
Discovery and Putative Perception Mechanisms of Novel Umami Peptides from Ruditapes philippinarum Cooking Liquid: In Silico Screening, Molecular Docking, Sensory Evaluation, and STC-1 Cell-Based Validation

Abstract
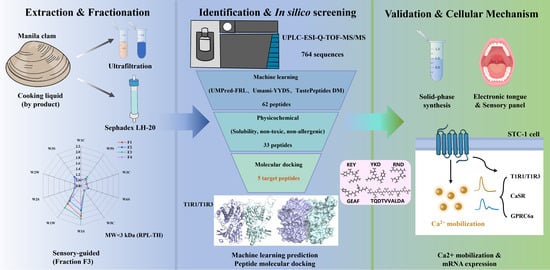
1. Introduction
2. Results and Discussion
2.1. Characterization of Flavor Compounds and Enrichment of Umami Fractions
2.1.1. Profiling of Umami-Related Matrix Components in the Cooking Liquid
2.1.2. Sensory-Guided Fractionation and Enrichment of Umami Peptides
2.2. Sequence Identification and In Silico Screening
2.2.1. Peptidomic Profiling and In Silico Screening
2.2.2. Molecular Docking and Receptor Interactions Analysis
2.3. Taste Evaluation of the Synthesized Peptides
2.3.1. Sensory Evaluation of Target Peptides
2.3.2. Electronic Tongue Analysis and Principal Component Analysis (PCA)
2.4. Effects of Peptides on Ca2+ Release in STC-1 Cells
2.5. Multi-Receptor Perception Mechanisms: Gene Expression Profiling of Taste Receptors
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Preparation and Purification of Umami Peptides
3.3. Sequence Identification and Virtual Screening
3.4. Homology Modeling and Molecular Docking
3.5. Sensory Evaluation and Electronic Tongue Analysis
3.5.1. Sensory Evaluation
3.5.2. Electronic Nose Analysis
3.5.3. Electronic Tongue Analysis
3.6. Cell Culture and Functional Assays
3.6.1. Cell Viability Assay
3.6.2. Intracellular Ca2+ Imaging
3.7. RNA Extraction, Reverse Transcription, and Quantitative PCR
3.7.1. RNA Extraction
3.7.2. Reverse Transcription
3.7.3. Quantitative PCR
3.7.4. Data Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| VFT | Venus flytrap |
| FAA | Free amino acid |
| BMP | Beefy meaty peptide |
| CaSR | calcium-sensing receptor |
| CID | collision-induced dissociation |
| DDA | data-dependent acquisition |
| CCK-8 | Cell counting kit-8 |
| MSG | Monosodium glutamate |
| HBSS | Hank’s balanced salt solution |
| RPL | Ruditapes philippinarum cooking liquid |
| RPL-TT | The fraction (MW > 10 kDa) obtained by ultrafiltration of the umami peptide from the cooking liquor of Ruditapes philippinarum |
| RPL-TE | The fractions (MW 3–10 kDa) obtained by ultrafiltration of the umami peptide from the cooking liquor of Ruditapes philippinarum |
| RPL-TH | The fractions (MW < 3 kDa) obtained by ultrafiltration of the umami peptide from the cooking liquor of Ruditapes philippinarum |
| qPCR | Quantitative real-time polymerase chain reaction |
References
- Sun, C.; Jia, A.; Luan, H.; Bu, Y.; Zhu, W.; Li, X. Exploring of novel salt-enhancing peptides from manila clam: Isolation, peptidomics and molecular characteristics. Food Chem. 2026, 500, 147501. [Google Scholar] [PubMed]
- Jia, A.; Xu, Y.; Sun, C.; Luan, H.; Liu, Z.; Bu, Y.; Zhu, W.; Li, X. Identification and molecular mechanisms of novel antioxidant peptides from Manila clam hydrolysate: Caenorhabditis elegans model, molecular simulations, and quantum chemistry. Food Chem. 2025, 490, 145079. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fu, Z.; Liu, Y.; Song, Z.; Yang, X.; Yu, D.; Wang, Q.; Chi, H.; Zheng, J. A comprehensive and comparative study on the biochemical composition and non-volatile taste compounds of thirteen shellfish species. Foods 2025, 14, 1595. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ji, W.; Jiang, H.; Shi, Y.; He, L.; Gu, Z.; Zhu, S. Comparison of biochemical composition and non-volatile taste active compounds in raw, high hydrostatic pressure-treated and steamed oysters Crassostrea hongkongensis. Food Chem. 2021, 344, 128632. [Google Scholar] [PubMed]
- Hao, L.; Fan, Y.; Tian, Q.; Fan, C.; Zhang, Y.; Dong, P.; Wang, Y.; Hou, H. Current progress in umami peptides from blue foods: Preparation, screening, evaluation, synergistic effects, and taste mechanism. Trends Food Sci. Technol. 2024, 149, 104567. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Sheng, Y.; Chi, C.; Wang, B. A systematic review on marine umami peptides: Biological sources, preparation methods, structure-umami relationship, mechanism of action and biological activities. Food Biosci. 2024, 57, 103637. [Google Scholar] [CrossRef]
- Liang, Q.; Zhong, Y.; Ren, X.; Liang, Z.; Zhu, C.; Wang, L.; Mou, H. Mining for novel umami peptides from sea cucumber viscera hydrolysate with ACE inhibitory activity. J. Agric. Food Chem. 2025, 73, 15751–15766. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shi, Y.; Qiu, Z.; Zhang, J.; Liu, T.; Shi, W.; Wang, X. Food-derived umami peptides: Bioactive ingredients for enhancing flavor. Crit. Rev. Food Sci. Nutr. 2025, 65, 5970–5986. [Google Scholar]
- Xu, C.; Tang, H.; Wang, X. Systematic review on functional umami peptides: Physiological perceptions, bioinformatics and peptidomics approaches, developments and applications. Trends Food Sci. Technol. 2025, 162, 105068. [Google Scholar] [CrossRef]
- Zhao, J.; Li, X.; Huang, L.; Lin, H.; He, Q. Unlocking the potential of umami peptides: A comprehensive review of preparation methods, evaluation strategies, health benefits, and taste transduction mechanisms. Food Res. Int. 2025, 225, 118024. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Ueji, K.; Mizuta, H.; Inui-Yamamoto, C.; Kumamoto, N.; Shibata, Y.; Ugawa, S. GPRC6A as a novel kokumi receptor responsible for enhanced taste preferences by ornithine. Elife 2025, 13, RP101629. [Google Scholar] [CrossRef]
- Yang, J.; Liao, J.; Dong, H.; Huang, G.; Bai, W.; Tu, J.; Zeng, X. Synergistic effect of kokumi-active γ-glutamyl peptides and l-glutamate on enhancing umami sensation and stimulating cholecystokinin secretion via T1R1/T1R3 activation in STC-1 cells. J. Agric. Food Chem. 2022, 70, 14395–14402. [Google Scholar] [CrossRef] [PubMed]
- Santos-Hernández, M.; Vivanco-Maroto, S.M.; Miralles, B.; Recio, I. Food peptides as inducers of CCK and GLP-1 secretion and GPCRs involved in enteroendocrine cell signalling. Food Chem. 2023, 402, 134225. [Google Scholar] [PubMed]
- Zhang, W.; Guan, H.; Wang, M.; Wang, W.; Pu, J.; Zou, H.; Li, D. Exploring the relationship between small peptides and the T1R1/T1R3 umami taste receptor for umami peptide prediction: A combined approach. J. Agric. Food Chem. 2024, 72, 13262–13272. [Google Scholar] [PubMed]
- Geng, H.; Xu, C.; Ma, H.; Dai, Y.; Jiang, Z.; Yang, M.; Zhu, D. In silico discovery and sensory validation of umami peptides in fermented sausages: A study integrating deep learning and molecular modeling. Foods 2025, 14, 2422. [Google Scholar] [CrossRef] [PubMed]
- An, F.; Wu, J.; Xu, X.; Du, M.; Ji, S.; Guo, J.; Pan, G.; Jiang, J.; Cao, K.; Tao, D. An integrated in silico and multi-omics workflow identifies novel microbiota-derived umami peptides and elucidates T1R1/T1R3 binding mechanisms. Food Chem. 2026, 517, 149468. [Google Scholar] [PubMed]
- Gu, Y.; Zhou, X.; Niu, Y.; Zhang, J.; Sun, B.; Liu, Z.; Mao, X.; Zhang, Y.; Li, K.; Zhang, Y. Screening and identification of novel umami peptides from yeast proteins: Insights into their mechanism of action on receptors T1R1/T1R3. Food Chem. 2025, 463, 141138. [Google Scholar] [PubMed]
- Zhao, J.; Liao, S.; Han, J.; Xie, Y.; Tang, J.; Zhao, J.; Shao, W.; Wang, Q.; Lin, H. Revealing the secret of umami taste of peptides derived from fermented broad bean paste. J. Agric. Food Chem. 2023, 71, 4706–4716. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, X.; Zheng, J.; Sun, Y.; Dong, X.; Zhou, D.; Zhu, B.; Qin, L. Comprehensive metabolomic and lipidomic profiling of the seasonal variation of blue mussels (Mytilus edulis L.): Free amino acids, 5′-nucleotides, and lipids. Lwt 2021, 149, 111835. [Google Scholar] [CrossRef]
- Zhu, S.; Zhu, L.; Ke, Z.; Chen, H.; Zheng, Y.; Yang, P.; Xiang, X.; Zhou, X.; Jin, Y.; Deng, S. A comparative study on the taste quality of Mytilus coruscus under different shucking treatments. Food Chem. 2023, 412, 135480. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Regenstein, J.M.; Luo, Y. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [PubMed]
- Yan, F.; Cui, H.; Zhang, Q.; Hayat, K.; Yu, J.; Hussain, S.; Tahir, M.U.; Zhang, X.; Ho, C.-T. Small peptides hydrolyzed from pea protein and their Maillard reaction products as taste modifiers: Saltiness, umami, and kokumi enhancement. Food Bioprocess Technol. 2021, 14, 1132–1141. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, J.; Hansen, E.T.; Bredie, W.L.; Lametsch, R. Structural characteristics of low bitter and high umami protein hydrolysates prepared from bovine muscle and porcine plasma. Food Chem. 2018, 257, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hua, Y.; Pan, D.; Qi, L.; Xiao, C.; Xiong, Y.; Lu, W.; Dang, Y.; Gao, X.; Zhao, Y. A rapid selection strategy for umami peptide screening based on machine learning and molecular docking. Food Chem. 2023, 404, 134562. [Google Scholar] [PubMed]
- Hu, K.; Sun, G.; Yu, W.; Zhang, M.; Wang, S.; Cao, Y.; Hu, D.; Liang, L.; He, G.; Hu, J. Molecular recognition and modification strategies of umami dipeptides with T1R1/T1R3 receptors. Molecules 2025, 30, 2774. [Google Scholar] [PubMed]
- Meng, H.; Cui, Z.; Yu, Y.; Li, Y.; Jiang, S.; Liu, Y. From molecular dynamics to taste sensory perception: A comprehensive study on the interaction of umami peptides with the T1R1/T1R3-VFT receptor. J. Agric. Food Chem. 2024, 72, 6533–6543. [Google Scholar] [PubMed]
- An, F.; Cao, K.; Ji, S.; Wang, Y.; Pan, G.; Ma, Y.; Zhao, Y.; Wu, J.; Wu, R. Identification, taste characterization, and molecular docking study of a novel microbiota-derived umami peptide. Food Chem. 2023, 404, 134583. [Google Scholar] [PubMed]
- Cui, Z.; Meng, H.; Zhou, T.; Yu, Y.; Gu, J.; Zhang, Z.; Zhu, Y.; Zhang, Y.; Liu, Y.; Wang, W. Noteworthy consensus effects of D/E residues in umami peptides used for designing the novel umami peptides. J. Agric. Food Chem. 2024, 72, 2789–2800. [Google Scholar] [CrossRef] [PubMed]
- Zazeri, G.; Povinelli, A.P.R.; Le Duff, C.S.; Tang, B.; Cornelio, M.L.; Jones, A.M. Synthesis and spectroscopic analysis of piperine-and piperlongumine-inspired natural product scaffolds and their molecular docking with IL-1β and NF-κB proteins. Molecules 2020, 25, 2841. [Google Scholar] [PubMed]
- Zazeri, G.; Povinelli, A.P.R.; Pavan, N.M.; Jones, A.M.; Ximenes, V.F. Solvent-induced lag phase during the formation of lysozyme amyloid fibrils triggered by sodium dodecyl sulfate: Biophysical experimental and in silico study of solvent effects. Molecules 2023, 28, 6891. [Google Scholar] [CrossRef] [PubMed]
- Jünger, M.; Mittermeier-Kleßinger, V.K.; Farrenkopf, A.; Dunkel, A.; Stark, T.; Fröhlich, S.; Somoza, V.; Dawid, C.; Hofmann, T. Sensoproteomic discovery of taste-modulating peptides and taste re-engineering of soy sauce. J. Agric. Food Chem. 2022, 70, 6503–6518. [Google Scholar] [PubMed]
- Zhang, J.; Zhao, M.; Su, G.; Lin, L. Identification and taste characteristics of novel umami and umami-enhancing peptides separated from peanut protein isolate hydrolysate by consecutive chromatography and UPLC–ESI–QTOF–MS/MS. Food Chem. 2019, 278, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.; Zuker, C.S. An amino-acid taste receptor. Nature 2002, 416, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, K.; Zhu, W. Amino acid sensing in the gut and its mediation in gut-brain signal transduction. Anim. Nutr. 2016, 2, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Brennan, S.C.; Davies, T.S.; Schepelmann, M.; Riccardi, D. Emerging roles of the extracellular calcium-sensing receptor in nutrient sensing: Control of taste modulation and intestinal hormone secretion. Brit. J. Nutr. 2014, 111, S16–S22. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Verfaillie, D.; Janssen, F.; Royen, G.V.; Wouters, A.G.B. A systematic study of the impact of the isoelectric precipitation process on the physical properties and protein composition of soy protein isolates. Food Res. Int. 2023, 163, 112177. [Google Scholar] [PubMed]
- Badjona, A.; Bradshaw, R.; Millman, C.; Howarth, M.; Dubey, B. Response surface methodology guided approach for optimization of protein isolate from Faba bean. Part 1/2. Ultrason. Sonochem. 2024, 109, 107012. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M.; Sieniawski, K.; Starowicz, P. BIOPEP database of sensory peptides and amino acids. Food Res. Int. 2016, 85, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Hameduh, T.; Haddad, Y.; Adam, V.; Heger, Z. Homology modeling in the time of collective and artificial intelligence. Comput. Struct. Biotec. 2020, 18, 3494–3506. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Limon, A.; Aguilar-Toalá, J.E.; Liceaga, A.M. Integration of molecular docking analysis and molecular dynamics simulations for studying food proteins and bioactive peptides. J. Agric. Food Chem. 2022, 70, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Feng, Y.; Thomas-Danguin, T.; Zhao, M. Enhancement of saltiness perception by odorants selected from Chinese soy sauce: A gas chromatography/olfactometry-associated taste study. Food Chem. 2021, 335, 127664. [Google Scholar] [PubMed]
- Zhang, N.; Ayed, C.; Wang, W.; Liu, Y. Sensory-guided analysis of key taste-active compounds in pufferfish (Takifugu obscurus). J. Agric. Food Chem. 2019, 67, 13809–13816. [Google Scholar] [PubMed]
- Han, J.; Liang, J.; Yu, X.; Chen, L.; Feng, Y.; Yang, Z.; Mukhtar, A.; Abugri, A.; Zhou, C. Characterization of flavor profiles in chive leaves and stems at different moisture transfer points during combined drying: An integrated approach with e-nose, GC-IMS, GC–MS, and machine learning. Food Chem. 2026, 509, 148496. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAAs | Taste Properties | Taste Threshold (g/100 g) | TAV | FAA Content (g/100 g) |
|---|---|---|---|---|
| Asp | Umami (+) | 0.10 | 4.91 | 0.49 ± 0.10 |
| Thr | Sweetness (+) | 0.26 | 0.32 | 0.08 ± 0.02 |
| Ser | Sweetness (+) | 0.15 | 0.43 | 0.06 ± 0.01 |
| Glu | Umami (+) | 0.03 | 36.21 | 1.09 ± 0.22 |
| Gly | Sweetness (+) | 0.13 | 20.86 | 2.71 ± 0.50 |
| Ala | Sweetness (+) | 0.06 | 14.17 | 0.85 ± 0.18 |
| Cys | Bitterness/Sweetness/Astringency (−) | - | - | 0.02 ± 0.00 |
| Val | Sweetness/Bitterness (−) | 0.04 | 1.72 | 0.07 ± 0.01 |
| Met | Bitterness/Sweetness/Astringency (−) | 0.03 | 1.26 | 0.04 ± 0.01 |
| Ile | Bitterness (−) | 0.09 | 0.63 | 0.06 ± 0.01 |
| Leu | Bitterness (−) | 0.38 | 0.19 | 0.07 ± 0.02 |
| Tyr | Bitterness (−) | - | - | 0.03 ± 0.00 |
| Phe | Bitterness (−) | 0.09 | 0.70 | 0.06 ± 0.00 |
| Lys | Sweetness/Bitterness (−) | 0.05 | 2.79 | 0.14 ± 0.02 |
| His | Bitterness (−) | 0.02 | 7.80 | 0.16 ± 0.18 |
| Arg | Sweetness/Bitterness (+) | 0.05 | 32.68 | 1.63 ± 1.75 |
| Umami amino acids | 1.58 ± 0.33 | |||
| Sweet amino acids | 2.80 ± 0.52 | |||
| Total FAA content | 7.56 ± 0.86 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Wang, R.; Sun, Q.; Zhang, S.; Wang, H.; Xu, T.; Zhao, Q.; Li, Z. Discovery and Putative Perception Mechanisms of Novel Umami Peptides from Ruditapes philippinarum Cooking Liquid: In Silico Screening, Molecular Docking, Sensory Evaluation, and STC-1 Cell-Based Validation. Molecules 2026, 31, 2193. https://doi.org/10.3390/molecules31122193
Wang R, Sun Q, Zhang S, Wang H, Xu T, Zhao Q, Li Z. Discovery and Putative Perception Mechanisms of Novel Umami Peptides from Ruditapes philippinarum Cooking Liquid: In Silico Screening, Molecular Docking, Sensory Evaluation, and STC-1 Cell-Based Validation. Molecules. 2026; 31(12):2193. https://doi.org/10.3390/molecules31122193
Chicago/Turabian StyleWang, Ruiying, Qi Sun, Siyu Zhang, Haibo Wang, Tanye Xu, Qiancheng Zhao, and Zhibo Li. 2026. "Discovery and Putative Perception Mechanisms of Novel Umami Peptides from Ruditapes philippinarum Cooking Liquid: In Silico Screening, Molecular Docking, Sensory Evaluation, and STC-1 Cell-Based Validation" Molecules 31, no. 12: 2193. https://doi.org/10.3390/molecules31122193
APA StyleWang, R., Sun, Q., Zhang, S., Wang, H., Xu, T., Zhao, Q., & Li, Z. (2026). Discovery and Putative Perception Mechanisms of Novel Umami Peptides from Ruditapes philippinarum Cooking Liquid: In Silico Screening, Molecular Docking, Sensory Evaluation, and STC-1 Cell-Based Validation. Molecules, 31(12), 2193. https://doi.org/10.3390/molecules31122193