

Systemic Administration of the Phytochemical, Myricetin, Attenuates the Excitability of Rat Nociceptive Secondary Trigeminal Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
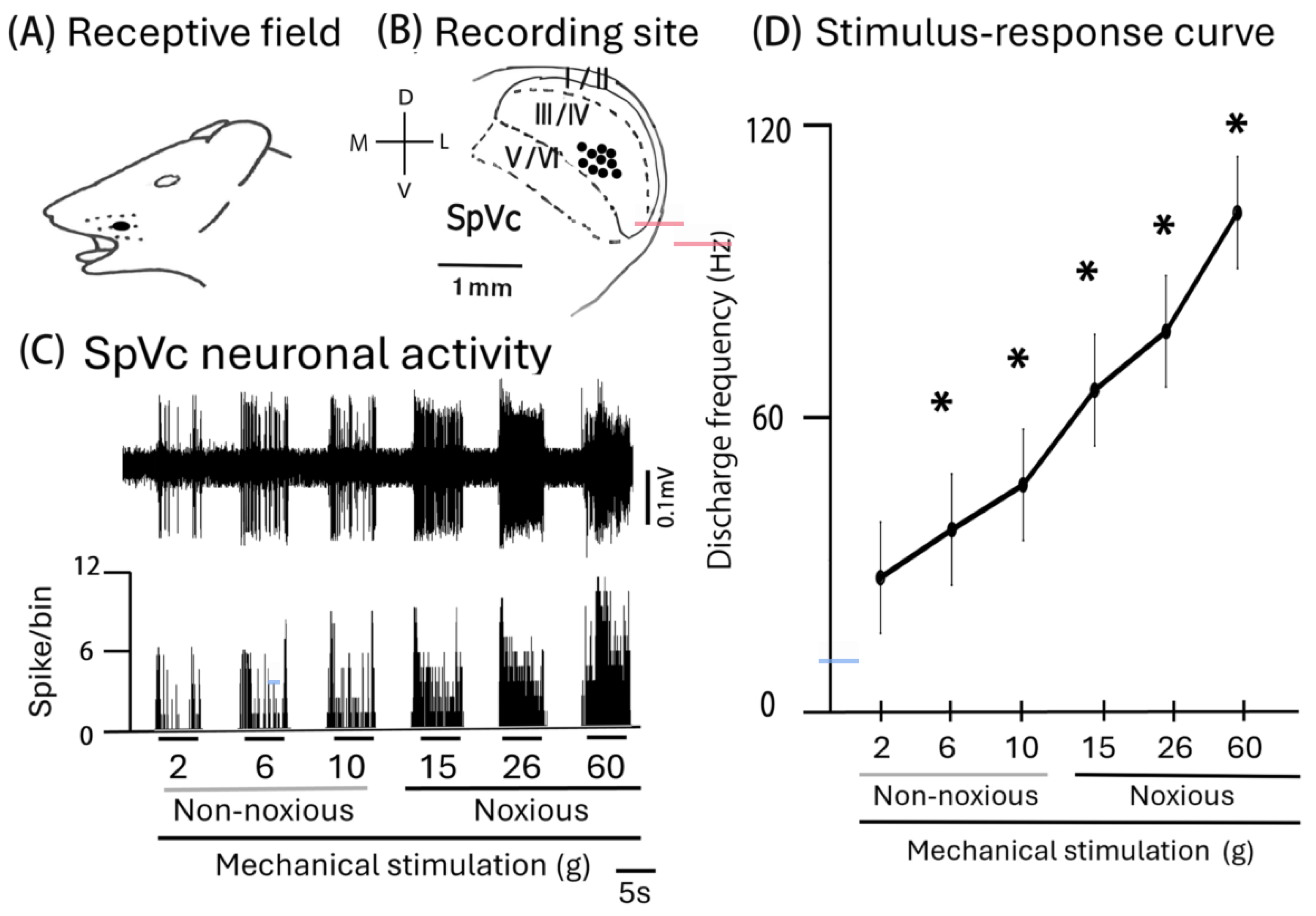
2.1. General Characteristics of SpVc WDR Neurons That Supply Trigeminal Nerve Fibers to the Facial Skin
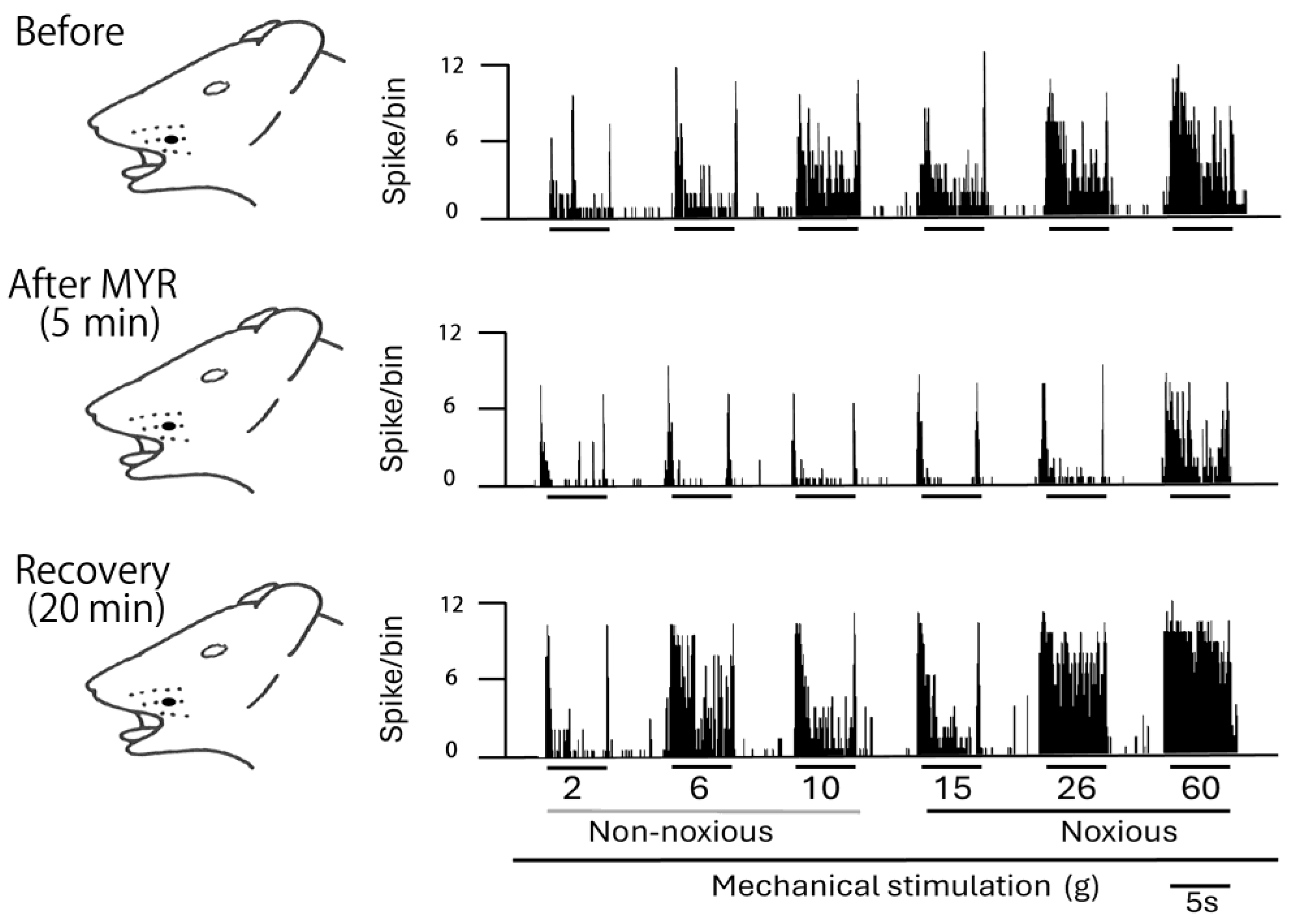
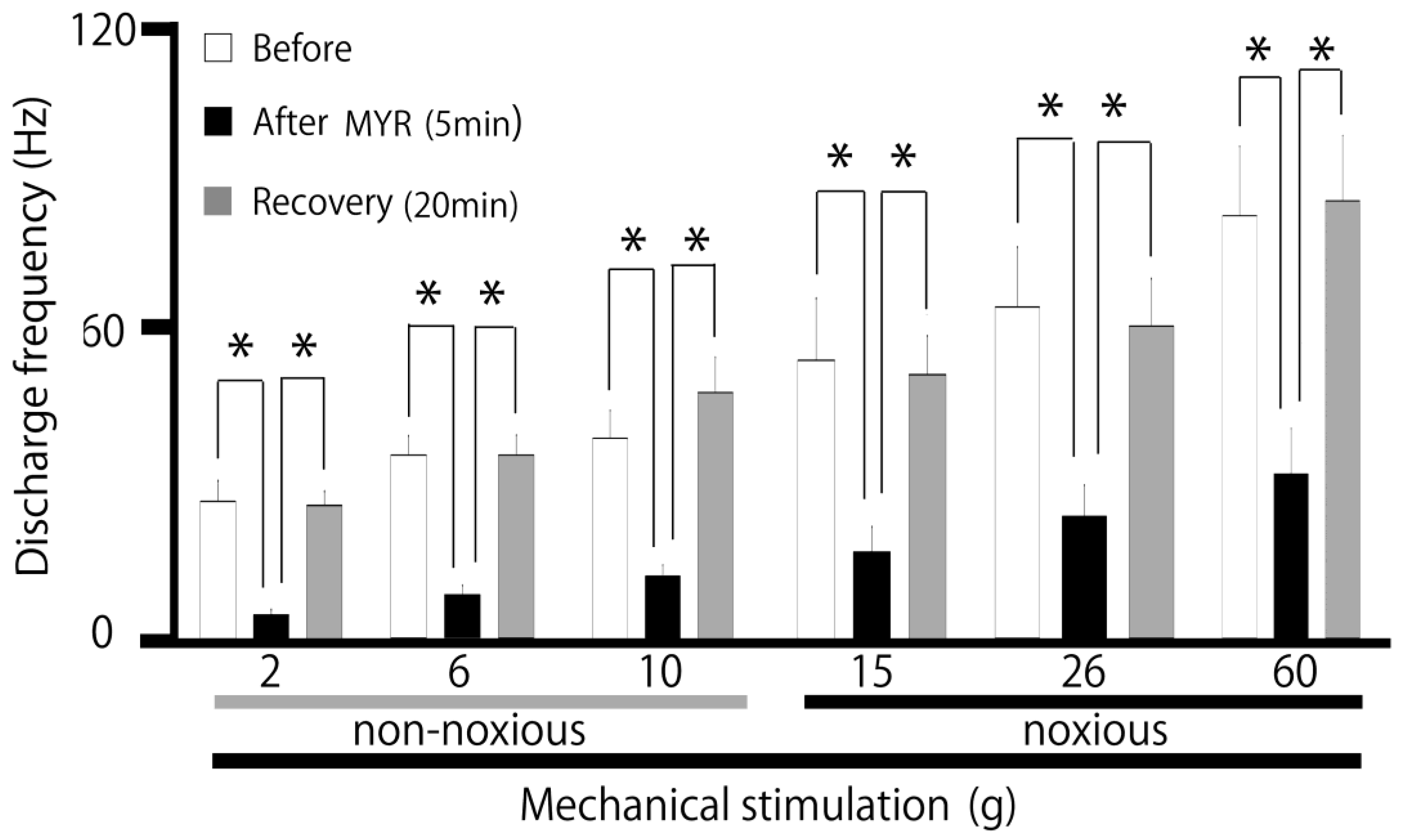
2.2. Influence of Intravenous MYR on the Excitability of SpVc WDR Neurons When Exposed to Non-Noxious and Noxious Stimuli
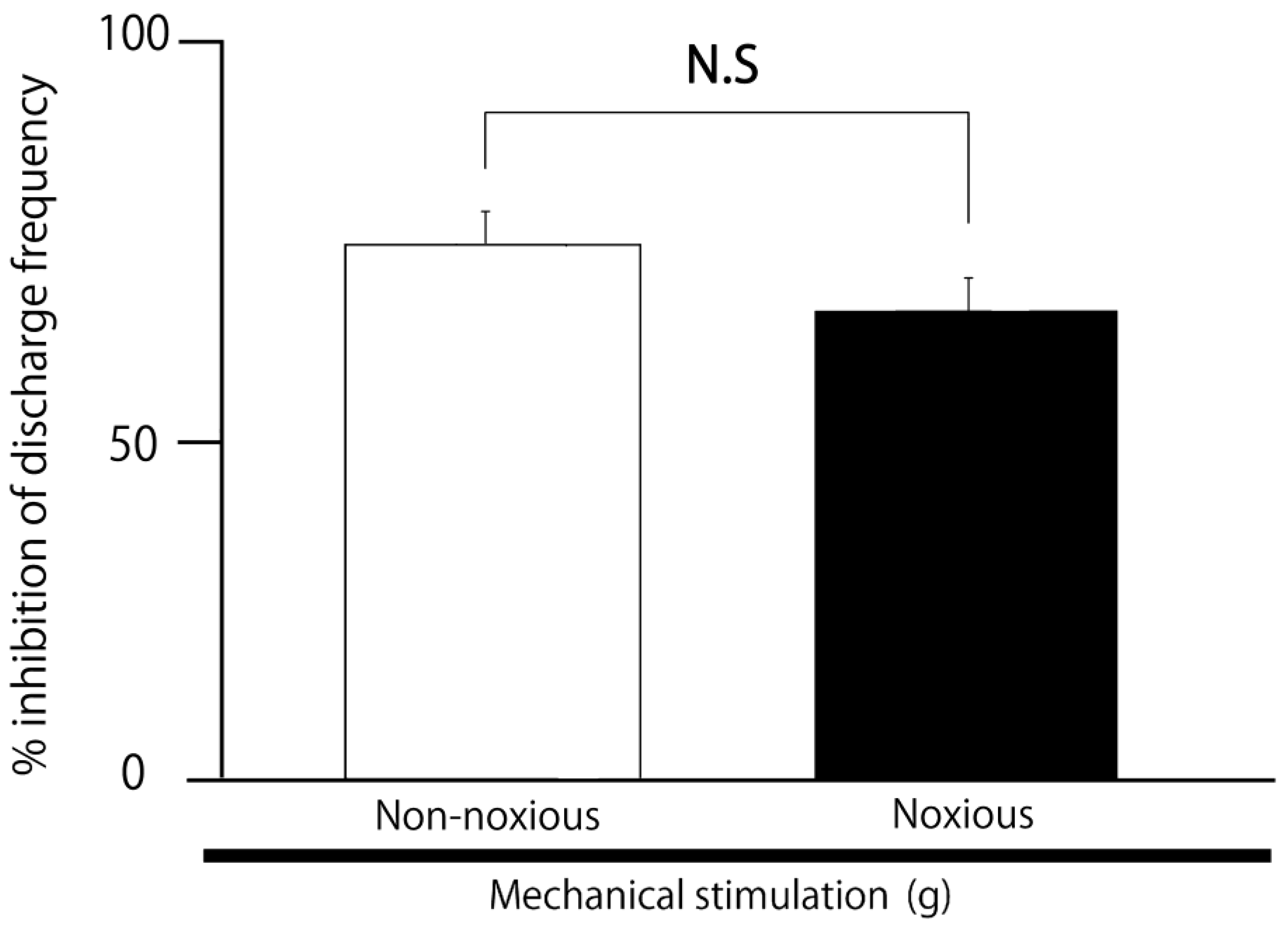
2.3. SpVc WDR Neuronal Activity in Response to Noxious vs. Non-Noxious Stimuli After MYR
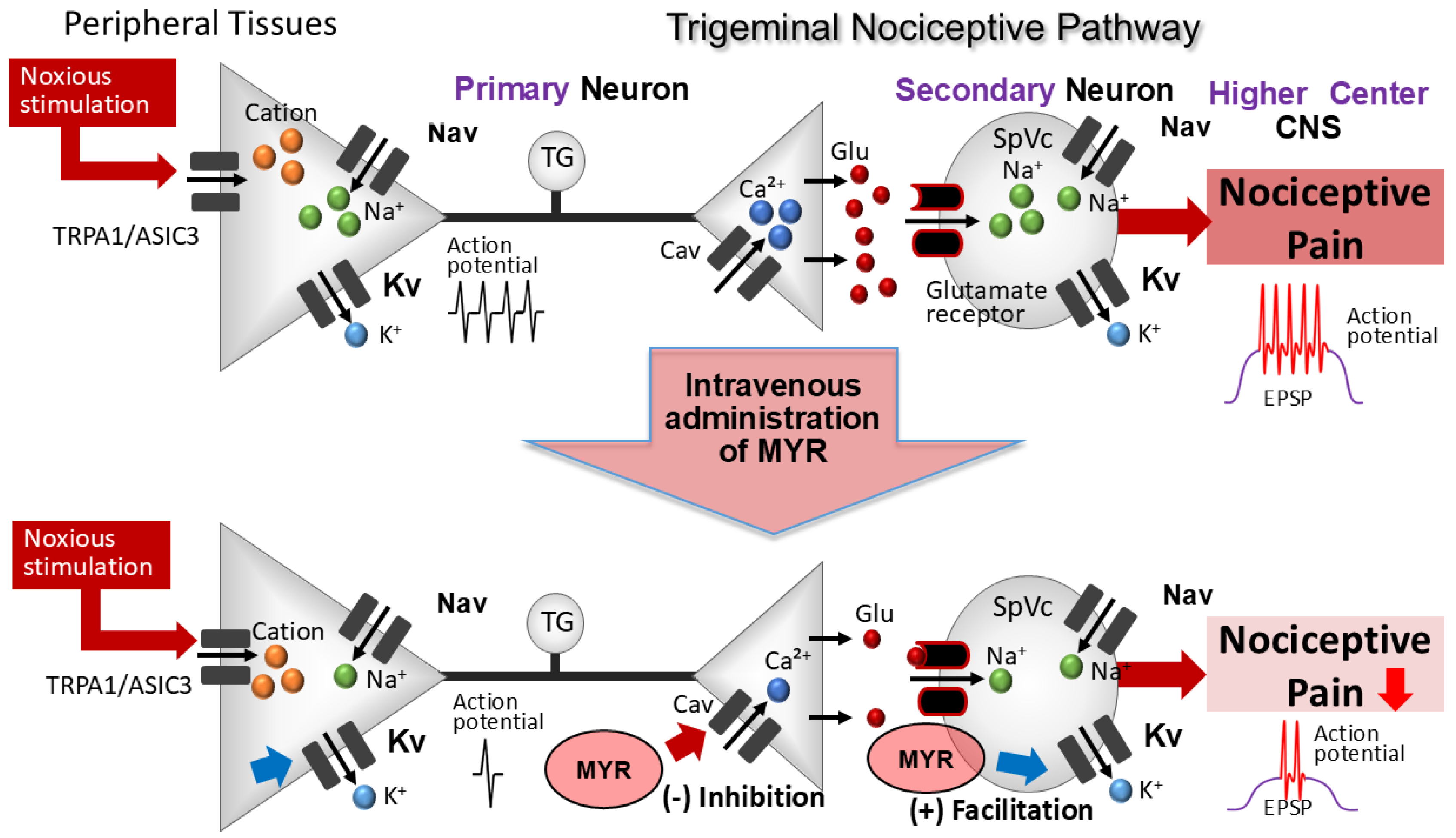
3. Discussion
3.1. Acute Intravenous Administration of MYR Reduced the Excitability of SpVc WDR Neurons
3.2. The Excitability of SpVc WDR Neurons Was Suppressed by MYR via Mechanisms at Both Peripheral and Central Levels
3.3. The Significance of MYR’s Suppression of SpVc Neuron Excitability in Response to Nociceptive Stimulation
4. Materials and Methods
4.1. Extracellular Single-Unit Recording of WDR Neuronal Activity in the SpVc
4.2. Experimental Protocols
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iwata, K.; Takeda, M.; Oh, S.; Shinoda, M. Neurophysiology of Orofacial Pain. In Contemporary Oral Medicine; Farah, C.S., Balasubramaniam, R., McCullough, M.J., Eds.; Springer International Publishing: New York, NY, USA, 2017. [Google Scholar]
- Sessle, B.J. Peripheral and central mechanisms of orofacial pain and their clinical correlates. Minerva Anestesiol. 2005, 71, 117–136. [Google Scholar]
- Shinoda, M.; Imamura, Y.; Hayashi, Y.; Noma, N.; Okada-Ogawa, A.; Hitomi, S.; Iwata, K. Orofacial neuropathic pain-basic research and their clinical relevancies. Front. Mol. Neurosci. 2021, 14, 691396. [Google Scholar] [CrossRef] [PubMed]
- Sessle, B.J. Chronic orofacial pain: Models, mechanisms, and Genetic and related environmental influences. Int. J. Mol. Sci. 2021, 22, 7112. [Google Scholar] [CrossRef]
- Rao, J.K.; Mihaliak, K.; Kroenke, K.; Bradley, J.; Tierney, W.M.; Weinberger, M. Use of complementary therapies for arthritis among patients of rheumatologists. Ann. Int. Med. 1999, 131, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Konvicka, J.J.; Meyer, T.A.; McDavid, A.J.; Roberson, C.R. Complementary/alternative medicine use among chronic pain clinic patients. J. Perianesthes. Nurs. 2008, 23, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.I.; Genao, I.; Chen, I.; Mechaber, A.J.; Wood, J.A.; Faselis, C.J.; Kurz, J.; Menon, M.; O’Rorke, J.; Panda, M.; et al. Complementary and alternative medicine use by primary care patients with chronic pain. Pain Med. 2008, 9, 1065–1072. [Google Scholar] [CrossRef]
- Bauer, B.; Tilburt, C.; Sood, A.; Li, G.-X.; Wang, S.-H. Complementary and alternative medicine therapies for chronic pain. Chin. J. Integr. Med. 2016, 6, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Frémont, L. Biological effects of resveratrol. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, S. Resveratrol: From grapevines to mammalian biology. FASEB J. 2003, 17, 1975–1985. [Google Scholar] [CrossRef]
- Hakkinen, S.H.; Karenlampi, S.O.; Heinonen, I.M.; Mykkanen, H.M.; Torronen, A.R. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J. Agric. Food Chem. 1999, 47, 2274–2279. [Google Scholar] [CrossRef]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Valdez, L.B.; Alvarez, S.; Zaobornyj, T.; Boveris, A. Polyphenols and red wine as antioxidants against peroxynitrite and other oxidants. Biol. Res. 2004, 37, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Chang, C.-Y.; Wang, S.-J.; Huang, S.-K. Myricetin inhibits the release of glutamate in rat cerebrocortical nerve terminals. J. Med. Food 2015, 18, 516–523. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, T. Myricetin facilitates potassium currents and inhibits neuronal activity of PVN neurons. Neurochem. Res. 2012, 37, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Toyota, R.; Itou, H.; Sashide, Y.; Takeda, M. Suppression of the excitability of rat nociceptive primary sensory neurons following local administration of the phytochemical quercetin. J. Pain 2023, 24, 540–549. [Google Scholar] [CrossRef]
- Zhang, X.H.; Ma, Z.G.; Rowlands, D.K.; Gou, Y.L.; Fok, K.L.; Wong, H.Y.; Yu, M.K.; Tsang, L.L.; Mu, L.; Chen, L.; et al. Flavonoid myricetin modulates GABAA receptor activity through activation of Ca2 + channels and CaMK-II pathway. Evid. Based Complement. Alternat. Med. 2012, 2012, 758097. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid permeability across an in situ model of the blood-brain barrier. Free Radic. Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef]
- Kakita, K.; Tsubouchi, H.; Adachi, M.; Takehana, S.; Shimazu, Y.; Takeda, M. Local subcutaneous injection of chlorogenic acid inhibits the nociceptive trigeminal spinal nucleus caudalis neurons in rats. Neurosci. Res. 2018, 134, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Berridge, M.J.; Bootman, M.D.; Lipp, P. Calcium- a life and death signal. Nature 1998, 395, 645–648. [Google Scholar] [CrossRef]
- Zamponi, G.W.; Lewis, R.J.; Todorovic, S.M.; Arneric, S.P.; Snutch, T.P. Role of voltage-gated calcium channels in ascending pain pathways. Brain Res. Rev. 2009, 60, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Schaible, H.G.; Richter, F. Pathophysiology of pain. Langenbecks Arch. Surg. 2004, 389, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.E.; Snutch, T.P. Contributions of T-type Ca channels to the pathophysiology of pain signaling. Drug Discov. Today Dis. Mech. 2006, 3, 335–341. [Google Scholar] [CrossRef]
- Ficke, E.; Heinemann, U. Slow and fast transient potassium current in cultured rat hippocampus cells. J. Physiol. 2001, 445, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. Potassium channels and chloride channels. In Ion Channels of Excitable Membranes, 3rd ed.; Hille, B., Ed.; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 134–167. [Google Scholar]
- Pearce, R.J.; Duchen, M.R. Differential expression of membrane currents in dissociated mouse primary sensory neurons. Neuroscience 1994, 63, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K. Potassium channels as targets for the management of pain. Cent. Nerv. Syst. Agents Med. Chem. 2006, 6, 119–128. [Google Scholar] [CrossRef]
- Takeda, M.; Tanimoto, T.; Ikeda, M.; Nasu, M.; Kadoi, J.; Yoshida, S.; Matsumoto, S. Enhanced excitability of rat trigeminal root ganglion neurons via decrease in A-type potassium currents following temporomandibular joint inflammation. Neuroscience 2006, 138, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Hara, N.; Takeda, M.; Takahashi, M.; Matsumoto, S. Iontophoretic application of an A-type potassium channel blocker to the trigeminal ganglion neurons enhances the excitability of Aδ- and C-neurons innervating the temporomandibular joint. Neurosci. Res. 2012, 74, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanimoto, T.; Matsumoto, S. Change in mechanical receptive field properties induced by GABAA receptor activation in the trigeminal spinal nucleus caudalis neurons in rats. Exp. Brain Res. 2000, 134, 409–416. [Google Scholar] [CrossRef]
- Takeda, M.; Takehana, S.; Sekiguchi, K.; Kubota, Y.; Shimazu, Y. Modulatory mechanism of nociceptive neuronal activity by dietary constituent resveratrol. Int. J. Mol. Sci. 2016, 17, 1702. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.-H.; Lee, S.-H.; Jung, K.; Yoo, H.; Park, G. Inhibitory effects of myricetin on lipopolysaccharide -induced neuroinflammation. Brain Sci. 2020, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Martinez, M.; Gutierrez-Vegas, G. Myricetin inhibition of peptidoglycan-induced Cox-2 expression in H9c2 cardiomyocytes. Prev. Nutr. Food Sci. 2019, 24, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Venegas, G.; Luna, O.A.; Arreguin-Cano, J.A.; Hernandez-Bermudez, C. Myricetin blocks lipoteichoic acid-induced Cox-2 expression in human gingival fibroblasts. Cell Mol. Biol. Lett. 2014, 19, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Hagenacker, T.; Hileberand, I.; Wissmann, A.; Busselberg, D.; Schafers, M. Anti-allodynic effect of the flavonoid myricetin in a rat model of neuropathic pain: Involvement of p38 and protein kinase C meditated Ca2+ channels. Eur. J. Pain 2010, 14, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, S.; Chida, R.; Utugi, S.; Sashide, Y.; Takeda, M. Systemic Administration of the Phytochemical, Myricetin, Attenuates the Excitability of Rat Nociceptive Secondary Trigeminal Neurons. Molecules 2025, 30, 1019. https://doi.org/10.3390/molecules30051019
Yamaguchi S, Chida R, Utugi S, Sashide Y, Takeda M. Systemic Administration of the Phytochemical, Myricetin, Attenuates the Excitability of Rat Nociceptive Secondary Trigeminal Neurons. Molecules. 2025; 30(5):1019. https://doi.org/10.3390/molecules30051019
Chicago/Turabian StyleYamaguchi, Sana, Risako Chida, Syogo Utugi, Yukito Sashide, and Mamoru Takeda. 2025. "Systemic Administration of the Phytochemical, Myricetin, Attenuates the Excitability of Rat Nociceptive Secondary Trigeminal Neurons" Molecules 30, no. 5: 1019. https://doi.org/10.3390/molecules30051019
APA StyleYamaguchi, S., Chida, R., Utugi, S., Sashide, Y., & Takeda, M. (2025). Systemic Administration of the Phytochemical, Myricetin, Attenuates the Excitability of Rat Nociceptive Secondary Trigeminal Neurons. Molecules, 30(5), 1019. https://doi.org/10.3390/molecules30051019