
Metabolite-Centered Evaluation of Plant-Based Substrates: Integrated Profiling of Short-Chain Fatty Acids (SCFAs) and Neuroactive Compounds with Potential Relevance to the Gut–Brain Axis


Abstract
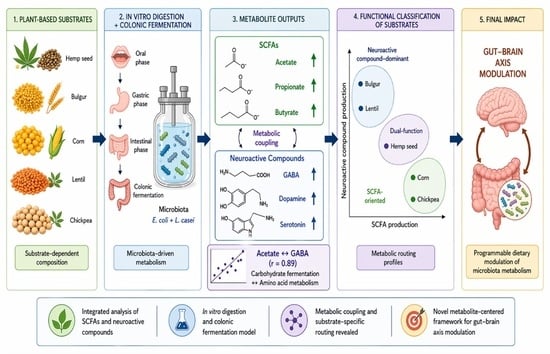
1. Introduction
2. Results
2.1. Short—Chain Fatty Acid Production Composition of Plant-Based Substrates
2.2. Neuroactive Compound Profiles During In Vitro Fermentation
2.3. Comparative Analysis of SCFAs and Neuroactive Compounds
2.4. Correlation Analysis Between SCFAs and Neurotransmitters
3. Discussion
3.1. Substrate-Dependent SCFA Production
3.2. Neuroactive Compound Profiles
3.3. Correlation Analysis and Fermentation-Associated Metabolic Profile Interpretation
3.4. Neuroactive Potential Mapping Interpretation
3.5. Metabolite-Centered Interpretation
3.6. Methodological Considerations and Limitations
4. Materials and Methods
4.1. Samples
4.2. Chemical Reagents and Enzymes
4.3. Sample Preparation
4.4. Oral Phase
4.5. Gastric Phase
4.6. Intestinal Phase
4.7. In Vitro Model Design
4.8. In Vitro Colonic Fermentation and HPLC Analysis of SCFAs and Neuroactive Compounds
4.9. Statistical Analysis
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martino, C.; Dilmore, A.H.; Burcham, Z.M.; Metcalf, J.L.; Jeste, D.; Knight, R. Microbiota succession throughout life from the cradle to the grave. Nat. Rev. Microbiol. 2022, 20, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut–brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Fu, J.; Zheng, Y.; Gao, Y.; Xu, W. Dietary fiber intake and gut microbiota in human health. Microorganisms 2022, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-León, M.J.; Mangalam, A.K.; Regaldiz, A.; González-Madrid, E.; Rangel-Ramírez, M.A.; Álvarez-Mardonez, O.; Vallejos, O.P.; Méndez, C.; Bueno, S.M.; Melo-González, F.; et al. Gut microbiota short-chain fatty acids and their impact on host metabolism and endocrine health. Front. Endocrinol. 2023, 14, 1192216. [Google Scholar] [CrossRef] [PubMed]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Del Chierico, F. Short-chain fatty acids and human health. Life 2024, 14, 559. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Adebo, J.A.; Njobeh, P.B.; Adebiyi, J.A.; Kayitesi, E. Fermentation of cereals and legumes: Impact on nutritional constituents and nutrient bioavailability. Fermentation 2022, 8, 63. [Google Scholar] [CrossRef]
- Yıldız, M. A potential product for sustainable food systems and nutrition: Physicochemical characterization of registered native narlı and vezir hemp seeds. Gıda 2025, 50, 946–960. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Vlckova, K.; Marx, W.; Schellekens, H.; Stanton, C.; Clarke, G.; Jacka, F.N.; Dinan, T.G.; Cryan, J.F. Diet and the microbiota–gut–brain axis: Sowing the Seeds of Good Mental Health. Adv. Nutr. 2021, 12, 1237–1260. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr. Nutr. Rep. 2022, 11, 232–247. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota–gut–brain axis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 625–639. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. Gamma-aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Raes, J.; Letunic, I.; Yamada, T.; Jensen, L.J.; Bork, P. Toward molecular trait-based ecology through integration of biogeochemical, geographical and metagenomic data. Mol. Syst. Biol. 2011, 7, 473. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Lyte, M. Microbial endocrinology in the microbiome–gut–brain axis. Adv. Exp. Med. Biol. 2013, 817, 3–24. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain–gut–microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Agustí, A.; García-Pardo, M.P.; López-Almela, I.; Campillo, I.; Maes, M.; Romaní-Pérez, M.; Sanz, Y.; Campoy, C. Interplay between the gut-brain axis, obesity and cognitive function. Front. Neurosci. 2018, 12, 155. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, S.Y.; Chung, M.S.; Hur, S.J. Development of novel in vitro human digestion systems for screening the bioavailability and digestibility of foods. J. Funct. Foods 2016, 22, 113–121. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- De Baere, S.; Eeckhaut, V.; Steppe, M.; De Maesschalck, C.; De Backer, P.; Van Immerseel, F.; Croubels, S. Development of a HPLC–UV method for the quantitative determination of short-chain fatty acids and lactic acid produced by intestinal bacteria during in vitro fermentation. J. Pharm. Biomed. Anal. 2013, 80, 107–115. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Acetate | Propionate | Butyrate |
|---|---|---|---|
| Chickpea | 2.55 ± 0.05 c | 2.30 ± 0.10 c | 0.252 ± 0.008 a |
| Bulgur | 1.93 ± 0.13 d | 2.45 ± 0.05 c | 0.272 ± 0.013 a |
| Lentil | 2.80 ± 0.05 c | 4.83 ± 0.08 a | 0.188 ± 0.008 c |
| Corn | 2.45 ± 0.05 c | 4.52 ± 0.14 b | 0.168 ± 0.008 c |
| Hemp seed | 4.67 ± 0.16 a | 1.77 ± 0.08 d | 0.227 ± 0.015 b |
| Terebinth | 3.50 ± 0.30 b | 0.98 ± 0.08 e | 0.136 ± 0.006 d |
| Sample | GABA (µg/g) | Dopamine (µg/g) | Serotonin (µg/g) |
|---|---|---|---|
| Chickpea | 29.23 ± 0.87 e | 12.30 ± 0.25 f | 11.33 ± 0.86 c |
| Bulgur | 22.98 ± 0.56 f | 82.90 ± 2.58 a | 21.37 ± 1.06 a |
| Lentil | 47.80 ± 0.61 b | 41.97 ± 1.51 c | 10.78 ± 0.63 c |
| Corn flour | 36.40 ± 0.86 c | 24.93 ± 0.74 d | 8.37 ± 0.25 d |
| Hemp flour | 114.00 ± 0.87 a | 15.97 ± 0.49 e | 8.78 ± 0.18 d |
| Terebinth flour | 35.10 ± 0.56 d | 47.30 ± 1.07 b | 16.68 ± 0.28 b |
| GABA | Dopamine | Serotonin | |
|---|---|---|---|
| Acetate | 0.89 | −0.49 | −0.42 |
| Propionate | −0.18 | −0.06 | −0.43 |
| Butyrate | 0.03 | 0.16 | 0.26 |
| Substrate | Functional Properties Observed |
|---|---|
| Hemp flour | Highest GABA production → potential stress-reducing and neuroprotective effects [9,16] |
| Bulgur | Highest dopamine and serotonin levels → support for cognitive function and mood regulation [22,32] |
| Lentil | Balanced neurotransmitter production → overall support for the gut–brain axis [13,14] |
| Terebinth flour | Elevated dopamine and serotonin → high potential for neuroactive metabolite production [28,29,33] |
| Corn flour | Moderate GABA and dopamine levels → supportive functional effects [14,15] |
| Chickpea | Moderate serotonin and GABA levels → supportive role in gut health [24,32] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Yıldız, M. Metabolite-Centered Evaluation of Plant-Based Substrates: Integrated Profiling of Short-Chain Fatty Acids (SCFAs) and Neuroactive Compounds with Potential Relevance to the Gut–Brain Axis. Molecules 2026, 31, 2073. https://doi.org/10.3390/molecules31122073
Yıldız M. Metabolite-Centered Evaluation of Plant-Based Substrates: Integrated Profiling of Short-Chain Fatty Acids (SCFAs) and Neuroactive Compounds with Potential Relevance to the Gut–Brain Axis. Molecules. 2026; 31(12):2073. https://doi.org/10.3390/molecules31122073
Chicago/Turabian StyleYıldız, Mustafa. 2026. "Metabolite-Centered Evaluation of Plant-Based Substrates: Integrated Profiling of Short-Chain Fatty Acids (SCFAs) and Neuroactive Compounds with Potential Relevance to the Gut–Brain Axis" Molecules 31, no. 12: 2073. https://doi.org/10.3390/molecules31122073
APA StyleYıldız, M. (2026). Metabolite-Centered Evaluation of Plant-Based Substrates: Integrated Profiling of Short-Chain Fatty Acids (SCFAs) and Neuroactive Compounds with Potential Relevance to the Gut–Brain Axis. Molecules, 31(12), 2073. https://doi.org/10.3390/molecules31122073