into melanin: Comparison of mushroom and mammalian tyrosinases. Biochim. Biophys. Acta 1992, 1116, 173–182. [Google Scholar]
- Galindo, JD; Martinez, JH; Lopez-Ballester, JA; Penafiel, R; Solano, F; Lozano, JA. The effect of polyamines on tyrosinase activity. Biochem. Int 1987, 15, 1151–1158. [Google Scholar]
- Rescigno, A; Sollai, F; Pisu, B; Rinaldi, A; Sanjust, E. Tyrosinase inhibition: General and applied aspects. J. Enzyme. Inhib. Med. Chem 2002, 17, 207–218. [Google Scholar]
- Seo, SY; Sharma, VK; Sharma, N. Mushroom tyrosinase: Recent prospects. J. Agric. Food Chem 2003, 51, 2837–2853. [Google Scholar]
- Kim, YJ; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell. Molec. Life Sci 2005, 62, 1707–1723. [Google Scholar]
- Marrero-Ponce, Y; Khan, MTH; Martin, GMC; Ather, A; Sultankhodzhaev, MN; Torrens, F; Rotondo, R. Prediction of tyrosinase inhibition activity using atom-based bilinear indices. Chem. Med. Chem 2007, 2, 449–478. [Google Scholar]
- Kahn, V; Zakin, V. Effect of salicylhydroxamic acid (SHAM) on dl-DOPA oxidation by mushroom tyrosinae and by NaIO4. J. Food Biochem 2000, 24, 399–415. [Google Scholar]
- Chen, QX; Ke, LN; Song, KK; Huang, H; Liu, XD. Inhibitory effects of hexylresorcinol and dodecylresorcinaol on mushroom (Agaricus bisporus) tyrosinase. Prot. J 2004, 23, 135–141. [Google Scholar]
- Kahn, V; Andrawis, A. Inhibition of mushroom tyrosinase by tropolone. Phytochemistry 1985, 24, 905–908. [Google Scholar]
- Andrawis, A; Kahn, V. Effect of methimazole on the activity of mushroom tyrosinase. Biochem. J 1986, 235, 91–96. [Google Scholar]
- Park, KH; Park, YD; Lee, JR; Hahn, HS; Lee, SJ; Bae, CD; Yang, JM; Kim, DE; Hahn, MJ. Inhibition kinetics of mushroom tyrosinase by copper-chelating ammonium tetrathiomolybdate. Biochim. Biophys. Acta 2005, 1726, 115–120. [Google Scholar]
- Park, YD; Kim, SY; Lyou, YJ; Lee, JY; Yang, JM. A new type of uncompetitive inhibition of tyrosinase by Cl− binding. Biochimie 2005, 87, 931–937. [Google Scholar]
- Masamoto, Y; Ando, H; Murata, Y; Shimoishi, Y; Tada, M; Takahata, K. Mushroom tyrosinase inhibitory activtiy of esculetin isolated from seeds of Euphorbia lathyris, L. Biosci. Biotechnol. Biochem 2003, 67, 631–634. [Google Scholar]
- Kim, YJ; No, JK; Lee, JH; Chung, HY. 4,4′-Dihydroxybiphenyl as a new potent tyrosinase inhibitor. Biol. Pharm. Bull 2005, 28, 323–327. [Google Scholar]
- Wang, Q; Qiu, L; Chen, X; Song, KK; Shi, Y; Chen, QX. Inhibitory effects of phloridzin dehydrate on the activity of mushroom (Agaricus bisporus) tyrosinase. Bioorg. Med. Chem 2007, 15, 1568–1571. [Google Scholar]
- Li, B; Huang, Y; Paskewitz, SM. Hen egg white lysozyme as an inhibitor of mushroom tyrosinase. FEBS Lett 2006, 580, 1877–1882. [Google Scholar]
- Han, HY; Zou, HC; Jeon, JY; Wang, JY; Xu, WA; Yang, JM; Park, YD. The inhibition kinetics and thermodynamic changes of tyrosinase via zinc ion. Biochim. Biophys. Acta 2007, 1774, 822–827. [Google Scholar]
- Ha, TJ; Tamura, S; Kubo, I. Effects of mushroom tyrosinase on anisaldehyde. J. Agric. Food Chem 2005, 53, 7024–7028. [Google Scholar]
- Chen, QX; Liu, XD; Huang, H. Inactivation kinetics of mushroom tyrosinase in the dimethyl sulfoxide solution. Biochemistry (Moscow) 2003, 68, 644–649. [Google Scholar]
- Jimenez, M; Garcis-Carmona, F. 4-Substituted resorcinols (sulfite alternatives) as slow binding inhibitors of tyrosinase catecholase activity. J. Agric. Food Chem 1997, 45, 2061–2065. [Google Scholar]
- Munoz-Munoz, JL; Garcia-Molina, F; Varon, R; Rodriquez-Lopez, JN; Garcia-Canovas, F; Tudela, J. Kinetic chara-cterization of the oxidation of esculetin by polyphenol oxidase and peroxidase. Biosci. Biotechnol. Biochem 2007, 71, 390–396. [Google Scholar]
- Sollai, F; Zucca, P; Sanjust, E; Steri, D; Rescigno, A. Umbelliferone and esculetin: Inhibitors or substrates for polyphenol oxidases? Biol. Pharm. Bull 2008, 31, 2187–2193. [Google Scholar]
- Kasraee, B. Peroxidase-mediated mechanisms are involved in the melanocytotoxic and melanogenesis-inhibiting effects of chemical agents. Dermatology 2002, 205, 329–339. [Google Scholar]
- Passi, S; Nazzaro-Porro, M. Molecular basis of substrate and inhibitory specificity of tyrosinase: Phenolic compounds. Br. J. Dermatol 1981, 104, 659–665. [Google Scholar]
- Lee, HS. Tyrosinase inhibitors of Pulsatila cernua root-derived materials. J. Agric. Food Chem 2002, 50, 1400–1403. [Google Scholar]
- Perez-Gilabert, M; Garcia-Carmona, F. Dimethyl sulfide, a volatile flavor constituent, is a slow binding inhibitor of tyrosinase. Biochem. Biophys. Res. Commun 2001, 285, 257–261. [Google Scholar]
- Arakawa, T; Kita, Y; Timasheff, SN. Protein precipitation and denaturation by dimethyl sulfoxide. Biophys. Chem 2007, 131, 62–70. [Google Scholar]
- Tjernberg, A; Markova, N; Griffiths, WJ; Hallen, D. DMSO-related effects in protein characterization. J. Biomolec. Screen 2006, 11, 131–137. [Google Scholar]
- Fan, Y; Flurkey, WH. Purification and characterization of tyrosinase from gill tissue of Portabella mushrooms. Phytochemistry 2004, 65, 671–678. [Google Scholar]
- Ingebrigtsen, J; Flurkey, WH. Affinity and hydrophobic chromatography of mushroom tyrosinase. Phytochemistry 1988, 27, 1593–1599. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Crude MT μM | Commer. MT μM | Purified MT μM | Reported IC50 Values μM |
|---|---|---|---|---|
| kojic acid | 36 | 47 | 45 | 10–300 [10,18] |
| SHAM | 0.24 | 0.4 | 0.45 | 0.45 [19], 0.8, 1.2 [10] |
| 4-HR | 5.8 | 1.5 | 1.2 | 0.85 [20], 2.5, 18 [10] |
| tropolone | 0.39 | 1.2 | 0.95 | 0.4 [21], 1.3, 1.7 [10] |
| methimazole | 46 | 55 | 50 | 200 [22], 40, 47 [10] |
| ATTM | 3.4 | 3.3 | 2 | 2 [23], 7.5, 28 [10] |
| NaCl | 65 mM | 270 mM | 360 mM | 25 mM [24] |
| esculetin* | 70 | 280 | 685 | 43 [25], 210, 225 [10] |
| biphenol* | 50a | 10b | 67c | 1.9 (tyrosine) [26] |
| phloridzin* | 625 | 1740 | 1400 | 110 [27] |
| hydroquinone | activatesd | activatese | activatesf | 26 [18] |
| lysozyme | >10 mg/mLg | >20 mg/mLh | >20 mg/mLi | 0.32 μM [28] |
| ZnSO4 | none@1 mM | none@1 mM | none@1 mM | 50 [29] |
| anisaldehyde* | 1 mM | 1.4 mM | 1.8 mMj | 160 [30] |
| DMSO | 2 M | 1.75 M | 1.4 M | 2.45 M [31] |
| dimethyl sulfide | NI to 500 μM | NI to 500 μM | NI to 500 μM | |
| dimethyl sulfone | NI to 2 mM | NI to 2 mM | NI to 2 mM | |
| ethanol | 2.3 M | 2.1 M | 1.8 M |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Neeley, E.; Fritch, G.; Fuller, A.; Wolfe, J.; Wright, J.; Flurkey, W. Variations in IC50 Values with Purity of Mushroom Tyrosinase. Int. J. Mol. Sci. 2009, 10, 3811-3823. https://doi.org/10.3390/ijms10093811
Neeley E, Fritch G, Fuller A, Wolfe J, Wright J, Flurkey W. Variations in IC50 Values with Purity of Mushroom Tyrosinase. International Journal of Molecular Sciences. 2009; 10(9):3811-3823. https://doi.org/10.3390/ijms10093811
Chicago/Turabian StyleNeeley, Elizabeth, George Fritch, Autumn Fuller, Jordan Wolfe, Jessica Wright, and William Flurkey. 2009. "Variations in IC50 Values with Purity of Mushroom Tyrosinase" International Journal of Molecular Sciences 10, no. 9: 3811-3823. https://doi.org/10.3390/ijms10093811
APA StyleNeeley, E., Fritch, G., Fuller, A., Wolfe, J., Wright, J., & Flurkey, W. (2009). Variations in IC50 Values with Purity of Mushroom Tyrosinase. International Journal of Molecular Sciences, 10(9), 3811-3823. https://doi.org/10.3390/ijms10093811