
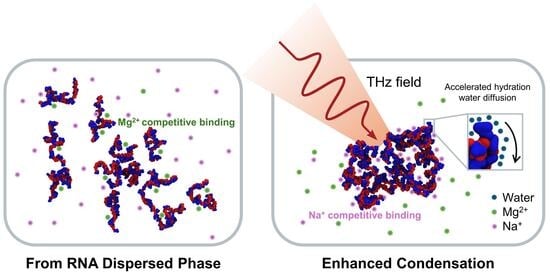
Enhanced Condensation of RNA Repeats Induced by Terahertz Oscillatory Fields

 , , , and
, , , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Effects of THz Oscillatory Fields on the Macroscopic Parameters
2.2. Nucleobase–Nucleobase (Nb–Nb) Interactions
2.3. RNA–Cation Interactions
2.4. Effects of THz Oscillatory Fields on the Hydration Layer
3. Methods
3.1. Model Validation
3.2. CGMD Simulation Setup
3.3. Simulation Procedure Parameters
3.4. Analysis Methods
3.4.1. Trajectory Quality Control and Exclusion of Anomalous Data
3.4.2. Extended Analysis of THz Modulation
3.4.3. MD Analysis of RNA–Cation Contacts
3.4.4. Hydration Water Analysis and Diffusion Measurements
3.4.5. Statistics and Reproducibility
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyman, A.A.; Weber, C.A.; Jülicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular Condensates: Organizers of Cellular Biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Bisaria, N.; Greenfeld, M.; Limouse, C.; Mabuchi, H.; Herschlag, D. Quantitative Tests of a Reconstitution Model for RNA Folding Thermodynamics and Kinetics. Proc. Natl. Acad. Sci. USA 2017, 114, E7688–E7696. [Google Scholar] [CrossRef]
- Guo, Q.; Shi, X.; Wang, X. RNA and Liquid-Liquid Phase Separation. Non-Coding RNA Res. 2021, 6, 92–99. [Google Scholar] [CrossRef]
- Bevilacqua, P.C.; Williams, A.M.; Chou, H.-L.; Assmann, S.M. RNA Multimerization as an Organizing Force for Liquid–Liquid Phase Separation. RNA 2022, 28, 16–26. [Google Scholar] [CrossRef]
- Jain, A.; Vale, R.D. RNA Phase Transitions in Repeat Expansion Disorders. Nature 2017, 546, 243–247. [Google Scholar] [CrossRef]
- Grese, Z.R.; Bastos, A.C.; Mamede, L.D.; French, R.L.; Miller, T.M.; Ayala, Y.M. Specific RNA Interactions Promote TDP-43 Multivalent Phase Separation and Maintain Liquid Properties. EMBO Rep. 2021, 22, e53632. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Duncan, S.; Yang, X.; Abdelhamid, M.A.S.; Huang, L.; Zhang, H.; Benfey, P.N.; Waller, Z.A.E.; Ding, Y. G-Quadruplex Structures Trigger RNA Phase Separation. Nucleic Acids Res. 2019, 47, 11746–11754. [Google Scholar] [CrossRef]
- Ramachandran, V.; Potoyan, D. Molecular Drivers of RNA Phase Separation. Proc. Natl. Acad. Sci. USA 2025, 122, e2511348122. [Google Scholar] [CrossRef]
- Preto, J.; Pettini, M.; Tuszynski, J.A. Possible Role of Electrodynamic Interactions in Long-Distance Biomolecular Recognition. Phys. Rev. E 2015, 91, 052710. [Google Scholar] [CrossRef] [PubMed]
- Pezzotti, S.; König, B.; Ramos, S.; Schwaab, G.; Havenith, M. Liquid–Liquid Phase Separation? Ask the Water! J. Phys. Chem. Lett. 2023, 14, 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Fay, M.M.; Anderson, P.J.; Ivanov, P. ALS/FTD-Associated C9ORF72 Repeat RNA Promotes Phase Transitions In Vitro and in Cells. Cell Rep. 2017, 21, 3573–3584. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Berry, J.; Pannucci, N.; Haataja, M.P.; Toettcher, J.E.; Brangwynne, C.P. Spatiotemporal Control of Intracellular Phase Transitions Using Light-Activated optoDroplets. Cell 2017, 168, 159–171.e14. [Google Scholar] [CrossRef]
- Lee, M.; Moon, H.C.; Jeong, H.; Kim, D.W.; Park, H.Y.; Shin, Y. Optogenetic Control of mRNA Condensation Reveals an Intimate Link between Condensate Material Properties and Functions. Nat. Commun. 2024, 15, 3216. [Google Scholar] [CrossRef]
- Nakamura, H.; Lee, A.A.; Afshar, A.S.; Watanabe, S.; Rho, E.; Razavi, S.; Suarez, A.; Lin, Y.-C.; Tanigawa, M.; Huang, B.; et al. Intracellular Production of Hydrogels and Synthetic RNA Granules by Multivalent Molecular Interactions. Nat. Mater. 2018, 17, 79–89. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Yoshii, T.; Ikuta, M.; Tsukiji, S. Synthetic Protein Condensates That Inducibly Recruit and Release Protein Activity in Living Cells. J. Am. Chem. Soc. 2021, 143, 6434–6446. [Google Scholar] [CrossRef]
- Peng, X.-Y.; Zhou, H. Biological effects of terahertz waves. Acta Phys. Sin. 2021, 70, 240701–240714. [Google Scholar] [CrossRef]
- Wang, P.; Lou, J.; Fang, G.; Chang, C. Progress on Cutting-Edge Infrared-Terahertz Biophysics. IEEE Trans. Microw. Theory Tech. 2022, 70, 5117–5140. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, L.; Wang, K.; Guo, L.; Ning, H.; Wang, S.; Gong, Y. Terahertz Waves Regulate the Mechanical Unfolding of Tau Pre-mRNA Hairpins. iScience 2023, 26, 107572. [Google Scholar] [CrossRef]
- Ahlers, J.; Adams, E.M.; Bader, V.; Pezzotti, S.; Winklhofer, K.F.; Tatzelt, J.; Havenith, M. The Key Role of Solvent in Condensation: Mapping Water in Liquid-Liquid Phase-Separated FUS. Biophys. J. 2021, 120, 1266–1275. [Google Scholar] [CrossRef]
- Hattori, T.; Huang, C.; Ahmed, F.; Nomoto, A.; Shiraki, K. Terahertz Spectroscopic Study of Liquid-Liquid Phase Separation of Protein Solutions 2024. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=5031950 (accessed on 24 November 2024).
- Valdes-Garcia, G.; Heo, L.; Lapidus, L.J.; Feig, M. Modeling Concentration-Dependent Phase Separation Processes Involving Peptides and RNA via Residue-Based Coarse-Graining. J. Chem. Theory Comput. 2023, 19, 669–678. [Google Scholar] [CrossRef]
- Jussupow, A.; Bartley, D.; Lapidus, L.J.; Feig, M. COCOMO2: A Coarse-Grained Model for Interacting Folded and Disordered Proteins. J. Chem. Theory Comput. 2025, 21, 2095–2107. [Google Scholar] [CrossRef] [PubMed]
- Laghmach, R.; Malhotra, I.; Potoyan, D.A. Multiscale Modeling of Protein-RNA Condensation in and Out of Equilibrium. Methods Mol. Biol. Clifton NJ 2023, 2563, 117–133. [Google Scholar] [CrossRef]
- Uusitalo, J.J.; Ingólfsson, H.I.; Marrink, S.J.; Faustino, I. Martini Coarse-Grained Force Field: Extension to RNA. Biophys. J. 2017, 113, 246–256. [Google Scholar] [CrossRef]
- Tsanai, M.; Frederix, P.W.J.M.; Schroer, C.F.E.; Souza, P.C.T.; Marrink, S.J. Coacervate Formation Studied by Explicit Solvent Coarse-Grain Molecular Dynamics with the Martini Model. Chem. Sci. 2021, 12, 8521–8530. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Wan, Z.; Ngai, T.; Steve Tse, Y.-L. Capturing Coacervate Formation and Protein Partition by Molecular Dynamics Simulation. Chem. Sci. 2023, 14, 1168–1175. [Google Scholar] [CrossRef]
- Zhang, Q.; Valério, M.; Grünewald, L.; Borges-Araújo, L.; Grünewald, F.; Wang, S.; Marrink, S.J.; Gong, Y.; Souza, P.C.T. Coarse-Grained Simulations Reveal Salt- and Length-Dependent Condensation of G4C2 RNA Repeats. Press. J. Phys. Chem. Lett. 2026. [Google Scholar] [CrossRef]
- Souza, P.C.T.; Alessandri, R.; Barnoud, J.; Thallmair, S.; Faustino, I.; Grünewald, F.; Patmanidis, I.; Abdizadeh, H.; Bruininks, B.M.H.; Wassenaar, T.A.; et al. Martini 3: A General Purpose Force Field for Coarse-Grained Molecular Dynamics. Nat. Methods 2021, 18, 382–388. [Google Scholar] [CrossRef]
- Wang, M.; Falke, S.; Schubert, R.; Lorenzen, K.; Cheng, Q.; Exner, C.; Brognaro, H.; Nzanzu Mudogo, C.; Betzel, C. Pulsed Electric Fields Induce Modulation of Protein Liquid–Liquid Phase Separation. Soft Matter 2020, 16, 8547–8553. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, N.; Chen, X.; Wu, Z.; Zhong, W.; Yu, D.; Zhang, H. Effects of Moderate Electric Field on the Structural Properties and Aggregation Characteristics of Soybean Protein Isolate. Food Hydrocoll. 2022, 133, 107911. [Google Scholar] [CrossRef]
- Axelrod, R.; Beyrer, M.; Mathys, A. Impact of the Electric Field Intensity and Treatment Time on Whey Protein Aggregate Formation. J. Dairy Sci. 2022, 105, 6589–6600. [Google Scholar] [CrossRef]
- Hu, X.; Wang, H.; Hu, Y.; Tu, Z. Insight into the Effects of Pulsed Electric Field on the Structure, Aggregation Characteristics and Functional Properties of Whey Proteins. Food Hydrocoll. 2024, 154, 110111. [Google Scholar] [CrossRef]
- Agrawal, A.; Douglas, J.F.; Tirrell, M.; Karim, A. Manipulation of Coacervate Droplets with an Electric Field. Proc. Natl. Acad. Sci. USA 2022, 119, e2203483119. [Google Scholar] [CrossRef]
- Lin, C.-J.; Wang, J.-J.; Jiang, Y.; Chen, S.-L.; Li, H.-F.; Zhao, W.-H.; Huang, Q.-R.; Rong, C.-R.; Duan, X.-Z. Structural Properties of Ions and Polyelectrolytes in Aqueous Solutions under External Electric Fields: The Sign Effect. Chin. J. Polym. Sci. 2024, 42, 1341–1352. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.; Souza, P.C.T.; Garcia, W.; Batista, F.A.H.; Portugal, R.V.; Nascimento, A.S.; Nakahira, M.; Lima, L.M.T.R.; Polikarpov, I.; Skaf, M.S. On the Denaturation Mechanisms of the Ligand Binding Domain of Thyroid Hormone Receptors. J. Phys. Chem. B 2010, 114, 1529–1540. [Google Scholar] [CrossRef]
- Petrov, A.S.; Bowman, J.C.; Harvey, S.C.; Williams, L.D. Bidentate RNA–Magnesium Clamps: On the Origin of the Special Role of Magnesium in RNA Folding. RNA 2011, 17, 291–297. [Google Scholar] [CrossRef]
- Onuchic, P.L.; Milin, A.N.; Alshareedah, I.; Deniz, A.A.; Banerjee, P.R. Divalent Cations Can Control a Switch-like Behavior in Heterotypic and Homotypic RNA Coacervates. Sci. Rep. 2019, 9, 12161. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, J. Driving Forces of RNA Condensation Revealed through Coarse-Grained Modeling with Explicit Mg2+. bioRxiv 2025. [Google Scholar] [CrossRef]
- Boeynaems, S.; Holehouse, A.S.; Weinhardt, V.; Kovacs, D.; Van Lindt, J.; Larabell, C.; Van Den Bosch, L.; Das, R.; Tompa, P.S.; Pappu, R.V.; et al. Spontaneous Driving Forces Give Rise to protein−RNA Condensates with Coexisting Phases and Complex Material Properties. Proc. Natl. Acad. Sci. USA 2019, 116, 7889–7898. [Google Scholar] [CrossRef]
- Poudyal, R.R.; Sieg, J.P.; Portz, B.; Keating, C.D.; Bevilacqua, P.C. RNA Sequence and Structure Control Assembly and Function of RNA Condensates. RNA 2021, 27, 1589–1601. [Google Scholar] [CrossRef]
- Alessandri, R.; Barnoud, J.; Gertsen, A.S.; Patmanidis, I.; de Vries, A.H.; Souza, P.C.T.; Marrink, S.J. Martini 3 Coarse-Grained Force Field: Small Molecules. Adv. Theory Simul. 2022, 5, 2100391. [Google Scholar] [CrossRef]
- Grünewald, F.; Punt, M.H.; Jefferys, E.E.; Vainikka, P.A.; König, M.; Virtanen, V.; Meyer, T.A.; Pezeshkian, W.; Gormley, A.J.; Karonen, M.; et al. Martini 3 Coarse-Grained Force Field for Carbohydrates. J. Chem. Theory Comput. 2022, 18, 7555–7569. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling through Velocity Rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef]
- Tang, J.; Yin, H.; Ma, J.; Bo, W.; Yang, Y.; Xu, J.; Liu, Y.; Gong, Y. Terahertz Electric Field-Induced Membrane Electroporation by Molecular Dynamics Simulations. J. Membr. Biol. 2018, 251, 681–693. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, T.; Ru, Y.; Yang, K.; Su, J. Terahertz Electric Field Serves as a Freeze Button for Water. Phys. E Low-Dimens. Syst. Nanostructures 2024, 163, 116037. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Li, S.; Su, J. Water Jumps over a Nanogap between Two Disjoint Carbon Nanotubes Assisted by Terahertz Electric Field. Phys. Fluids 2024, 36, 022006. [Google Scholar] [CrossRef]
- Yu, Y.-S.; Tan, R.-R.; Zhu, D.; Ding, H.-M. Terahertz Waves Modulate Serum Protein Adsorption on Graphene Oxide by Reshaping Interfacial Hydration and Protein Dynamics. Langmuir 2026, 42, 3063–3070. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.; Dornberger, U.; Rozners, E.; Fritzsche, H.; Strömberg, R.; Gräslund, A. FTIR Spectroscopic Studies of Oligonucleotides That Model a Triple-Helical Domain in Self-Splicing Group I Introns. Biochemistry 1997, 36, 15463–15471. [Google Scholar] [CrossRef] [PubMed]
- Globus, T.R.; Woolard, D.L.; Samuels, A.C.; Gelmont, B.L.; Hesler, J.; Crowe, T.W.; Bykhovskaia, M. Submillimeter-Wave Fourier Transform Spectroscopy of Biological Macromolecules. J. Appl. Phys. 2002, 91, 6105–6113. [Google Scholar] [CrossRef]
- Globus, T.; Woolard, D.; Crowe, T.W.; Khromova, T.; Gelmont, B.; Hesler, J. Terahertz Fourier Transform Characterization of Biological Materials in a Liquid Phase. J. Phys. Appl. Phys. 2006, 39, 3405. [Google Scholar] [CrossRef]
- Maréchal, Y. The Molecular Structure of Liquid Water Delivered by Absorption Spectroscopy in the Whole IR Region Completed with Thermodynamics Data. J. Mol. Struct. 2011, 1004, 146–155. [Google Scholar] [CrossRef]
- Michaud-Agrawal, N.; Denning, E.J.; Woolf, T.B.; Beckstein, O. MDAnalysis: A Toolkit for the Analysis of Molecular Dynamics Simulations. J. Comput. Chem. 2011, 32, 2319–2327. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array Programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- McKinney, W. Data Structures for Statistical Computing in Python. scipy 2010, 445, 51–56. [Google Scholar]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Waskom, M.L. Seaborn: Statistical Data Visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Quemener, E.; Corvellec, M. SIDUS—The Solution for Extreme Deduplication of an Operating System. Linux J. 2013, 2013, 3. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Zhang, Q.; Valério, M.; Wang, K.; Yang, L.; Wang, S.; Souza, P.C.T.; Gong, Y. Enhanced Condensation of RNA Repeats Induced by Terahertz Oscillatory Fields. Molecules 2026, 31, 1903. https://doi.org/10.3390/molecules31111903
Zhang Q, Valério M, Wang K, Yang L, Wang S, Souza PCT, Gong Y. Enhanced Condensation of RNA Repeats Induced by Terahertz Oscillatory Fields. Molecules. 2026; 31(11):1903. https://doi.org/10.3390/molecules31111903
Chicago/Turabian StyleZhang, Qin, Mariana Valério, Kaicheng Wang, Lixia Yang, Shaomeng Wang, Paulo C. T. Souza, and Yubin Gong. 2026. "Enhanced Condensation of RNA Repeats Induced by Terahertz Oscillatory Fields" Molecules 31, no. 11: 1903. https://doi.org/10.3390/molecules31111903
APA StyleZhang, Q., Valério, M., Wang, K., Yang, L., Wang, S., Souza, P. C. T., & Gong, Y. (2026). Enhanced Condensation of RNA Repeats Induced by Terahertz Oscillatory Fields. Molecules, 31(11), 1903. https://doi.org/10.3390/molecules31111903