
Chrysin-Loaded Extracellular Vesicles Attenuate LPS-Induced Neuroinflammation in BV2 Microglial Cells In Vitro: A Novel Neuroprotective Strategy

 , , , , , ,
, , , , , ,  ,
,  , and
, and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
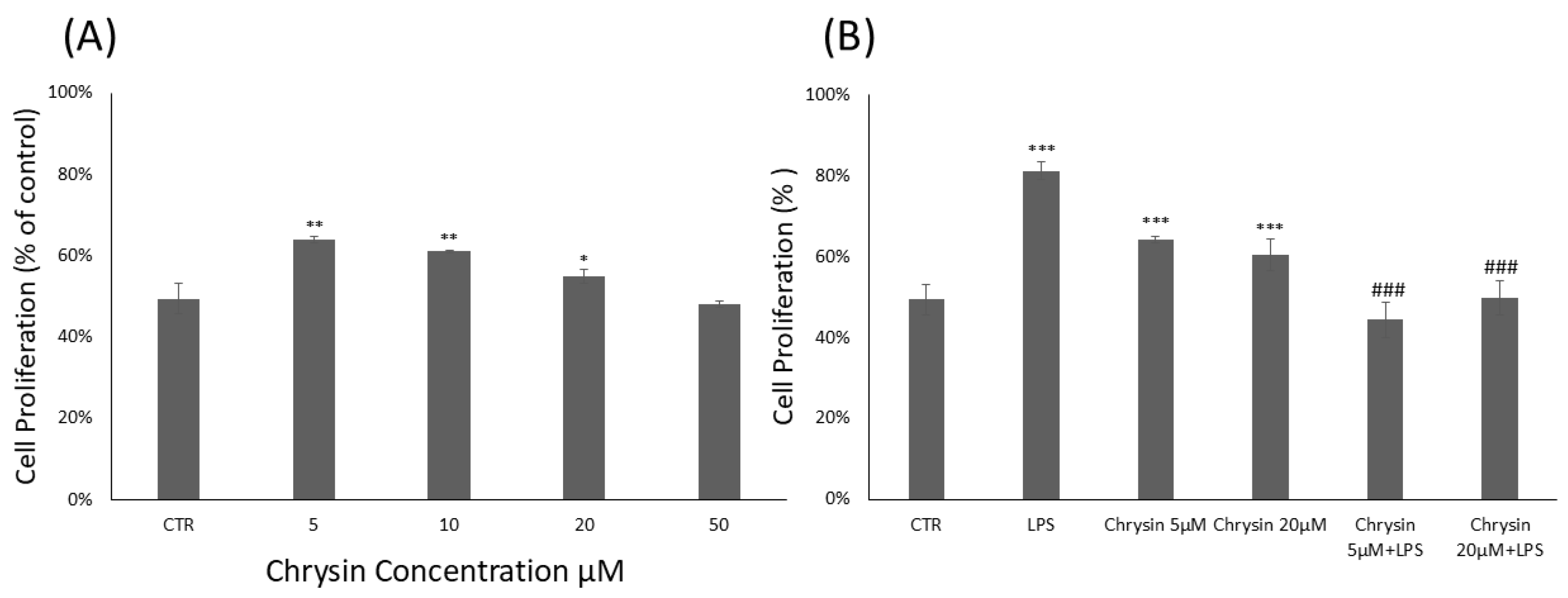
2.1. Chrysin Cytotoxicity and Microglial Proliferation Effects
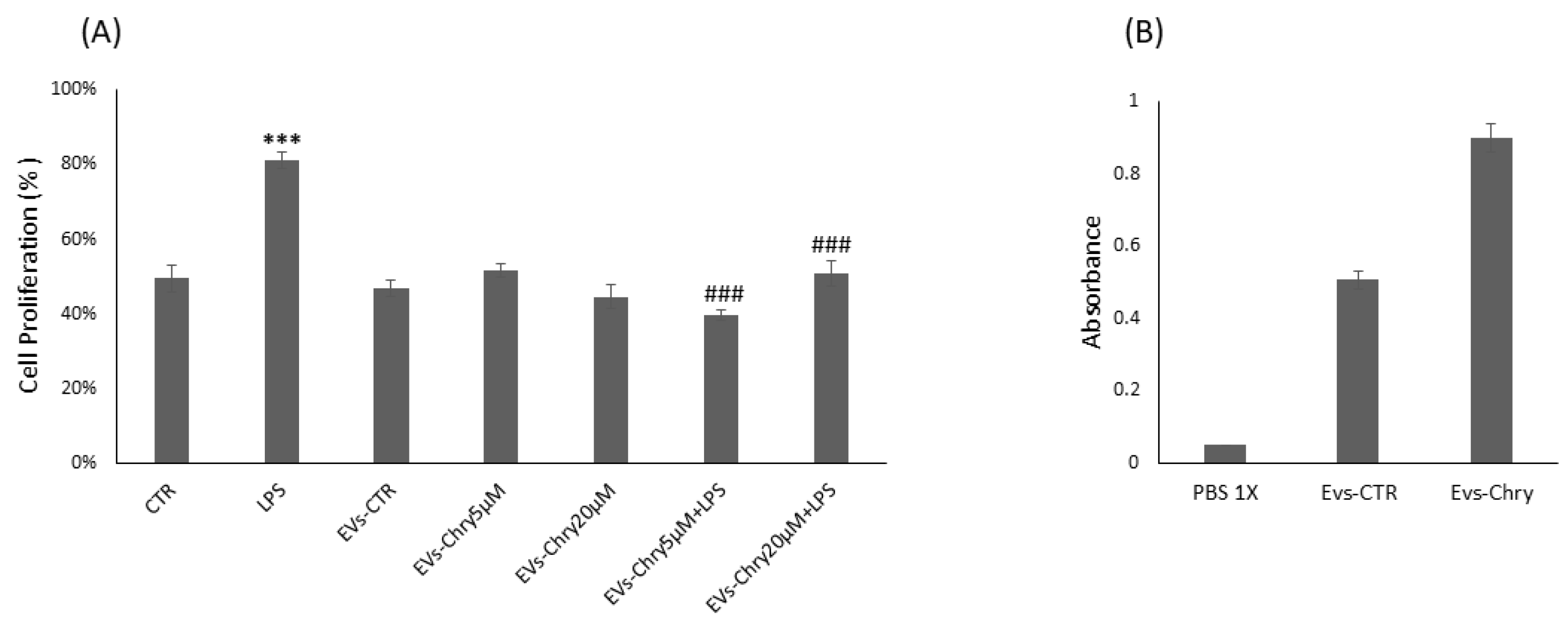
2.2. Isolation, Characterization and Cytotoxic Effect of Chrysin-Loaded EVs from Microglial Cells
2.3. BV-2 Cells Activated with LPS Show Anti-Inflammatory M2 State After EVs-Chry Stimulation
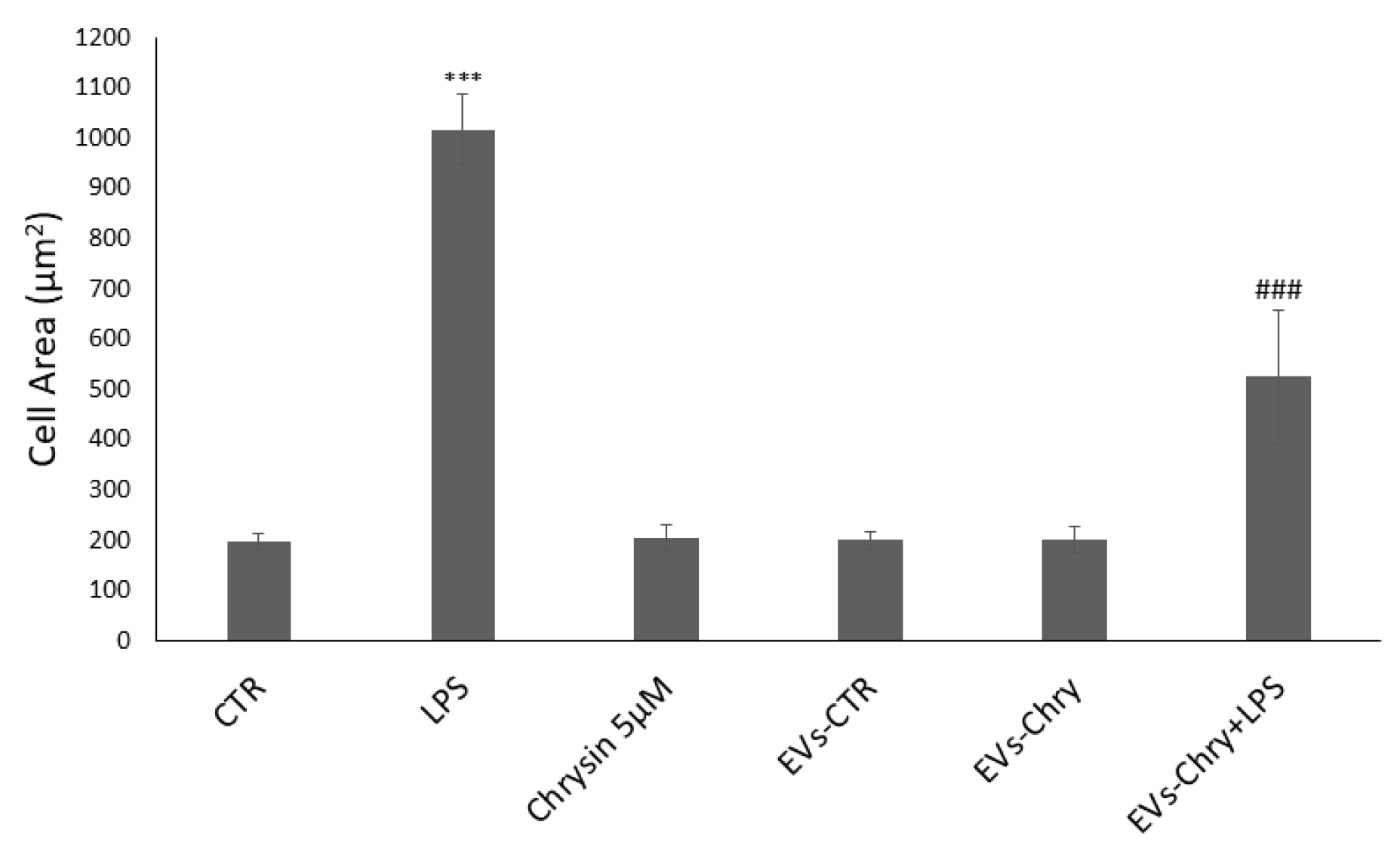
2.4. EVs-Chry Attenuate LPS-Induced Migration of BV2 Microglial Cells
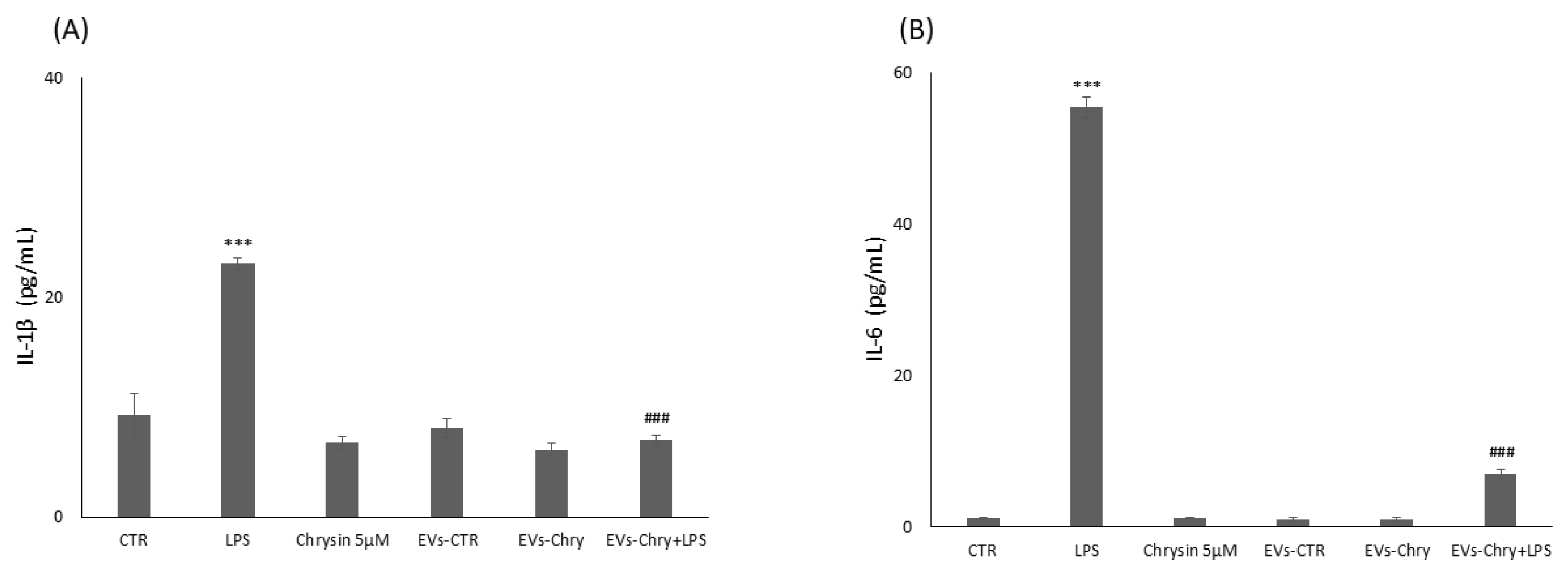
2.5. Suppression Effect of EVs-Chry on Expression of Pro-Inflammatory Molecules in BV2 Cells
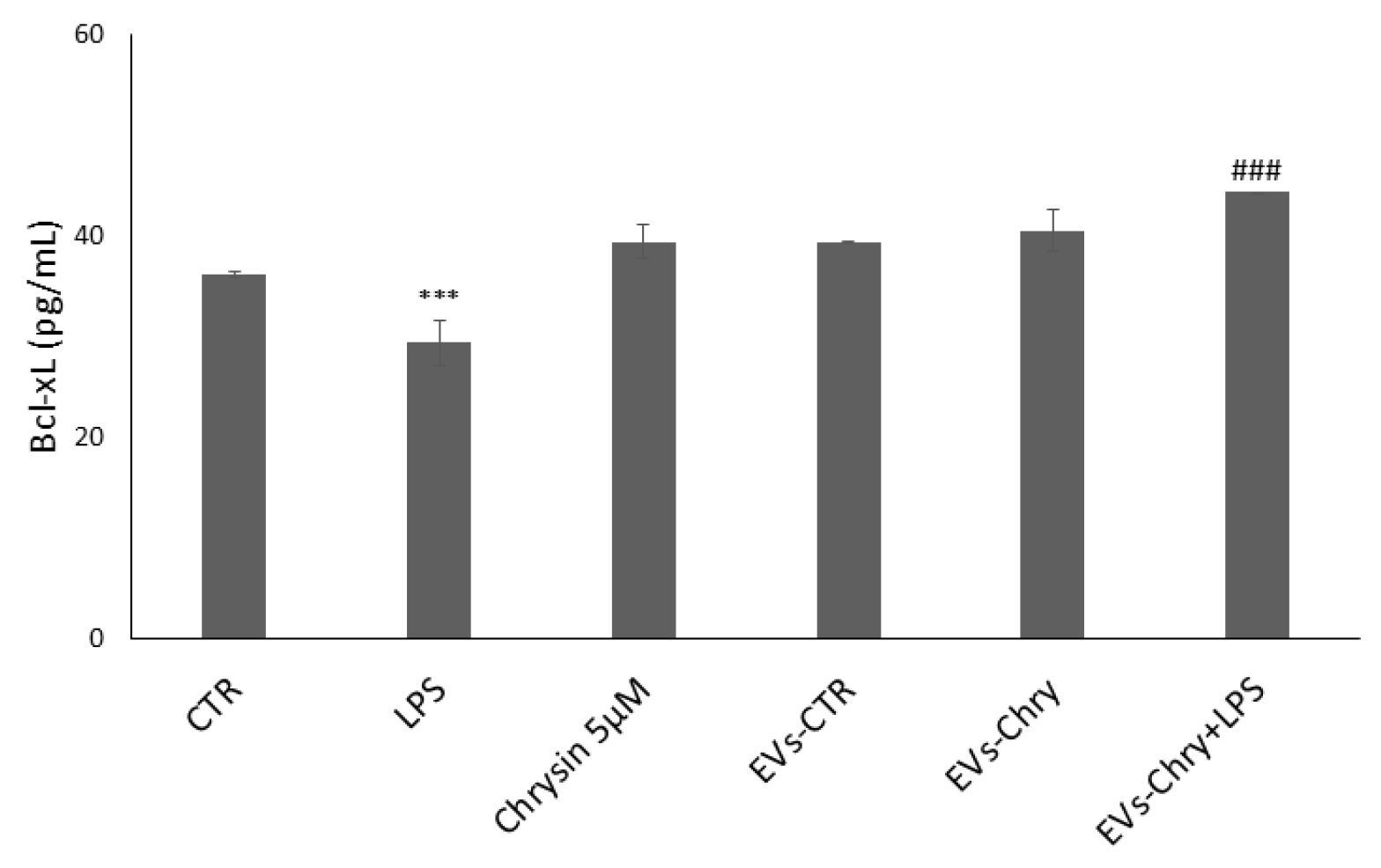
2.6. EVs-Chry Stimulate the Increase in the Anti-Apoptotic Protein Bcl-xL in BV-2 Cells
3. Discussion
4. Materials and Methods
4.1. Microglial Cell Culture
4.2. EVs Production and Isolation
4.3. EVs Analysis
4.4. Crhysin Solution Preparation
4.5. Cell Proliferation Assay
4.6. Cell Morphology Analysis
4.7. Wound Healing Assay
4.8. EISA Quantification of Cytokines
4.9. Protein Extraction and Western Blot Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rauf, A.; Badoni, H.; Abu-Izneid, T.; Olatunde, A.; Rahman, M.; Painuli, S.; Semwal, P.; Wilairatana, P.; Mubarak, M.S. Neuroinflammatory Markers: Key Indicators in the Pathology of Neurodegenerative Diseases. Molecules 2022, 27, 3194. [Google Scholar] [CrossRef]
- Gong, P.; Jia, H.-Y.; Li, R.; Ma, Z.; Si, M.; Qian, C.; Zhu, F.-Q.; Sheng-Yong, L. Downregulation of Nogo-B Ameliorates Cer-ebral Ischemia/Reperfusion Injury in Mice through Regulating Microglia Polarization via TLR4/NF-kappaB Pathway. Neurochem. Int. 2023, 167, 105553. [Google Scholar] [CrossRef]
- Wang, M.; Pan, W.; Xu, Y.; Zhang, J.; Wan, J.; Jiang, H. Microglia-Mediated Neuroinflammation: A Potential Target for the Treatment of Cardiovascular Diseases. J. Inflamm. Res. 2022, 15, 3083–3094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Miao, L.; Peng, Q.; Fan, X.; Song, W.; Yang, B.; Zhang, P.; Liu, G.; Liu, J. Parthenolide modulates cerebral ischemia-induced microglial polarization and alleviates neuroinflammatory injury via the RhoA/ROCK pathway. Phytomedicine 2022, 105, 154373. [Google Scholar] [CrossRef]
- 5Ajami, B.; Bennett, J.L.; Krieger, C.; Tetzlaff, W.; Rossi, F.M.V. Local self-renewal can sustain CNS microglia maintenance and function throughout adult life. Nat. Neurosci. 2007, 10, 1538–1543. [Google Scholar] [CrossRef] [PubMed]
- Dubbelaar, M.L.; Kracht, L.; Eggen, B.J.L.; Boddeke, E.W.G.M. The Kaleidoscope of Microglial Phenotypes. Front. Immunol. 2018, 9, 1753. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma in Vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef]
- Von Bernhardi, R.; Heredia, F.; Salgado, N.; Muñoz, P. Microglia Function in the Normal Brain. Adv. Exp. Med. Biol. 2016, 949, 67–92. [Google Scholar]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef]
- Cronk, J.C.; Kipnis, J. Microglia–the Brain’s Busy Bees. F1000Prime Rep. 2013, 5, 53. [Google Scholar] [CrossRef]
- Spiers, J.G.; Vassileff, N.; Hill, A.F. Neuroinflammatory Modulation of Extracellular Vesicle Biogenesis and Cargo Loading. NeuroMolecular Med. 2022, 24, 385–391. [Google Scholar] [CrossRef]
- Bourgognon, J.-M.; Spiers, J.G.; Robinson, S.W.; Scheiblich, H.; Glynn, P.; Ortori, C.; Bradley, S.J.; Tobin, A.B.; Steinert, J.R. Inhibition of neuroinflammatory nitric oxide signaling suppresses glycation and prevents neuronal dysfunction in mouse prion disease. Proc. Natl. Acad. Sci. USA 2021, 118, e2009579118. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [PubMed]
- Scalavino, V.; Liso, M.; Cavalcanti, E.; Gigante, I.; Lippolis, A.; Mastronardi, M.; Chieppa, M.; Serino, G. miR-369-3p modulates inducible nitric oxide synthase and is involved in regulation of chronic inflammatory response. Sci. Rep. 2020, 10, 15942. [Google Scholar] [CrossRef] [PubMed]
- Vassileff, N.; Cheng, L.; Hill, A.F. Extracellular vesicles–propagators of neuropathology and sources of potential biomarkers and therapeutics for neurodegenerative diseases. J. Cell Sci. 2020, 133, jcs243139. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Stoica, B.A.; Loane, D.J.; Yang, M.; Abulwerdi, G.; Khan, N.; Kumar, A.; Thom, S.R.; Faden, A.I. Microglial-derived microparticles mediate neuroinflammation after traumatic brain injury. J. Neuroinflam. 2017, 14, 1–17. [Google Scholar] [CrossRef]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and mi-croRNA Dysregulation. Front. Cell Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef]
- D’EGidio, F.; Castelli, V.; D’ANgelo, M.; Ammannito, F.; Quintiliani, M.; Cimini, A. Brain incoming call from glia during neuroinflammation: Roles of extracellular vesicles. Neurobiol. Dis. 2024, 201, 106663. [Google Scholar] [CrossRef]
- Yan, B.; Liao, P.; Liu, Y.; Han, Z.; Wang, C.; Chen, F.; Lei, P. Therapeutic potential of microglia-derived extracellular vesicles in ischemic stroke. Int. Immunopharmacol. 2024, 139, 112712. [Google Scholar] [CrossRef]
- Wang, A.-P.; Tian, Y.; Zhang, W.; Tian, T.; Gong, S.-X.; Huang, W.-Q.; Zhou, Q.-Y. Microglia-associated neuroinflammation is a potential therapeutic target for ischemic stroke. Neural Regen. Res. 2021, 16, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Kopustinskiene, D.M.; Simal-Gandara, J.; Bernatoniene, J.; Samarghandian, S. An Up-dated Review on the Versatile Role of Chrysin in Neurological Diseases: Chemistry, Pharmacology, and Drug Delivery Ap-proaches. Biomed. Pharmacother. 2021, 141, 111906. [Google Scholar] [CrossRef] [PubMed]
- Hadjmohammadi, M.R.; Nazari, S.S.S.J. Separation optimization of quercetin, hesperetin and chrysin in honey by micellar liquid chromatography and experimental design. J. Sep. Sci. 2010, 33, 3144–3151. [Google Scholar] [CrossRef]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Samarghandian, S. Molecular Mechanism-Based Therapeutic Properties of Honey. Biomed. Pharmacother. 2020, 130, 110590. [Google Scholar] [CrossRef]
- Al-Hatamleh, M.A.I.; Boer, J.C.; Wilson, K.L.; Plebanski, M.; Mohamud, R.; Mustafa, M.Z. Antioxidant-Based Medicinal Properties of Stingless Bee Products: Recent Progress and Future Directions. Biomolecules 2020, 10, 923. [Google Scholar] [CrossRef]
- Dinda, B.; SilSarma, I.; Dinda, M.; Rudrapaul, P. Oroxylum indicum (L.) Kurz, an important Asian traditional medicine: From traditional uses to scientific data for its commercial exploitation. J. Ethnopharmacol. 2015, 161, 255–278. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Yanni, A.E.; Koutrotsios, G.; Aloupi, M. Bioactive microconstituents and antioxidant properties of wild edible mushrooms from the island of Lesvos, Greece. Food Chem. Toxicol. 2013, 55, 378–385. [Google Scholar] [CrossRef]
- Kamat, S.; Kumari, M.; Sajna, K.V.; Jayabaskaran, C. Endophytic fungus, Chaetomium globosum, associated with marine green alga, a new source of Chrysin. Sci. Rep. 2020, 10, 18726. [Google Scholar] [CrossRef]
- Samarghandian, S.; Afshari, J.T.; Davoodi, S. Chrysin reduces proliferation and induces apoptosis in the human prostate cancer cell line pc-3. Clinics 2011, 66, 1073–1079. [Google Scholar] [CrossRef]
- Moghadam, E.R.; Ang, H.L.; Asnaf, S.E.; Zabolian, A.; Saleki, H.; Yavari, M.; Esmaeili, H.; Zarrabi, A.; Ashrafizadeh, M.; Kumar, A.P. Broad-Spectrum Preclinical Antitumor Activity of Chrysin: Current Trends and Future Perspectives. Biomolecules 2020, 10, 1374. [Google Scholar] [CrossRef]
- Saleem, S.; Banerjee, R.; Kannan, R.R. Chrysin-Loaded Chitosan Nanoparticle-Mediated Neuroprotection in Aβ1–42-Induced Neurodegenerative Conditions in Zebrafish. ACS Chem. Neurosci. 2022, 13, 2017–2034. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Abo Elseoud, O.G.; Mohamedy, M.H.; Amer, M.M.; Mohamed, Y.Y.; Elmansy, S.A.; Kadry, M.M.; Attia, A.A.; Fanous, R.A.; Kamel, M.S.; et al. Nose-to-Brain Delivery of Chrysin Transfersomal and Composite Vesicles in Doxorubi-cin-Induced Cognitive Impairment in Rats: Insights on Formulation, Oxidative Stress and TLR4/NF-kB/NLRP3 Pathways. Neuropharmacology 2021, 197, 108738. [Google Scholar] [CrossRef]
- Xiao, T.; Pan, M.; Wang, Y.; Huang, Y.; Tsunoda, M.; Zhang, Y.; Wang, R.; Hu, W.; Yang, H.; Li, L.-S.; et al. In vitro blood brain barrier permeability study of four main active ingredients from Alpiniae oxyphyllae fructus. J. Pharm. Biomed. Anal. 2023, 235, 115637. [Google Scholar] [CrossRef]
- Mishra, A.; Mishra, P.S.; Bandopadhyay, R.; Khurana, N.; Angelopoulou, E.; Paudel, Y.N.; Piperi, C. Neuroprotective Poten-tial of Chrysin: Mechanistic Insights and Therapeutic Potential for Neurological Disorders. Molecules 2021, 26, 6456. [Google Scholar] [CrossRef] [PubMed]
- Komath, S.; Garg, A.; Wahajuddin, M.; Din, W. Development and evaluation of Chrysin-Phospholipid complex loaded solid lipid nanoparticles–storage stability and in vitro anti-cancer activity. J. Microencapsul. 2018, 35, 600–617. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yang, X.; Li, T. Natural flavonoids from herbs and nutraceuticals as ferroptosis inhibitors in central nervous system diseases: Current preclinical evidence and future perspectives. Front. Pharmacol. 2025, 16, 1570069. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, Z.; Wu, C.; Zhou, C.; Zhang, X.; Gu, T.; Yang, J.; Yang, H.; Zheng, E.; Xu, Z.; et al. Extracellular vesicle-encapsulated miR-21-5p in seminal plasma prevents sperm capacitation via Vinculin inhibition. Theriogenology 2022, 193, 103–113. [Google Scholar] [CrossRef]
- Danis, E.G.; Mogulkoc, R.; Baltaci, A.K. Flavonoids in Brain Ischemia-Reperfusion and their Effect on Kinases as Signaling Pathway Activity. CNS Neurol. Disord. Drug Targets 2025. [Google Scholar] [CrossRef]
- Gomes, A.R.; Sangani, N.B.; Fernandes, T.G.; Diogo, M.M.; Curfs, L.M.G.; Reutelingsperger, C.P. Extracellular Vesicles in CNS Developmental Disorders. Int. J. Mol. Sci. 2020, 21, 9428. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, D.; Zhou, H.; Tao, B.; Chang, L.; Liu, H.; Luo, H.; Wang, D.; Liu, W. A New Nanomaterial Based on Extracellular Vesicles Containing Chrysin-Induced Cell Apoptosis Through Let-7a in Tongue Squamous Cell Carcinoma. Front. Bioeng. Biotechnol. 2021, 9, 766380. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, L.; Bortolotto, V.C.; Ferreira, L.M.; Sari, M.H.M.; Furian, A.F. Chrysin’s anti-inflammatory action in the central nervous system: A scoping review and an evidence-gap mapping of its mechanisms. Eur. J. Pharmacol. 2025, 997, 177602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jeppesen, D.K.; Higginbotham, J.N.; Franklin, J.L.; Coffey, R.J. Comprehensive isolation of extracellular vesicles and nanoparticles. Nat. Protoc. 2023, 18, 1462–1487. [Google Scholar] [CrossRef] [PubMed]
- Deregibus, M.C.; Figliolini, F.; D’ANtico, S.; Manzini, P.M.; Pasquino, C.; De Lena, M.; Tetta, C.; Brizzi, M.F.; Camussi, G. Charge-based precipitation of extracellular vesicles. Int. J. Mol. Med. 2016, 38, 1359–1366. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Marostica, G.; Gelibter, S.; Gironi, M.; Nigro, A.; Furlan, R. Extracellular Vesicles in Neuroinflammation. Front. Cell Dev. Biol. 2021, 8, 623039. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Wang, S.; Chu, C.-H.; Stewart, T.; Ginghina, C.; Wang, Y.; Nie, H.; Guo, M.; Wilson, B.; Hong, J.-S.; Zhang, J. α-Synuclein, a Chemoattractant, Directs Microglial Migration via H2O2-Dependent Lyn Phosphorylation. Proc. Natl. Acad. Sci. USA 2015, 112, E1926–E1935. [Google Scholar] [CrossRef]
- Thawkar, B.S.; Kaur, G. Inhibitors of NF-κB and P2X7/NLRP3/Caspase 1 Pathway in Microglia: Novel Therapeutic Opportu-nities in Neuroinflammation Induced Early-Stage Alzheimer’s Disease. J.Neuroimmunol. 2019, 326, 62–74. [Google Scholar] [CrossRef]
- Rose-John, S.; Jenkins, B.J.; Garbers, C.; Moll, J.M.; Scheller, J. Targeting IL-6 trans-signalling: Past, present and future prospects. Nat. Rev. Immunol. 2023, 23, 666–681. [Google Scholar] [CrossRef]
- Kumari, S.; Dhapola, R.; Sharma, P.; Nagar, P.; Medhi, B.; HariKrishnaReddy, D. The Impact of Cytokines in Neuroinflam-mation-Mediated Stroke. Cytokine Growth Factor Rev. 2024, 78, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.W.; Farooq, M.; Hwang, M.-J.; Haseeb, M.; Choi, S. Autoimmune Neuroinflammatory Diseases: Role of Interleukins. Int. J. Mol. Sci. 2023, 24, 7960. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Wang, F.; Xie, G.; Liu, S.; Li, Z.; Wang, P.; Liu, J.; Lin, L. Gastrodin regulates the TLR4/TRAF6/NF-κB pathway to reduce neuroinflammation and microglial activation in an AD model. Phytomedicine 2024, 128, 155518. [Google Scholar] [CrossRef]
- Khaboushan, A.S.; Yazdanpanah, N.; Rezaei, N. Neuroinflammation and Proinflammatory Cytokines in Epileptogenesis. Mol. Neurobiol. 2022, 59, 1724–1743. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.S.; Prause, M.; Williams, K.; Barrès, R.; Billestrup, N. Butyrate Inhibits IL-1β-Induced Inflammatory Gene Ex-pression by Suppression of NF-κB Activity in Pancreatic Beta Cells. J. Biol. Chem. 2022, 298, 102312. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Bazan, N.G.; Marcheselli, V.L.; Cole–Edwards, K. Brain Response to Injury and Neurodegeneration. Ann. N. Y. Acad. Sci. 2005, 1053, 137–147. [Google Scholar] [CrossRef]
- Okada, S.; Zhang, H.; Hatano, M.; Tokuhisa, T. A Physiologic Role of Bcl-xL Induced in Activated Macrophages. J. Immunol. 1998, 160, 2590–2596. [Google Scholar] [CrossRef]
- Filannino, F.M.; Panaro, M.A.; Benameur, T.; Pizzolorusso, I.; Porro, C. Extracellular Vesicles in the Central Nervous System: A Novel Mechanism of Neuronal Cell Communication. Int. J. Mol. Sci. 2024, 25, 1629. [Google Scholar] [CrossRef]
- Guo, W.; Su, L.; Zhang, H.; Mi, Z. Role of M2 macrophages-derived extracellular vesicles in IL-1β-stimulated chondrocyte proliferation and inflammatory responses. Cell Tissue Bank. 2022, 24, 93–107. [Google Scholar] [CrossRef] [PubMed]
- La Torre, M.E.; Panaro, M.A.; Ruggiero, M.; Polito, R.; Cianciulli, A.; Filannino, F.M.; Lofrumento, D.D.; Antonucci, L.; Benameur, T.; Monda, V.; et al. Extracellular Vesicles Cargo in Modulating Microglia Functional Responses. Biology 2022, 11, 1426. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Che, C.; Yang, S.; Ding, P.; Si, M.; Yang, G. Anti-inflammatory effects of extracellular vesicles from Morchella on LPS-stimulated RAW264.7 cells via the ROS-mediated p38 MAPK signaling pathway. Mol. Cell. Biochem. 2022, 478, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Abd Al Haleem, E.N.; Ahmed, H.I.; El-Naga, R.N. Lycopene and Chrysin through Mitigation of Neuroinflammation and Oxidative Stress Exerted Antidepressant Effects in Clonidine-Induced Depression-like Behavior in Rats. J. Diet. Suppl. 2023, 20, 391–410. [Google Scholar] [CrossRef]
- Xu, M.; Yang, Y.; Peng, J.; Zhang, Y.; Wu, B.; He, B.; Jia, Y.; Yan, T. Effects of Alpinae Oxyphyllae Fructus on microglial polarization in a LPS-induced BV2 cells model of neuroinflammation via TREM2. J. Ethnopharmacol. 2022, 302, 115914. [Google Scholar] [CrossRef]
- Goyal, A.; Singh, G.; Verma, A. A Comprehensive Review on Therapeutic Potential of Chrysin in Brain Related Disorders. CNS Neurol. Disord.–Drug Targets 2023, 22, 789–800. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, J.; Zhu, B.; Shen, J.; Ye, Z.; Peng, Q.; Zhang, Y. Microglia polarization in heat-induced early neural injury. Arch. Med Sci. 2021, 20, 1307–1313. [Google Scholar] [CrossRef]
- Zhang, N.; Cui, Y.; Li, Y.; Mi, Y. A Novel Role of Nogo Proteins: Regulating Macrophages in Inflammatory Disease. Cell. Mol. Neurobiol. 2021, 42, 2439–2448. [Google Scholar] [CrossRef]
- Cheng, L.; Zheng, M.-G.; Jing, J.-H.; Yu, S.-S.; Li, Z.-Y.; Xu, X.-Z.; Yao, F.; Luo, Y.; Liu, Y.-C. M1-type microglia can induce astrocytes to deposit chondroitin sulfate proteoglycan after spinal cord injury. Neural Regen. Res. 2022, 17, 1072–1079. [Google Scholar] [CrossRef]
- Yao, Y.; Li, J.; Zhou, Y.; Wang, S.; Zhang, Z.; Jiang, Q.; Li, K. Macrophage/microglia polarization for the treatment of diabetic retinopathy. Front. Endocrinol. 2023, 14, 1276225. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, C.S.; Seo, K.J.; Kim, I.Y.; Han, S.; Youn, I.; Yune, T.Y. IL-6/JAK2/STAT3 axis mediates neuropathic pain by regulating astrocyte and microglia activation after spinal cord injury. Exp. Neurol. 2023, 370, 114576. [Google Scholar] [CrossRef]
- Madry, C.; Kyrargyri, V.; Arancibia-Cárcamo, I.L.; Jolivet, R.; Kohsaka, S.; Bryan, R.M.; Attwell, D. Microglial Ramification, Surveillance, and Interleukin-1β Release Are Regulated by the Two-Pore Domain K+ Channel THIK-1. Neuron 2018, 97, 299–312.e6. [Google Scholar] [CrossRef]
- Lopez-Rodriguez, A.B.; Hennessy, E.; Murray, C.L.; Nazmi, A.; Delaney, H.J.; Healy, D.; Fagan, S.G.; Rooney, M.; Stewart, E.; Lewis, A.; et al. Acute Systemic Inflammation Exacerbates Neuroinflammation in Alzheimer’s Disease: IL-1β Drives Ampli-fied Responses in Primed Astrocytes and Neuronal Network Dysfunction. Alzheimers Dement 2021, 17, 1735–1755. [Google Scholar] [CrossRef]
- Liang, Z.; Damianou, A.; Vendrell, I.; Jenkins, E.; Lassen, F.H.; Washer, S.J.; Grigoriou, A.; Liu, G.; Yi, G.; Lou, H.; et al. Proximity proteomics reveals UCH-L1 as an essential regulator of NLRP3-mediated IL-1β production in human macrophages and microglia. Cell Rep. 2024, 43, 114152. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Lin, S.; Hu, G.; Wu, J.; Hu, Y.; Nong, F.; Lu, C.; Chen, R.; Liu, J. Molecular Mechanism of Rolupram Reducing Neuroinflammation in Noise Induced Tinnitus Mice through TLR4/NF kB/NLRP3 Protein/Caspase-1/IL-1 β Signaling Path-way. Int. J. Biol. Macromol. 2024, 278, 134987. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Zeng, Y.; Wang, D.; Wen, Y.; Fan, L.; He, Y.; Zhang, J.; Sun, W.; Liu, Y.; et al. Microglia-Neuron Interactions Promote Chronic Itch via the NLRP3-IL-1β-GRPR Axis. Allergy 2023, 78, 1570–1584. [Google Scholar] [CrossRef]
- Du, G.; Yang, Z.; Wen, Y.; Li, X.; Zhong, W.; Li, Z.; Zhang, S.; Luo, E.; Ding, H.; Li, W. Heat stress induces IL-1β and IL-18 overproduction via ROS-activated NLRP3 inflammasome: Implication in neuroinflammation in mice with heat stroke. NeuroReport 2024, 35, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Choudhury, M.E.; Takeda, H.; Sato, A.; Kihara, N.; Mikami, K.; Inoue, A.; Yano, H.; Watanabe, H.; Kumon, Y.; et al. Microglial re-modeling contributes to recovery from ischemic injury of rat brain: A study using a cytokine mixture containing granulocyte-macrophage colony-stimulating factor and interleukin-3. Front. Neurosci. 2022, 16, 941363. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, F.; Ma, H.; Xia, H.; Tian, J.; Sun, T. Amentoflavone Induces Cell Cycle Arrest, Apoptosis, and Autophagy in BV-2 Cells. Front. Biosci. Landmark 2020, 25, 798–816. [Google Scholar]
- Li, Z.; Chu, S.; He, W.; Zhang, Z.; Liu, J.; Cui, L.; Yan, X.; Li, D.; Chen, N. A20 as a novel target for the anti-neuroinflammatory effect of chrysin via inhibition of NF-κB signaling pathway. Brain, Behav. Immun. 2019, 79, 228–235. [Google Scholar] [CrossRef]
- Panaro, M.A.; Benameur, T.; Porro, C. Extracellular Vesicles miRNA Cargo for Microglia Polarization in Traumatic Brain Injury. Biomolecules 2020, 10, 901. [Google Scholar] [CrossRef]
- Bang, O.Y.; Kim, J.-E. Stem cell-derived extracellular vesicle therapy for acute brain insults and neurodegenerative diseases. BMB Rep. 2022, 55, 20–29. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filannino, F.M.; Soleti, R.; Ruggiero, M.; de Stefano, M.I.; Panaro, M.A.; Lofrumento, D.D.; Trotta, T.; Maffione, A.B.; Benameur, T.; Cianciulli, A.; et al. Chrysin-Loaded Extracellular Vesicles Attenuate LPS-Induced Neuroinflammation in BV2 Microglial Cells In Vitro: A Novel Neuroprotective Strategy. Molecules 2025, 30, 3131. https://doi.org/10.3390/molecules30153131
Filannino FM, Soleti R, Ruggiero M, de Stefano MI, Panaro MA, Lofrumento DD, Trotta T, Maffione AB, Benameur T, Cianciulli A, et al. Chrysin-Loaded Extracellular Vesicles Attenuate LPS-Induced Neuroinflammation in BV2 Microglial Cells In Vitro: A Novel Neuroprotective Strategy. Molecules. 2025; 30(15):3131. https://doi.org/10.3390/molecules30153131
Chicago/Turabian StyleFilannino, Francesca Martina, Raffaella Soleti, Melania Ruggiero, Maria Ida de Stefano, Maria Antonietta Panaro, Dario Domenico Lofrumento, Teresa Trotta, Angela Bruna Maffione, Tarek Benameur, Antonia Cianciulli, and et al. 2025. "Chrysin-Loaded Extracellular Vesicles Attenuate LPS-Induced Neuroinflammation in BV2 Microglial Cells In Vitro: A Novel Neuroprotective Strategy" Molecules 30, no. 15: 3131. https://doi.org/10.3390/molecules30153131
APA StyleFilannino, F. M., Soleti, R., Ruggiero, M., de Stefano, M. I., Panaro, M. A., Lofrumento, D. D., Trotta, T., Maffione, A. B., Benameur, T., Cianciulli, A., Calvello, R., Zoila, F., & Porro, C. (2025). Chrysin-Loaded Extracellular Vesicles Attenuate LPS-Induced Neuroinflammation in BV2 Microglial Cells In Vitro: A Novel Neuroprotective Strategy. Molecules, 30(15), 3131. https://doi.org/10.3390/molecules30153131