

Autophagy-Enhancing Properties of Hedyotis diffusa Extracts in HaCaT Keratinocytes: Potential as an Anti-Photoaging Cosmetic Ingredient

 , ,
, ,  , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
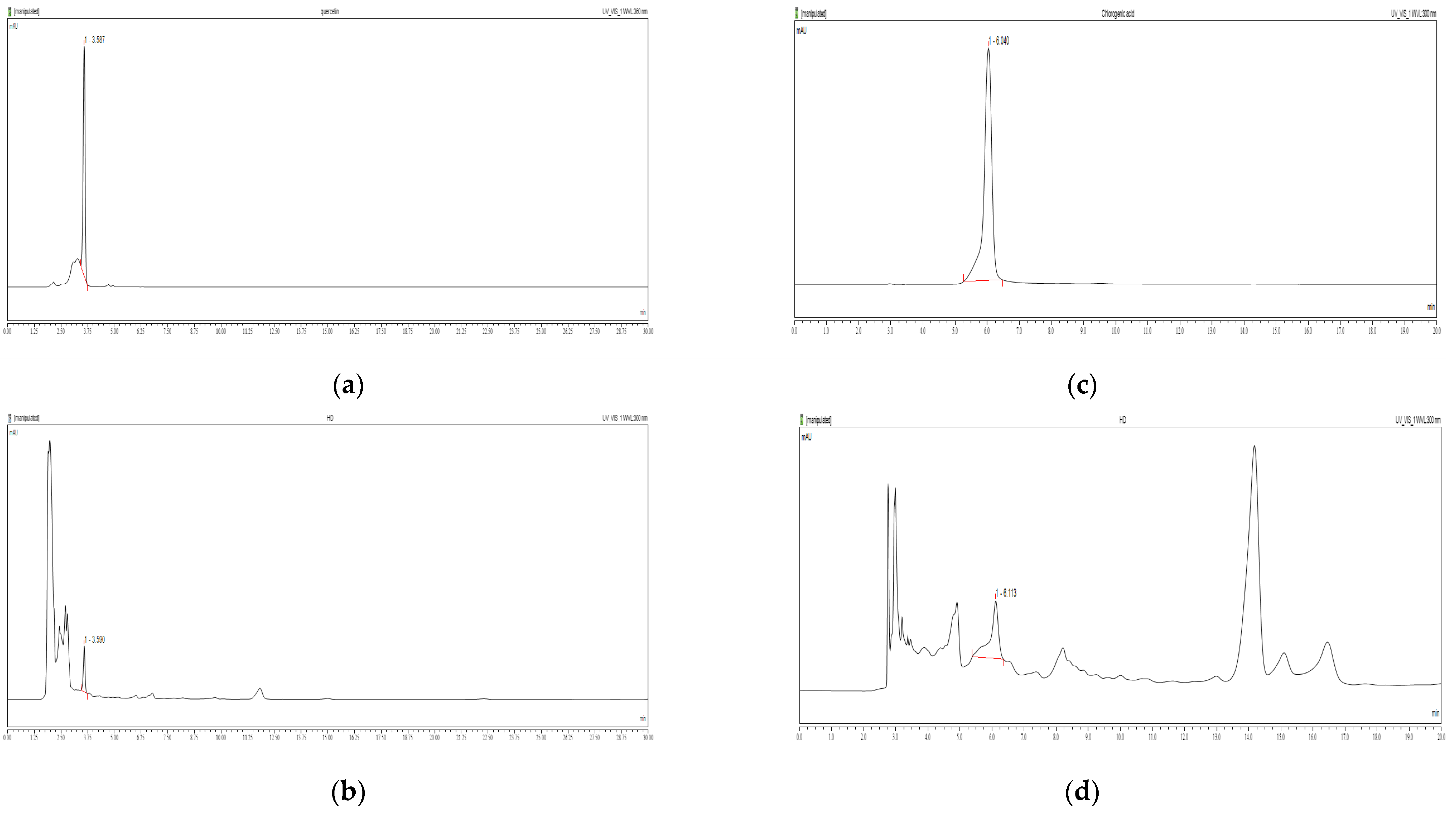
2.1. Chemical Contents of HD
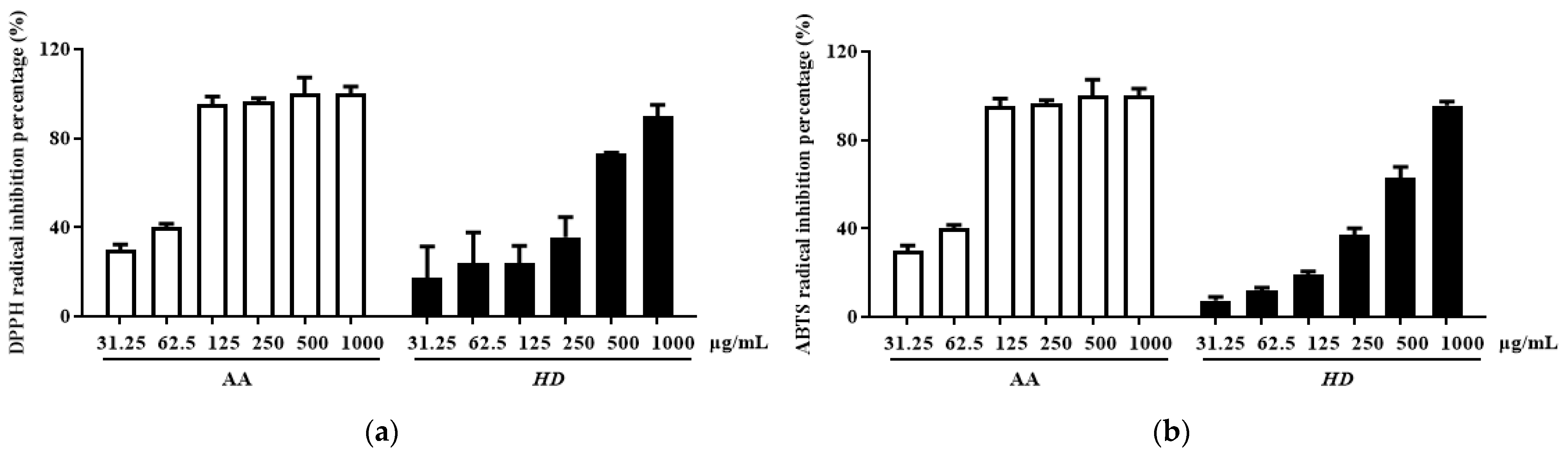
2.2. Antioxidant Effect of HD
2.3. Effects of HD on ROS Levels in HaCaT Keratinocytes Post-UVB Treatment
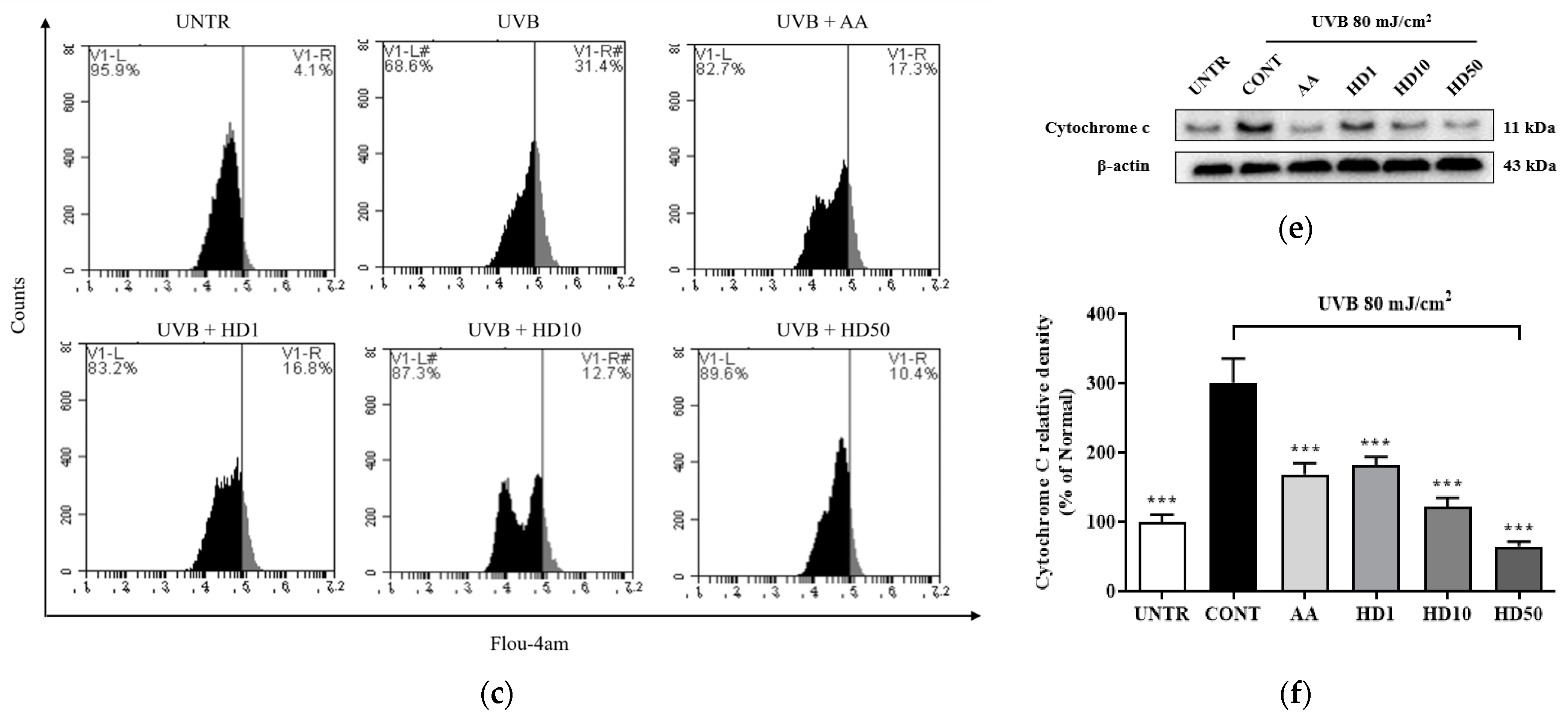
2.4. Effects of HD on Mitochondrial Membrane Function in HaCaT Keratinocytes Post-UVB Treatment
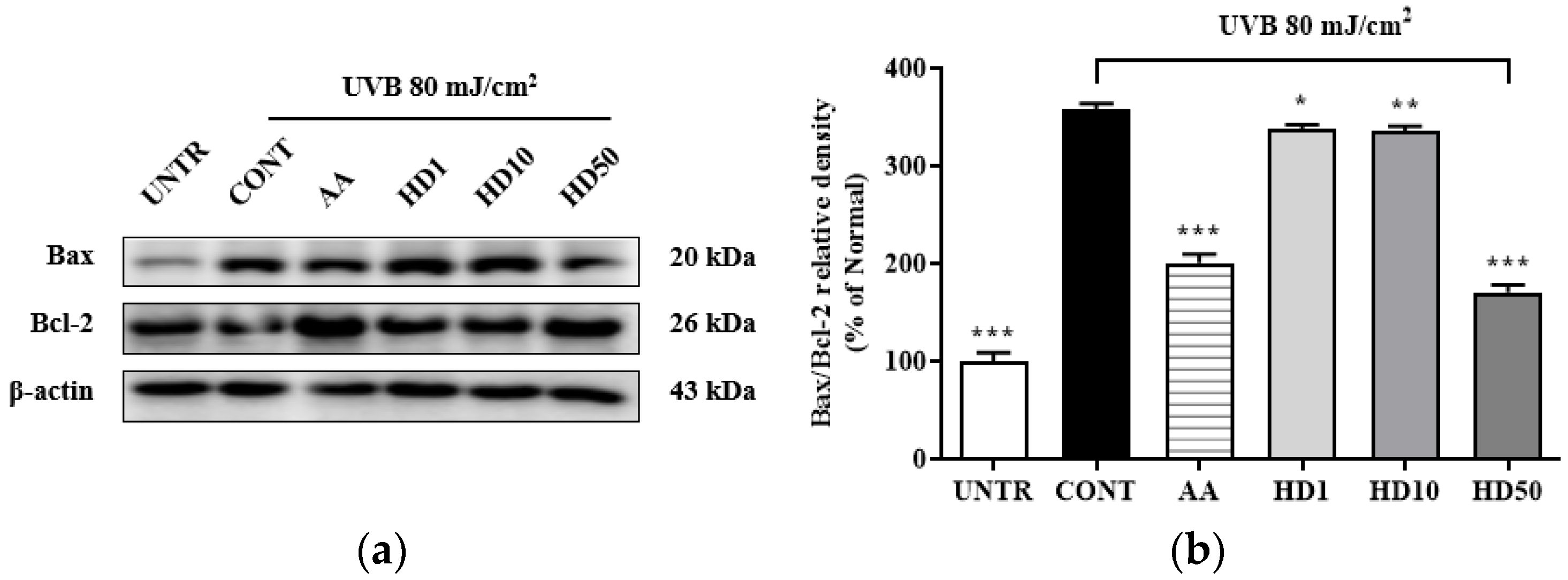
2.5. Effects of HD on the Bax/Bcl-2 Ratio in HaCaT Keratinocytes Post-UVB Treatment
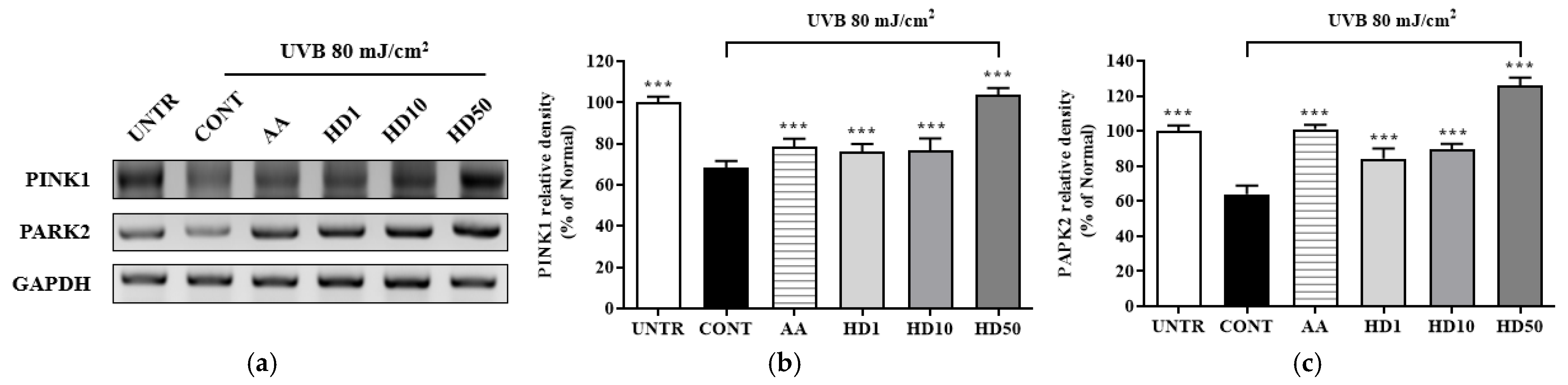
2.6. Effects of HD on the PINK/PARK2 and PI3K/AKT/mTOR Signaling Pathway in HaCaT Keratinocytes Post-UVB Treatment
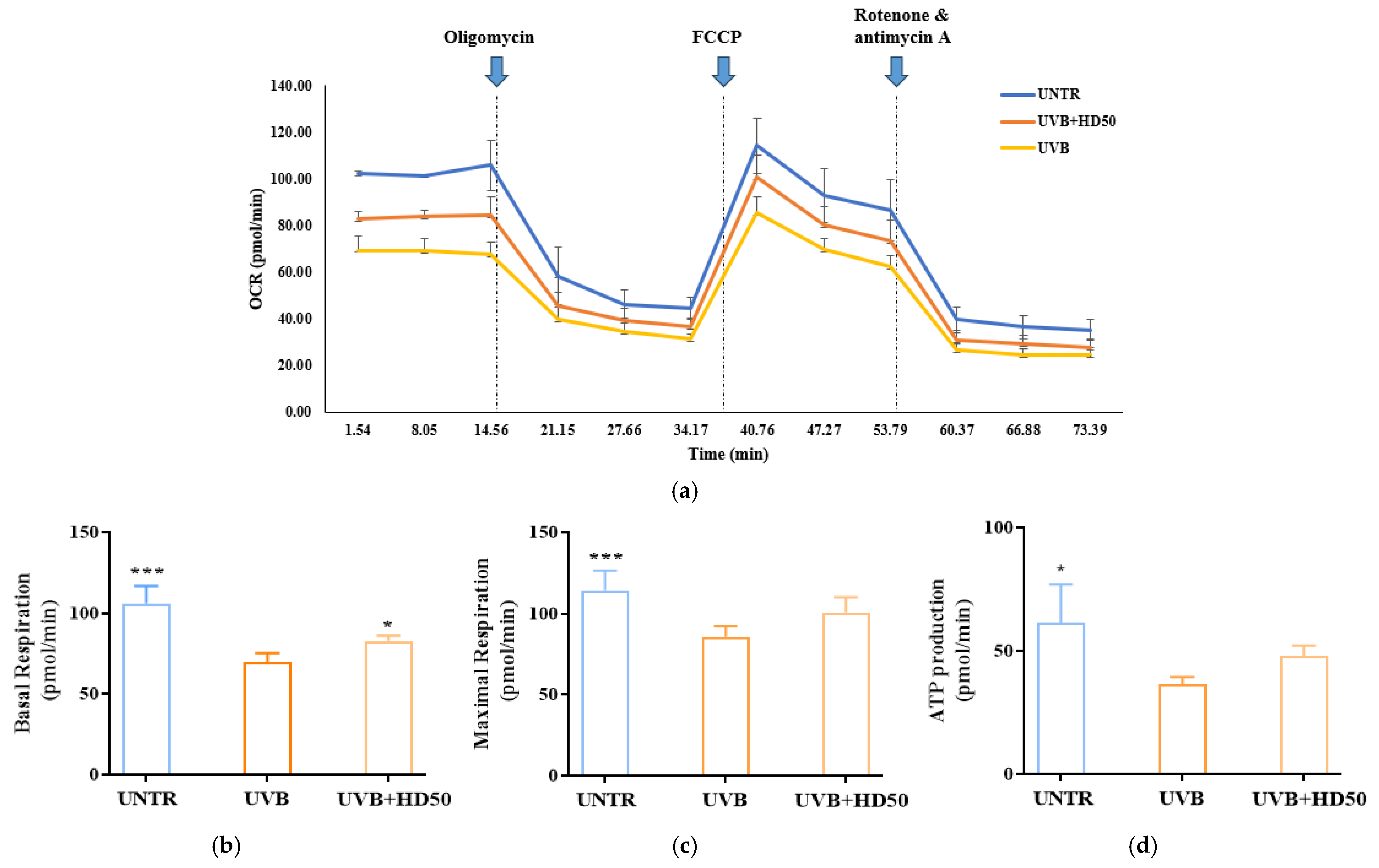
2.7. Effect of HD on Oxygen Consumption Rate (OCR) in HaCaT Keratinocytes Post-UVB Treatment
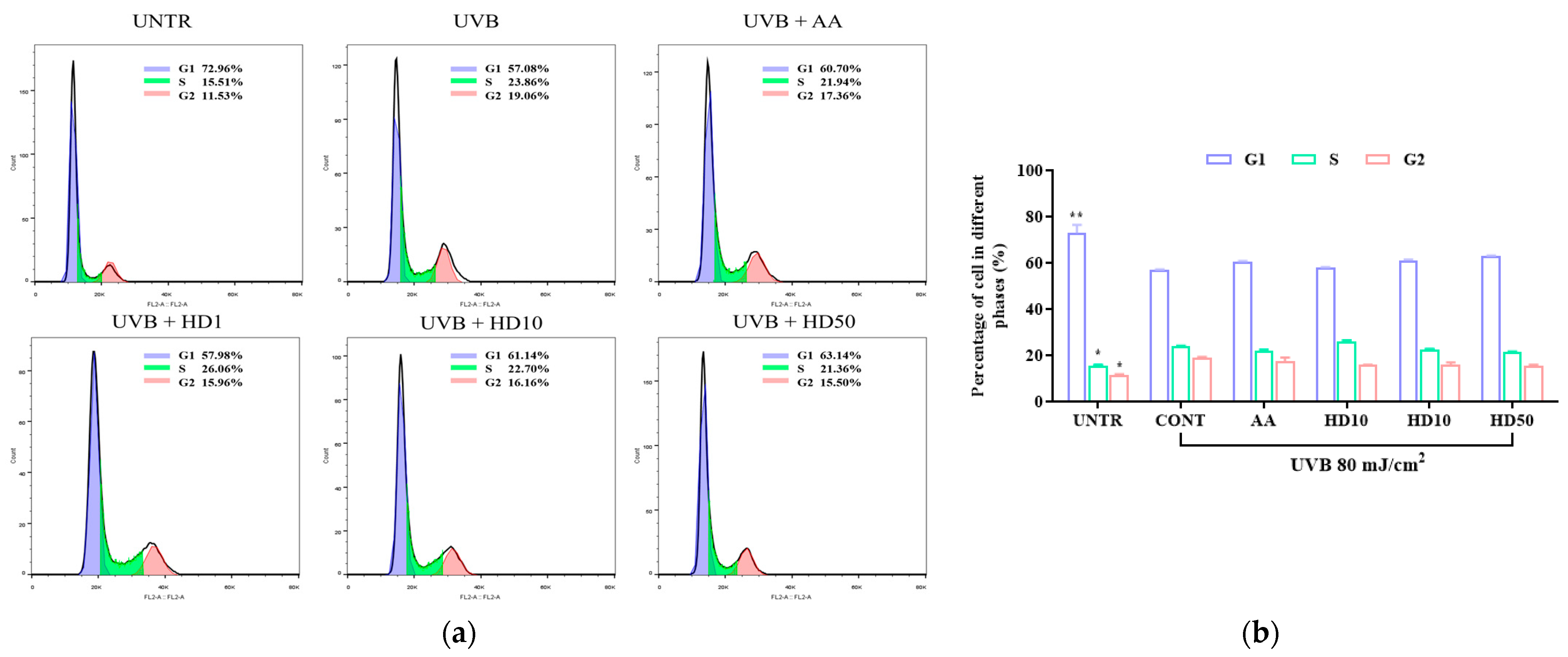
2.8. Effects of HD on HaCaT Keratinocytes Cycle Post-UVB Treatment
2.9. Effects of HD on Apoptosis-Related in HaCaT Keratinocytes Post-UVB Treatment
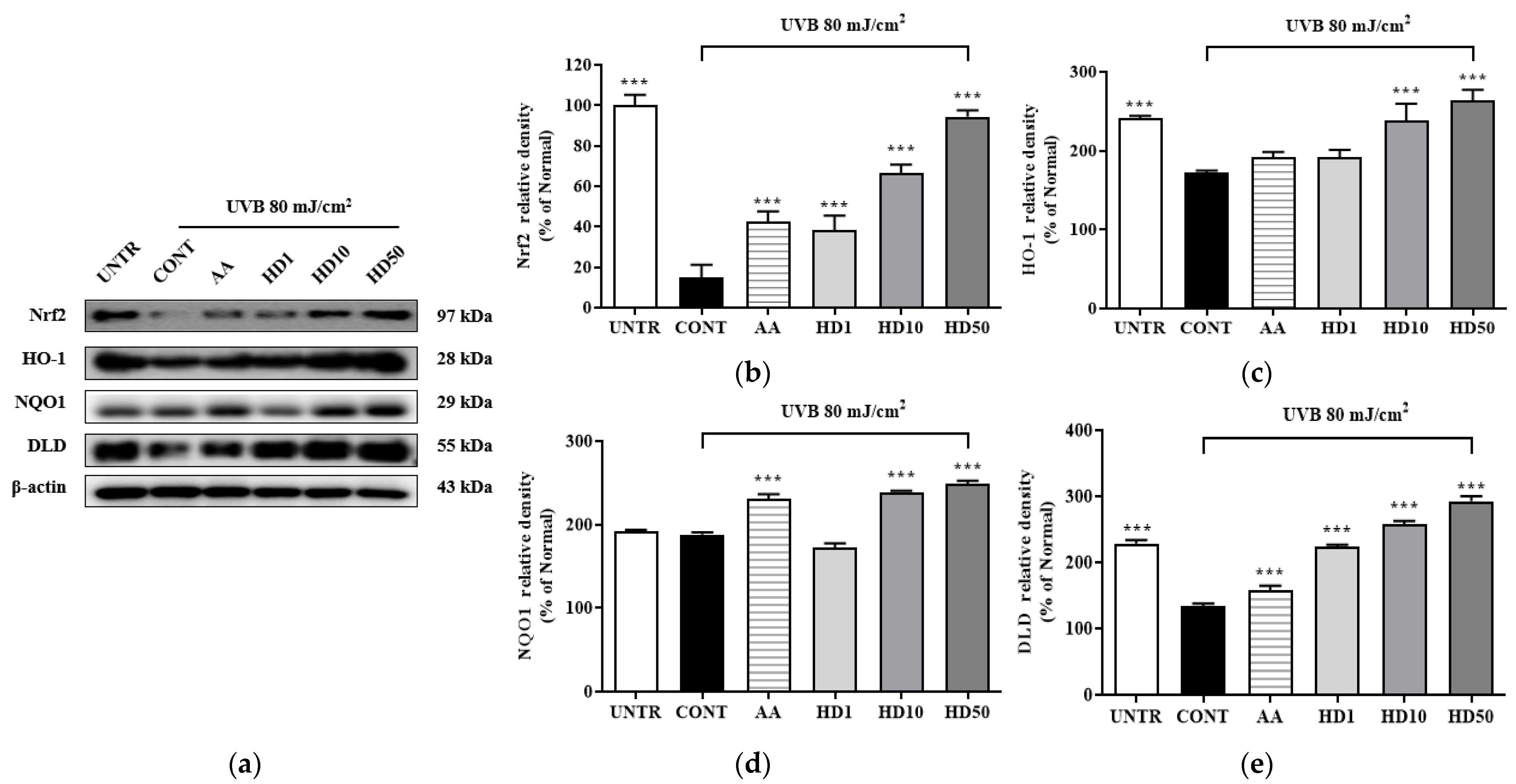
2.10. Effects of HD on the Nrf2/ARE Signaling Pathway in HaCaT Keratinocytes Post-UVB Treatment
3. Discussion
4. Materials and Methods
4.1. HD Extractions
4.2. HPLC
4.3. Measurement of Antioxidant Capacity
4.4. Cell Culture and Photoaging Model Establishment
4.5. Cytotoxicity
4.6. FACs
4.7. Measurement of the Cellular OCR
4.8. RT-PCR
4.9. Western Blotting
4.10. Statistical Methods and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, J.H.; Hanft, V.N.; Kang, S. Aging and Photoaging. J. Am. Acad. Dermatol. 2003, 49, 690–697. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV Light Touches the Brain and Endocrine System Through Skin, and Why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef]
- Kim, H.; Zheng, Q.; Oh, S.; Zheng, S.; Kim, M.; Yi, T.-H. Anti-Photoaging Effect of Jawoongo via Regulating Nrf2/ARE and TGF-β/Smad Signaling in In Vitro Photoaging Model. Appl. Sci. 2023, 13, 10425. [Google Scholar] [CrossRef]
- Umar, S.A.; Tasduq, S.A. Integrating DNA Damage Response and Autophagy Signalling Axis in Ultraviolet-B Induced Skin Photo-Damage: A Positive Association in Protecting Cells against Genotoxic Stress. RSC Adv. 2020, 10, 36317–36336. [Google Scholar] [CrossRef]
- Song, X.; Narzt, M.S.; Nagelreiter, I.M.; Hohensinner, P.; Terlecki-Zaniewicz, L.; Tschachler, E.; Grillari, J.; Gruber, F. Autophagy Deficient Keratinocytes Display Increased DNA Damage, Senescence and Aberrant Lipid Composition after Oxidative Stress in Vitro and in Vivo. Redox Biol. 2017, 11, 219–230. [Google Scholar] [CrossRef]
- Wang, M.; Charareh, P.; Lei, X.; Zhong, J.L. Autophagy: Multiple Mechanisms to Protect Skin from Ultraviolet Radiation-Driven Photoaging. Oxidative Med. Cell. Longev. 2019, 2019, 8135985. [Google Scholar] [CrossRef]
- Tang, Z.; Tong, X.; Huang, J.; Liu, L.; Wang, D.; Yang, S. Research Progress of Keratinocyte-programmed Cell Death in UV-induced Skin Photodamage. Photoderm. Photoimm. Photomed. 2021, 37, 442–448. [Google Scholar] [CrossRef]
- Ma, J.; Teng, Y.; Huang, Y.; Tao, X.; Fan, Y. Autophagy Plays an Essential Role in Ultraviolet Radiation-Driven Skin Photoaging. Front. Pharmacol. 2022, 13, 864331. [Google Scholar] [CrossRef]
- Slominski, R.M.; Chen, J.Y.; Raman, C.; Slominski, A.T. Photo-Neuro-Immuno-Endocrinology: How the Ultraviolet Radiation Regulates the Body, Brain, and Immune System. Proc. Natl. Acad. Sci. USA 2024, 121, e2308374121. [Google Scholar] [CrossRef]
- He, H.; Xiong, L.; Jian, L.; Li, L.; Wu, Y.; Qiao, S. Role of Mitochondria on UV-Induced Skin Damage and Molecular Mechanisms of Active Chemical Compounds Targeting Mitochondria. J. Photochem. Photobiol. B Biol. 2022, 232, 112464. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, X.; Hao, D.; Zhou, M.; Li, X.; He, G.; Jiang, X. Insights into Autophagy Machinery in Cells Related to Skin Diseases and Strategies for Therapeutic Modulation. Biomed. Pharmacother. 2019, 113, 108775. [Google Scholar] [CrossRef]
- Vikram, A.; Patel, S.K.; Singh, A.; Pathania, D.; Ray, R.S.; Upadhyay, A.K.; Dwivedi, A. Natural Autophagy Activators: A Promising Strategy for Combating Photoaging. Phytomedicine 2024, 132, 155508. [Google Scholar] [CrossRef]
- Cavinato, M.; Martic, I.; Wedel, S.; Pittl, A.; Koziel, R.; Weinmmüllner, R.; Schosserer, M.; Jenewein, B.; Bobbili, M.R.; Arcalis, E.; et al. Elimination of Damaged Mitochondria during UVB-induced Senescence Is Orchestrated by NIX-dependent Mitophagy. Aging Cell 2024, 23, e14186. [Google Scholar] [CrossRef]
- Oblong, J.E.; Bowman, A.; Rovito, H.A.; Jarrold, B.B.; Sherrill, J.D.; Black, M.R.; Nelson, G.; Kimball, A.B.; Birch-Machin, M.A. Metabolic Dysfunction in Human Skin: Restoration of Mitochondrial Integrity and Metabolic Output by Nicotinamide (Niacinamide) in Primary Dermal Fibroblasts from Older Aged Donors. Aging Cell 2020, 19, e13248. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Pan, Y.; Huang, P.; Wang, C. Autophagy Participates in Isoliquiritigenin–Induced Melanin Degradation in Human Epidermal Keratinocytes through PI3K/AKT/mTOR Signaling. Biomed. Pharmacother. 2018, 97, 248–254. [Google Scholar] [CrossRef]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR Pathway by Apigenin Induces Apoptosis and Autophagy in Hepatocellular Carcinoma Cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef]
- Heras-Sandoval, D.; Pérez-Rojas, J.M.; Hernández-Damián, J.; Pedraza-Chaverri, J. The Role of PI3K/AKT/mTOR Pathway in the Modulation of Autophagy and the Clearance of Protein Aggregates in Neurodegeneration. Cell. Signal. 2014, 26, 2694–2701. [Google Scholar] [CrossRef]
- Zhai, C.; Cheng, J.; Mujahid, H.; Wang, H.; Kong, J.; Yin, Y.; Li, J.; Zhang, Y.; Ji, X.; Chen, W. Selective Inhibition of PI3K/Akt/mTOR Signaling Pathway Regulates Autophagy of Macrophage and Vulnerability of Atherosclerotic Plaque. PLoS ONE 2014, 9, e90563. [Google Scholar] [CrossRef]
- Yang, H.-L.; Chen, S.-J.; Yeh, J.-T.; Vadivalagan, C.; Chiu, J.-H.; Hseu, J.-H.; Hseu, Y.-C. The Anti-Melanogenesis, Anti-Photoaging, and Anti-Inflammation of Coenzyme Q0, a Major Quinone Derivative from Antrodia Camphorata, through Antioxidant Nrf2 Signaling Pathways in UVA/B-Irradiated Keratinocytes. J. Funct. Foods 2024, 116, 106206. [Google Scholar] [CrossRef]
- Chaiprasongsuk, A.; Panich, U. Role of Phytochemicals in Skin Photoprotection via Regulation of Nrf2. Front. Pharmacol. 2022, 13, 823881. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, Y.; Hong, B.; Ma, L.; Zhao, Y.; Zhang, S.; Sun, S.; Ding, Q.; Wang, Y.; Liu, W.; et al. Dihydroquercetin Composite Nanofibrous Membrane Prevents UVA Radiation-Mediated Inflammation, Apoptosis and Oxidative Stress by Modulating MAPKs/Nrf2 Signaling in Human Epidermal Keratinocytes. Biomed. Pharmacother. 2022, 155, 113727. [Google Scholar] [CrossRef]
- Liu, C.; Vojnovic, D.; Kochevar, I.E.; Jurkunas, U.V. UV-A Irradiation Activates Nrf2-Regulated Antioxidant Defense and Induces P53/Caspase3-Dependent Apoptosis in Corneal Endothelial Cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2319. [Google Scholar] [CrossRef]
- Liu, W.; Yan, F.; Xu, Z.; Chen, Q.; Ren, J.; Wang, Q.; Chen, L.; Ying, J.; Liu, Z.; Zhao, J.; et al. Urolithin A Protects Human Dermal Fibroblasts from UVA-Induced Photoaging through NRF2 Activation and Mitophagy. J. Photochem. Photobiol. B Biol. 2022, 232, 112462. [Google Scholar] [CrossRef]
- Lu, F.; Zhou, Q.; Liang, M.; Liang, H.; Yu, Y.; Li, Y.; Zhang, Y.; Lu, L.; Zheng, Y.; Hao, J.; et al. α-Arbutin Ameliorates UVA-Induced Photoaging through Regulation of the SIRT3/PGC-1α Pathway. Front. Pharmacol. 2024, 15, 1413530. [Google Scholar] [CrossRef]
- Wang, K.-H.; Lin, R.-D.; Hsu, F.-L.; Huang, Y.-H.; Chang, H.-C.; Huang, C.-Y.; Lee, M.-H. Cosmetic Applications of Selected Traditional Chinese Herbal Medicines. J. Ethnopharmacol. 2006, 106, 353–359. [Google Scholar] [CrossRef]
- Zhang, R.; Ma, C.; Wei, Y.; Wang, X.; Jia, J.; Li, J.; Li, K.; Cao, G.; Yang, P. Isolation, Purification, Structural Characteristics, Pharmacological Activities, and Combined Action of Hedyotis Diffusa Polysaccharides: A Review. Int. J. Biol. Macromol. 2021, 183, 119–131. [Google Scholar] [CrossRef]
- Chen, R.; He, J.; Tong, X.; Tang, L.; Liu, M. The Hedyotis Diffusa Willd. (Rubiaceae): A Review on Phytochemistry, Pharmacology, Quality Control and Pharmacokinetics. Molecules 2016, 21, 710. [Google Scholar] [CrossRef]
- Park, J.H.; Whang, W.K. Bioassay-Guided Isolation of Anti-Alzheimer Active Components from the Aerial Parts of Hedyotis Diffusa and Simultaneous Analysis for Marker Compounds. Molecules 2020, 25, 5867. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Ahmed, T.; Nabavi, S.F. Quercetin and the Mitochondria: A Mechanistic View. Biotechnol. Adv. 2016, 34, 532–549. [Google Scholar] [CrossRef]
- Gao, L.-J.; Dai, Y.; Li, X.-Q.; Meng, S.; Zhong, Z.-Q.; Xu, S.-J. Chlorogenic Acid Enhances Autophagy by Upregulating Lysosomal Function to Protect against SH-SY5Y Cell Injury Induced by H2 O2. Exp. Ther. Med. 2021, 21, 426. [Google Scholar] [CrossRef]
- Yun, H.R.; Jo, Y.H.; Kim, J.; Shin, Y.; Kim, S.S.; Choi, T.G. Roles of Autophagy in Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 3289. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B. Autophagy in Mammalian Development and Differentiation. Nat. Cell Biol. 2010, 12, 823–830. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant Properties of Phenolic Compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Murphy, M.P. How Mitochondria Produce Reactive Oxygen Species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Chandra, D.; Liu, J.-W.; Tang, D.G. Early Mitochondrial Activation and Cytochrome c Up-Regulation during Apoptosis. J. Biol. Chem. 2002, 277, 50842–50854. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, Oxidative Stress and Cell Death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Duchen, M.R. Mitochondria and Calcium: From Cell Signalling to Cell Death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Richardson, A.P. The Mitochondrial Permeability Transition: A Current Perspective on Its Identity and Role in Ischaemia/Reperfusion Injury. J. Mol. Cell. Cardiol. 2015, 78, 129–141. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell Death. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 Protein Family: Opposing Activities That Mediate Cell Death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Li, C.; Zhu, Y.; Liu, W.; Xiang, W.; He, S.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ikejima, T. Impaired Mitophagy Causes Mitochondrial DNA Leakage and STING Activation in Ultraviolet B-Irradiated Human Keratinocytes HaCaT. Arch. Biochem. Biophys. 2023, 737, 109553. [Google Scholar] [CrossRef]
- Pickles, S.; Vigié, P.; Youle, R.J. Mitophagy and Quality Control Mechanisms in Mitochondrial Maintenance. Curr. Biol. 2018, 28, R170–R185. [Google Scholar] [CrossRef]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The Ubiquitin Kinase PINK1 Recruits Autophagy Receptors to Induce Mitophagy. Nature 2015, 524, 309–314. [Google Scholar] [CrossRef]
- Li, N.; Ragheb, K.; Lawler, G.; Sturgis, J.; Rajwa, B.; Melendez, J.A.; Robinson, J.P. Mitochondrial Complex I Inhibitor Rotenone Induces Apoptosis through Enhancing Mitochondrial Reactive Oxygen Species Production. J. Biol. Chem. 2003, 278, 8516–8525. [Google Scholar] [CrossRef]
- Jung, C.H.; Ro, S.-H.; Cao, J.; Otto, N.M.; Kim, D.-H. mTOR Regulation of Autophagy. FEBS Lett. 2010, 584, 1287–1295. [Google Scholar] [CrossRef]
- Brand, M.D.; Nicholls, D.G. Assessing Mitochondrial Dysfunction in Cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef]
- Nunnari, J.; Suomalainen, A. Mitochondria: In Sickness and in Health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef]
- Dranka, B.P.; Benavides, G.A.; Diers, A.R.; Giordano, S.; Zelickson, B.R.; Reily, C.; Zou, L.; Chatham, J.C.; Hill, B.G.; Zhang, J.; et al. Assessing Bioenergetic Function in Response to Oxidative Stress by Metabolic Profiling. Free Radic. Biol. Med. 2011, 51, 1621–1635. [Google Scholar] [CrossRef]
- Kim, G.; Han, D.-W.; Lee, J.H. The Cytoprotective Effects of Baicalein on H2O2-Induced ROS by Maintaining Mitochondrial Homeostasis and Cellular Tight Junction in HaCaT Keratinocytes. Antioxidants 2023, 12, 902. [Google Scholar] [CrossRef]
- Zhang, J.; Ney, P.A. Role of BNIP3 and NIX in Cell Death, Autophagy, and Mitophagy. Cell Death Differ. 2009, 16, 939–946. [Google Scholar] [CrossRef]
- Zhang, J.-A.; Luan, C.; Huang, D.; Ju, M.; Chen, K.; Gu, H. Induction of Autophagy by Baicalin Through the AMPK-mTOR Pathway Protects Human Skin Fibroblasts from Ultraviolet B Radiation-Induced Apoptosis. Drug Des. Dev. Ther. 2020, 14, 417–428. [Google Scholar] [CrossRef]
- Wang, A.S.; Dreesen, O. Biomarkers of Cellular Senescence and Skin Aging. Front. Genet. 2018, 9, 247. [Google Scholar] [CrossRef]
- Peng, X.; Ma, Y.; Yan, C.; Wei, X.; Zhang, L.; Jiang, H.; Ma, Y.; Zhang, S.; Xing, M.; Gao, Y. Mechanism, Formulation, and Efficacy Evaluation of Natural Products for Skin Pigmentation Treatment. Pharmaceutics 2024, 16, 1022. [Google Scholar] [CrossRef]
- Cerulli, A.; Masullo, M.; Montoro, P.; Piacente, S. Licorice (Glycyrrhiza glabra, G. uralensis, and G. inflata) and Their Constituents as Active Cosmeceutical Ingredients. Cosmetics 2022, 9, 7. [Google Scholar] [CrossRef]
- Chang, M.-J.; Huang, H.-C.; Chang, H.-C.; Chang, T.-M. Cosmetic Formulations Containing Lithospermum Erythrorhizon Root Extract Show Moisturizing Effects on Human Skin. Arch. Dermatol. Res. 2008, 300, 317–323. [Google Scholar] [CrossRef]
- Kang, J.; Cho, H.; Choi, H.; Lee, I. Anti-wrinkle Properties of Angelica gigas Nakai Root Extracts Using Mineral-rich Water. J. Cosmet. Dermatol. 2023, 22, 328–334. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in Skin Health, Aging, and Disease. Cell Death Dis. 2020, 11, 444. [Google Scholar] [CrossRef]
- Zhu, Y.; Xiang, W.; He, S.; San, Z.; Liu, W.; Wu, J.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; et al. Collagen I Protects Human Keratinocytes HaCaT against UVB Injury via Restoring PINK1/Parkin-Mediated Mitophagy. Arch. Biochem. Biophys. 2024, 753, 109905. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Youle, R.J.; Narendra, D.P. Mechanisms of Mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Yu, L.; Qiao, G.; Qin, D.; Law, B.Y.K.; Ren, F.; Wu, J.; Wu, A. Mitophagy and cGAS–STING Crosstalk in Neuroinflammation. Acta Pharm. Sin. B 2024, 14, 3327–3361. [Google Scholar] [CrossRef]
- Liang, H.; Ma, Z.; Zhong, W.; Liu, J.; Sugimoto, K.; Chen, H. Regulation of Mitophagy and Mitochondrial Function: Natural Compounds as Potential Therapeutic Strategies for Parkinson’s Disease. Phytother. Res. 2024, 38, 1838–1862. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and mTOR Regulate Autophagy through Direct Phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef]
- Yu, Q.; Zhang, R.; Li, T.; Yang, L.; Zhou, Z.; Hou, L.; Wu, W.; Zhao, R.; Chen, X.; Yao, Y.; et al. Correction to: Mitochondrial Hydrogen Peroxide Activates PTEN and Inactivates Akt Leading to Autophagy Inhibition-Dependent Cell Death in Neuronal Models of Parkinson’s Disease. Mol. Neurobiol. 2023, 60, 4166–4167. [Google Scholar] [CrossRef]
- Mao, Y.; Du, J.; Chen, X.; Al Mamun, A.; Cao, L.; Yang, Y.; Mubwandarikwa, J.; Zaeem, M.; Zhang, W.; Chen, Y.; et al. Maltol Promotes Mitophagy and Inhibits Oxidative Stress via the Nrf2/PINK1/Parkin Pathway after Spinal Cord Injury. Oxidative Med. Cell. Longev. 2022, 2022, 1337630. [Google Scholar] [CrossRef]
- Xiao, L.; Xu, X.; Zhang, F.; Wang, M.; Xu, Y.; Tang, D.; Wang, J.; Qin, Y.; Liu, Y.; Tang, C.; et al. The Mitochondria-Targeted Antioxidant MitoQ Ameliorated Tubular Injury Mediated by Mitophagy in Diabetic Kidney Disease via Nrf2/PINK1. Redox Biol. 2017, 11, 297–311. [Google Scholar] [CrossRef]
- Santos, A.C.; Morais, F.; Simões, A.; Pereira, I.; Sequeira, J.A.D.; Pereira-Silva, M.; Veiga, F.; Ribeiro, A. Nanotechnology for the Development of New Cosmetic Formulations. Expert. Opin. Drug Deliv. 2019, 16, 313–330. [Google Scholar] [CrossRef]
- Akhtar, N.; Singh, V.; Yusuf, M.; Khan, R.A. Non-Invasive Drug Delivery Technology: Development and Current Status of Transdermal Drug Delivery Devices, Techniques and Biomedical Applications. Biomed. Eng. Biomed. Tech. 2020, 65, 243–272. [Google Scholar] [CrossRef]
- Lee, S.; Oh, S.; Zheng, Q.; Zheng, S.; Kim, M.; Park, S.; Choi, W.; Yin, C.S.; Yi, T. Photoprotective Effects of Lithospermum erythrorhizon and Pueraria lobata Extracts on UVB-induced Photoaging: A Study on Skin Barrier Protection. Photoderm. Photoimm. Photomed. 2024, 40, e12950. [Google Scholar] [CrossRef]
- Kim, M.; Ha, L.-K.; Oh, S.; Fang, M.; Zheng, S.; Bellere, A.D.; Jeong, J.; Yi, T.-H. Antiphotoaging Effects of Damiana (Turnera diffusa) Leaves Extract via Regulation AP-1 and Nrf2/ARE Signaling Pathways. Plants 2022, 11, 1486. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Q.; Jin, X.; Nguyen, T.T.M.; Yi, E.-J.; Park, S.-J.; Yi, G.-S.; Yang, S.-J.; Yi, T.-H. Autophagy-Enhancing Properties of Hedyotis diffusa Extracts in HaCaT Keratinocytes: Potential as an Anti-Photoaging Cosmetic Ingredient. Molecules 2025, 30, 261. https://doi.org/10.3390/molecules30020261
Zheng Q, Jin X, Nguyen TTM, Yi E-J, Park S-J, Yi G-S, Yang S-J, Yi T-H. Autophagy-Enhancing Properties of Hedyotis diffusa Extracts in HaCaT Keratinocytes: Potential as an Anti-Photoaging Cosmetic Ingredient. Molecules. 2025; 30(2):261. https://doi.org/10.3390/molecules30020261
Chicago/Turabian StyleZheng, Qiwen, Xiangji Jin, Trang Thi Minh Nguyen, Eun-Ji Yi, Se-Jig Park, Gyeong-Seon Yi, Su-Jin Yang, and Tae-Hoo Yi. 2025. "Autophagy-Enhancing Properties of Hedyotis diffusa Extracts in HaCaT Keratinocytes: Potential as an Anti-Photoaging Cosmetic Ingredient" Molecules 30, no. 2: 261. https://doi.org/10.3390/molecules30020261
APA StyleZheng, Q., Jin, X., Nguyen, T. T. M., Yi, E.-J., Park, S.-J., Yi, G.-S., Yang, S.-J., & Yi, T.-H. (2025). Autophagy-Enhancing Properties of Hedyotis diffusa Extracts in HaCaT Keratinocytes: Potential as an Anti-Photoaging Cosmetic Ingredient. Molecules, 30(2), 261. https://doi.org/10.3390/molecules30020261