
Evaluation of Bioactivity of Essential Oils: Cytotoxic/Genotoxic Effects on Colorectal Cancer Cell Lines, Antibacterial Activity, and Survival of Lactic Acid Bacteria

 , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Cytotoxic Effect of EOs and EO Blends
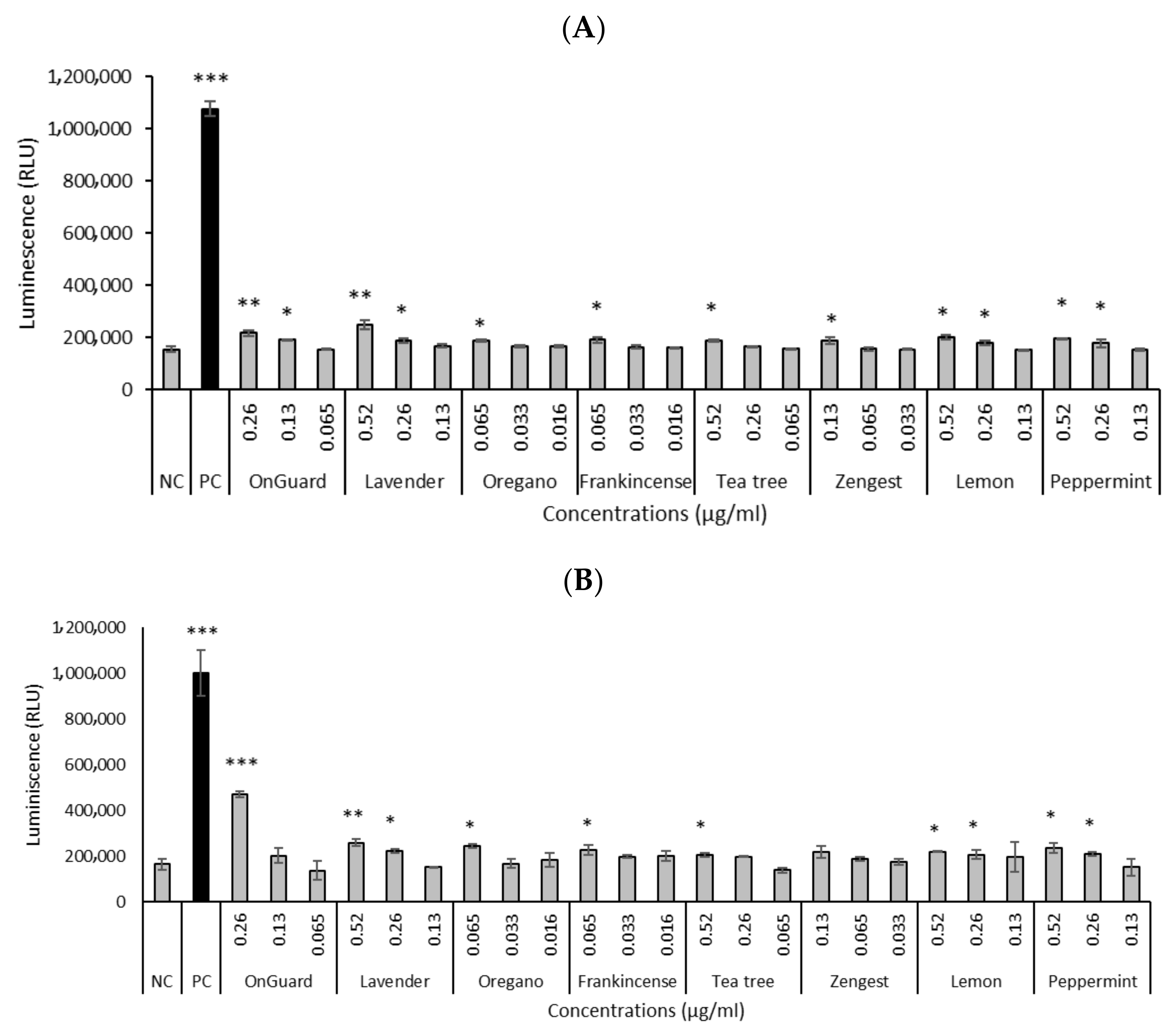
2.2. DNA-Damaging Effects of EOs and EO Blends
2.3. ROS Production
2.4. Determination of the Antibacterial Activity of EOs
2.5. Evaluation of Minimum Inhibitory Concentration (MIC) in Liquid Medium
2.6. Determination of the Number of Viable Bacteria
3. Discussion
4. Materials and Methods
4.1. Essential Oils
4.2. Cell Culture
4.3. Bacterial Strains
4.4. Determination of Cytotoxicity (MTT Assay)
4.5. Determination of Genotoxicity (Comet Assay, SCGE)
4.6. Determination of ROS Production
4.7. Evaluation of Antibacterial Activity
4.8. Evaluation of MIC in Liquid Medium
4.9. Determination of the Number Viable Bacteria
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Cancer Risk Factors Collaborators. The global burden of cancer attributable to risk factors, 2010–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 563–591. [Google Scholar] [CrossRef] [PubMed]
- Sakata, S.; Larson, D.W. Targeted therapy for colorectal cancer. Surg. Oncol. Clin. N. Am. 2022, 31, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, L.; Huntington, K.E.; El-Deiry, W.S. Immunotherapy for colorectal cancer: Mechanisms and predictive biomarkers. Cancers 2022, 14, 1028. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Kamiloglu, S.; Petroni, K.; Mishra, A.P.; Monser-rat-Mesquida, M.; Sureda, A.; Martorell, M.; Aidarbekovna, D.S.; Yessimsiitova, Z.; et al. Recent advances in the therapeutic potential of emodin for human health. Biomed. Pharmacother. 2022, 154, 113555. [Google Scholar] [CrossRef]
- Dhyani, P.; Quispe, C.; Sharma, E.; Bahukhandi, A.; Sati, P.; Attri, D.C.; Cho, W.C.; Szopa, A.; Sharifi-Rad, J.; Docea, A.O.; et al. Anticancer potential of alkaloids: A key emphasis to colchicine, vinblastine, vincris-tine, vindesine, vinorelbine and vincamine. Cancer Cell Int. 2022, 22, 206. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Patra, J.K.; Singh, Y.D.; Panda, M.K.; Das, G.; Adetunji, C.O.; Michael, O.S.; Sytar, O.; Polito, L.; et al. Paclitaxel: Application in modern oncology and nanomedicine-based cancer therapy. Oxidative Med. Cell. Longev. 2021, 2021, 3687700. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Sousa, C.; Duarte, D.; Silva-Lima, B.; Videira, M. Repurposing natural dietary flavonoids in the modulation of cancer tumorigenesis: Decrypting the molecular targets of naringenin, hesperetin, and myricetin. Nutr. Cancer 2022, 74, 1188–1202. [Google Scholar] [CrossRef]
- Adlard, E.R.K.; Can Başer, H.; Buchbauer, G. Handbook of Essential Oils: Science, Technology and Applications, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; Volume 79, p. 791. [Google Scholar]
- Blowman, K.; Magalhães, M.; Lemos, M.F.L.; Cabral, C.; Pires, I.M. Anticancer properties of essential oils and other natural products. Evid. Based Complement. Altern. Med. 2018, 2018, 3149362. [Google Scholar] [CrossRef]
- Gautam, N.; Mantha, A.; Mittal, S. Essential oils and their constituents as anticancer agents: A mechanistic view. BioMed Res. Int. 2014, 2014, 154106. [Google Scholar] [CrossRef] [PubMed]
- Azimi, M.; Mehrzad, J.; Ahmadi, A.; Ahmadi, E.; Ranjbary, A.G. Apoptosis induced by Ziziphora tenuior essential oil in human colorectal cancer cells. Biomed. Res. Int. 2021, 2021, 5522964. [Google Scholar] [CrossRef] [PubMed]
- Petrocelli, G.; Farabegoli, F.; Valerii, M.C.; Giovannini, C.; Sardo, A.; Spisni, E. Molecules present in plant essential oils for prevention and treatment of colorectal cancer (CRC). Molecules 2021, 26, 885. [Google Scholar] [CrossRef] [PubMed]
- Puškárová, A.; Bučková, M.; Kraková, L.; Pangallo, D.; Kozics, K. The antibacterial and antifungal activity of six essential oils and their cyto/genotoxicity to human HEL 12469 cells. Sci. Rep. 2017, 7, 8211. [Google Scholar] [CrossRef] [PubMed]
- Kozics, K.; Bučková, M.; Puškárová, A.; Kalászová, V.; Cabicarová, T.; Pangallo, D. The effect of ten essential oils on several cutaneous drug-resistant microorganisms and their cyto/genotoxic and antioxidant properties. Molecules 2019, 24, 4570. [Google Scholar] [CrossRef] [PubMed]
- Clerck, C.D.; Maso, S.D.; Parisi, O.; Dresen, F.; Zhiri, A.; Jijakli, M.H. Screening of antifungal and antibacterial activity of 90 commercial essential oils against 10 pathogens of agronomical importance. Foods 2020, 9, 1418. [Google Scholar] [CrossRef]
- Puvača, N.; Milenković, J.; Galonja Coghill, T.; Bursić, V.; Petrović, A.; Tanasković, S.; Pelić, M.; Ljubojević Pelić, D.; Miljković, T. Antimicrobial activity of selected essential oils against selected pathogenic bacteria: In vitro study. Antibiotics 2021, 10, 546. [Google Scholar] [CrossRef]
- Gheorghita, D.; Robu, A.; Antoniac, A.; Antoniac, I.; Ditu, L.M.; Raiciu, A.D.; Tomescu, J.; Grosu, E.; Saceleanu, A. In vitro antibacterial activity of some plant essential oils against four different microbial strains. Appl. Sci. 2022, 12, 9482. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Al-Yami, A.M.; Al-Mousa, A.T.; Al-Otaibi, S.A.; Khalifa, A.Y. Lactobacillus species as probiotics: Isolation sources and health benefits. J. Pure Appl. Microbiol. 2022, 16, 2270–2291. [Google Scholar] [CrossRef]
- Kim, Y.S.; Milner, J.A. Dietary modulation of colon cancer risk. J. Nutr. 2007, 137, 2576S–2579S. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ishida, H.; Kobo, K.; Furukawa, I.; Ikeda, Y.; Yonaha, M.; Aniya, Y.; Ohigashi, H. Suppressive effects of Okinawan food items on free radical generation from stimulated leukocytes and identification of some active constituents: Implications for the prevention of inflammation-associated carcinogenesis. Asian Pac. J. Cancer Prev. 2005, 6, 437. [Google Scholar] [PubMed]
- Aggarwal, B.B.; Kunnumakkara, A.B.; Harikumar, K.B.; Tharakan, S.T.; Sung, B.; Anand, P. Potential of spice-derived phytochemicals for cancer prevention. Planta Med. 2008, 74, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Khadir, A.; Sobeh, M.; Gad, H.A.; Benbelaid, F.; Bendahou, M.; Peixoto, H.; Sporer, F.; Ashour, M.L.; Wink, M. Chemical composition and biological activity of the essential oil from Thymus lanceolatus. Z. Naturforsch. C 2016, 71, 155–163. [Google Scholar] [CrossRef]
- Aydın, E.; Turkez, H.; Tasdemir, S.; Hacımuftuoglu, F. Anticancer, antioxidant and cytotoxic potential of thymol in vitro brain tumor cell model. CNS Agents Med. Chem. 2017, 17, 116–122. [Google Scholar] [CrossRef]
- Jamali, T.; Kavoosi, G.; Ardestani, S.K. In-vitro and in-vivo anti-breast cancer ac-tivity of OEO (Oliveria decumbens vent essential oil) through promoting apoptosis and immunomodulatory effects. J. Ethnopharmacol. 2020, 248, 112313. [Google Scholar] [CrossRef]
- Begnini, K.R.; Nedel, F.; Lund, R.G.; Carvalho, P.H.D.A.; Rodrigues, M.R.A.; Beira, F.T.A.; Del-Pino, F.A.B. Composition and antiproliferative effect of essential oil of Origanum vulgare against tumor cell lines. J. Med. Food 2014, 17, 1129–1133. [Google Scholar] [CrossRef]
- Zengin, G.; Ferrante, C.; Orlando, G.; Zheleva-Dimitrova, D.; Gevrenova, R.; Recinella, L.; Chiavaroli, A.; Leone, S.; Brunetti, L.; Aumeeruddy, M.Z.; et al. Chemical profiling and pharmaco-toxicological activity of Origanum sipyleum extracts: Exploring for novel sources for potential therapeutic agents. J. Food Biochem. 2019, 43, e13003. [Google Scholar] [CrossRef]
- Fahmy, M.A.; Farghaly, A.A.; Hassan, E.E.; Hassan, E.M.; Hassan, Z.M.; Mahmoud, K.; Omara, E.A. Evaluation of the anti-cancer/anti-mutagenic efficiency of Lavandula officinalis essential oil. Asian Pac. J. Cancer Prev. 2022, 23, 1215. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Jadegoud, Y.; Nagana Gowda, G.A.; Patil, B.S. Bioactive compounds from sour orange inhibit colon cancer cell proliferation and induce cell cycle arrest. J. Agric. Food Chem. 2010, 58, 180–186. [Google Scholar] [CrossRef]
- Thapa, D.; Richardson, A.J.; Zweifel, B.; Wallace, R.J.; Gratz, S.W. Genoprotective effects of essential oil compounds against oxidative and methylated DNA damage in human colon cancer cells. J. Food. Sci. 2019, 84, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Blažíčková, M.; Bučková, M.; Kozics, K. Characterization of the effects of thymol derivatives on colorectal cancer spheroids. Neoplasma 2024, 71, 2. [Google Scholar] [CrossRef] [PubMed]
- Malekifard, F.; Esmaeilnejad, B. In vitro anthelmintic efficacy of medicinal plant essential oils against Marshallagia marshalli: Evidence on oxidative/nitrosative stress biomarkers, DNA damage, and egg hatchability. Vet. Parasitol. 2024, 327, 110138. [Google Scholar]
- Nakamura, H.; Takada, K. Reactive oxygen species in cancer: Current findings and future directions. Cancer Sci. 2021, 112, 3945–3952. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef]
- Sœur, J.; Marrot, L.; Perez, P.; Iraqui, I.; Kienda, G.; Dardalhon, M.; Meunier, J.-R.; Averbeck, D.; Huang, M.-E. Selective cytotoxicity of Aniba rosaeodora essential oil towards epidermoid cancer cells through induction of apoptosis. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2011, 718, 24–32. [Google Scholar] [CrossRef]
- Paik, S.Y.; Koh, K.H.; Beak, S.M.; Paek, S.H.; Kim, J.A. The essential oils from Zanthoxylum schinifolium pericarp induce apoptosis of HepG2 human hepatoma cells through increased production of reactive oxygen species. Biol. Pharm. Bull. 2005, 28, 802–807. [Google Scholar] [CrossRef]
- Islam, A.; Chang, Y.C.; Tsao, N.W.; Wang, S.Y.; Chueh, P.J. Calocedrus formosana essential oils induce ROS-mediated autophagy and apoptosis by targeting SIRT1 in colon cancer cells. Antioxidants 2024, 13, 284. [Google Scholar] [CrossRef]
- Balan, D.J.; Rajavel, T.; Das, M.; Sathya, S.; Jeyakumar, M.; Devi, K.P. Thymol in-duces mitochondrial pathway-mediated apoptosis via ROS generation, macromolecu-lar damage, and SOD diminution in A549 cells. Pharmacol. Rep. 2021, 73, 240–254. [Google Scholar] [CrossRef]
- Satooka, H.; Kubo, I. Effects of thymol on B16-F10 melanoma cells. J. Agric. Food Chem. 2012, 60, 2746–2752. [Google Scholar] [CrossRef]
- Yenigun, V.B.; Kocyigit, A.; Kanimdan, E.; Balkan, E.; Gul, A.Z. Copper (II) in-creases anti-proliferative activity of thymoquinone in colon cancer cells by increasing genotoxic, apoptotic, and reactive oxygen species generating effects. Toxicon 2024, 250, 108103. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Bučková, M.; Puškárová, A.; Kalászová, V.; Kisová, Z.; Pangallo, D. Essential oils against multidrug-resistant gram-negative bacteria. Biologia 2018, 73, 803–808. [Google Scholar] [CrossRef]
- Fancello, F.; Petretto, G.L.; Marceddu, S.; Venditti, T.; Pintore, G.; Zara, G.; Mannazzu, I.; Budroni, M.; Zara, S. Antimicrobial activity of gaseous Citrus limon var pompia leaf essential oil against Listeria monocytogenes on ricotta salata cheese. Food Microbiol. 2020, 87, 103386. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Tiihonen, K.; Kettunen, H.; Peuranen, S.; Schulze, H.; Rautonen, N. In vitro effects of essential oils on potential pathogens and beneficial members of the normal microbiota. Vet. Med. 2010, 55, 71–78. [Google Scholar] [CrossRef]
- Badia, V.; de Oliveira, M.S.R.; Polmann, G.; Milkievicz, T.; Galvão, A.C.; da Silva Robazza, W. Effect of the addition of antimicrobial oregano (Origanum vulgare) and rosemary (Rosmarinus officinalis) essential oils on lactic acid bacteria growth in re-frigerated vacuum-packed Tuscan sausage. Braz. J. Microbiol. 2020, 51, 289–301. [Google Scholar] [CrossRef]
- De Souza, G.T.; De Carvalho, R.J.; De Sousa, J.P.; Tavares, J.F.; Schaffner, D.; De Souza, E.L.; Magnani, M. Effects of the essential oil from Origanum vulgare L. on survival of pathogenic bacteria and starter lactic acid bacteria in semihard cheese broth and slurry. J. Food Prot. 2016, 79, 246–252. [Google Scholar] [CrossRef]
- Horošová, K.; Bujňáková, D.; Kmeť, V. Effect of oregano essential oil on chicken lactobacilli and E. coli. Folia Microbiol. 2006, 51, 278–280. [Google Scholar] [CrossRef]
- Moritz, C.M.F.; Rall, V.L.M.; Saeki, M.J.; Fernandes Júnior, A. Inhibitory effect of essential oils against Lactobacillus rhamnosus and starter culture in fermented milk during its shelf-life period. Braz. J. Microbiol. 2012, 43, 1147–1156. [Google Scholar] [CrossRef]
- Dunn, L.L.; Davidson, P.M.; Critzer, F.J. Antimicrobial efficacy of an array of essential oils against lactic acid bacteria. J. Food Sci. 2016, 81, 438–444. [Google Scholar] [CrossRef]
- Woods, G.L. In vitro testing of antimicrobial agents. Infect. Dis. Clin. N. Am. 1995, 9, 463–481. [Google Scholar] [CrossRef]
- Sarabi-Jamab, M.; Niazmand, R. Effect of essential oil of Mentha piperita and Ziziphora clinopodioides on Lactobacillus acidophilus activity as bioyogurt starter culture. Am.-Eurasian J. Agric. Environ. Sci. 2009, 6, 129–131. [Google Scholar]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot09550. [Google Scholar] [CrossRef]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications, and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Ponce, A.G.; Fritz, R.; Del Valle, C.; Roura, S.I. Antimicrobial activity of essential oils on the native microflora of organic Swiss chard. LWT-Food Sci. Technol. 2003, 36, 679–684. [Google Scholar] [CrossRef]
- Jett, B.D.; Hatter, K.L.; Huycke, M.M.; Gilmore, M.S. Simplified agar plate method for quantifying viable bacteria. Biotechniques 1997, 23, 648–650. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| EOs/EO Blends | IC50 (µg/mL) | |
|---|---|---|
| HT-29 | HCT-116 | |
| LE | 0.62 | 0.34 |
| L | 0.38 | 0.29 |
| OR | 0.056 | 0.052 |
| OG | 0.33 | 0.29 |
| F | 0.083 | 0.05 |
| ZG | 0.05 | 0.05 |
| TT | 0.42 | 0.45 |
| P | 0.67 | 0.48 |
| (A) | ||||||||
| Concentrations of EOs and EO blends (µg/mL) | ||||||||
| 0 | 8 × 10−3 | 1.7 × 10−2 | 3.3 × 10−2 | 6.5 × 10−2 | 0.13 | 0.26 | 0.52 | |
| OnGuard | 9.11 ± 1.15 | 8.52 ± 2.03 | 8.92 ± 2.03 | 9.02 ± 1.05 | 10.36 ± 2.23 | 17.02 ± 1.23 ** | ND | ND |
| Lavender | 9.95 ± 2.11 | 9.08 ± 1.68 | 10.15 ± 2.47 | 9.22 ± 1.63 | 9.72 ± 1.51 | 10.67 ± 1.12 | 14.74 ± 1.4 * | 31.17 ± 1.76 *** |
| Oregano | 8.17 ± 1.25 | 8.86 ± 1.08 | 9.31 ± 0.65 | 14.12 ± 0.33 * | 24.31 ± 1.84 *** | 35 ± 2.15 *** | ND | ND |
| Tea tree | 7.52 ± 1.81 | 8.76 ± 0.84 | 8.12 ± 0.88 | 9.01 ± 1.05 | 8.10 ± 0.75 | 8.85 ± 2.21 | 11.96 ± 0.71 * | 21.38 ± 2.68 ** |
| Zengest | 9.39 ± 1.03 | 8.22 ± 0.93 | 8.12 ± 0.88 | 10.42 ± 2.12 | 9.90 ± 1.32 | 37.22 ± 4.58 *** | ND | ND |
| Lemon | 8.25 ± 1.34 | 9.06 ± 0.79 | 8.54 ± 1.22 | 9.11 ± 1.18 | 10.19 ± 0.65 | 10.45 ± 2.02 | 15.52 ± 1.82 * | 22.59 ± 2.72 ** |
| Peppermint | 7.52 ± 1.81 | 8.76 ± 0.84 | 8.12 ± 0.88 | 9.01 ± 1.05 | 8.10 ± 0.75 | 9.23 ± 0.44 | 12.26 ± 1.62 * | 22.96 ± 2.68 ** |
| Frankincense | 8.82 ± 1.21 | 10.16 ± 1.94 | 9.36 ± 1.27 | 10.76 ± 1.83 | 15.46 ± 1.13 * | ND | ND | ND |
| PC | 52.52 ± 2.83 | |||||||
| (B) | ||||||||
| Concentrations of EOs and EO blends (µg/mL) | ||||||||
| 0 | 8 × 10−3 | 1.7 × 10−2 | 3.3 × 10−2 | 6.5 × 10−2 | 0.13 | 0.26 | 0.52 | |
| OnGuard | 8.09 ± 0.95 | 10.02 ± 2.18 | 9.22 ± 1.93 | 9.58 ± 1.55 | 10.02 ± 2.65 | 12.24 ± 2.01 * | 17.82 ± 1.23 ** | ND |
| Lavender | 8.95 ± 0.71 | 8.28 ± 1.55 | 7.15 ± 0.77 | 8.85 ± 1.33 | 9.04 ± 1.03 | 11.32 ± 2.05 | 15.57 ± 0.70 *** | ND |
| Oregano | 6.75 ± 0.63 | 7.02 ± 0.95 | 6.85 ± 1.23 | 8.79 ± 2.91 | 15.75 ± 0.93 * | 20.00 ± 2.52 *** | ND | ND |
| Tea tree | 7.52 ± 1.81 | 7.67 ± 0.91 | 8.09 ± 0.73 | 8.98 ± 1.52 | 8.10 ± 1.05 | 9.52 ± 0.64 | 12.85 ± 0.87 * | 19.23 ± 1.19 ** |
| Zengest | 9.02 ± 1.25 | 7.98 ± 0.88 | 9.48 ± 1.72 | 10.28 ± 1.71 | 11.34 ± 3.27 | 30.09 ± 3.73 *** | ND | ND |
| Lemon | 8.74 ± 0.78 | 9.86 ± 1.32 | 9.42 ± 1.20 | 10.01 ± 1.21 | 9.45 ± 1.21 | 11.07 ± 3.06 | 19.02 ± 2.11 ** | 27.89 ± 3.53 *** |
| Peppermint | 9.03 ± 1.05 | 8.66 ± 0.72 | 8.56 ± 1.13 | 9.01 ± 1.11 | 8.98 ± 1.08 | 10.14 ± 1.51 | 17.78 ± 2.34 ** | 22.01 ± 1.30 ** |
| Frankincense | 10.23 ± 2.12 | 9.98 ± 1.53 | 10.06 ± 1.70 | 12.17 ± 2.80 | 20.48 ± 3.06 *** | ND | ND | ND |
| PC | 52.52 ± 2.83 | |||||||
| Strains | OR | TT | OG |
|---|---|---|---|
| L. plantarum | 0.05 | 0.5 | 0.5 |
| L. rhamnosus | 0.05 | 0.5 | 0.5 |
| L. pentosus | 0.05 | 0.25 | 0.25 |
| L. paracasei | 0.05 | 0.25 | 0.25 |
| L. brevis | 0.05 | 0.5 | 0.5 |
| W. coagulans | 0.05 | 0.5 | 0.5 |
| Strains (Log CFU/g) | Control | LE | P | L | F | ZG |
|---|---|---|---|---|---|---|
| L. plantarum | 6.60 ± 0.05 | 7.61 ± 0.51 | 7.52 ± 0.55 | 7.60 ± 0.51 | 6.55 ± 0.05 | 6.61 ± 0.05 |
| L. rhamnosus | 6.22 ± 0.06 | 7.21 ± 0.52 | 7.22 ± 0.59 | 7.20 ± 0.51 | 6.22 ± 0.09 | 6.12 ± 0.05 |
| L. pentosus | 6.52 ± 0.09 | 6.12 ± 0.11 | 6.22 ± 0.12 | 6.42 ± 0.09 | 6.45 ± 0.11 | 6.35 ± 0.12 |
| L. paracasei | 6.85 ± 0.15 | 6.75 ± 0.14 | 6.65 ± 0.11 | 6.81 ± 0.11 | 6.75 ± 0.13 | 6.85 ± 0.11 |
| L. brevis | 6.74 ± 0.08 | 6.64 ± 0.11 | 6.71 ± 0.09 | 6.70 ± 0.12 | 6.68 ± 0.08 | 6.70 ± 0.08 |
| W. coagulans | 5.61 ± 0.11 | 5.66 ± 0.16 | 5.65 ± 0.12 | 5.56 ± 0.15 | 5.55 ± 0.12 | 5.62 ± 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozics, K.; Mesárošová, M.; Šramková, M.; Bučková, M.; Puškárová, A.; Galová, D.; Pangallo, D. Evaluation of Bioactivity of Essential Oils: Cytotoxic/Genotoxic Effects on Colorectal Cancer Cell Lines, Antibacterial Activity, and Survival of Lactic Acid Bacteria. Molecules 2025, 30, 890. https://doi.org/10.3390/molecules30040890
Kozics K, Mesárošová M, Šramková M, Bučková M, Puškárová A, Galová D, Pangallo D. Evaluation of Bioactivity of Essential Oils: Cytotoxic/Genotoxic Effects on Colorectal Cancer Cell Lines, Antibacterial Activity, and Survival of Lactic Acid Bacteria. Molecules. 2025; 30(4):890. https://doi.org/10.3390/molecules30040890
Chicago/Turabian StyleKozics, Katarína, Monika Mesárošová, Monika Šramková, Mária Bučková, Andrea Puškárová, Dominika Galová, and Domenico Pangallo. 2025. "Evaluation of Bioactivity of Essential Oils: Cytotoxic/Genotoxic Effects on Colorectal Cancer Cell Lines, Antibacterial Activity, and Survival of Lactic Acid Bacteria" Molecules 30, no. 4: 890. https://doi.org/10.3390/molecules30040890
APA StyleKozics, K., Mesárošová, M., Šramková, M., Bučková, M., Puškárová, A., Galová, D., & Pangallo, D. (2025). Evaluation of Bioactivity of Essential Oils: Cytotoxic/Genotoxic Effects on Colorectal Cancer Cell Lines, Antibacterial Activity, and Survival of Lactic Acid Bacteria. Molecules, 30(4), 890. https://doi.org/10.3390/molecules30040890