Comparative Metabolomic Analysis of Three Medicinal Paphiopedilum Species Reveals Divergence in Antioxidant Capacity and Functional Compound Profiles

 ,
,
Abstract
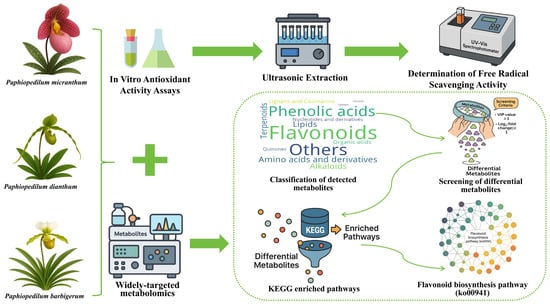
1. Introduction
2. Results
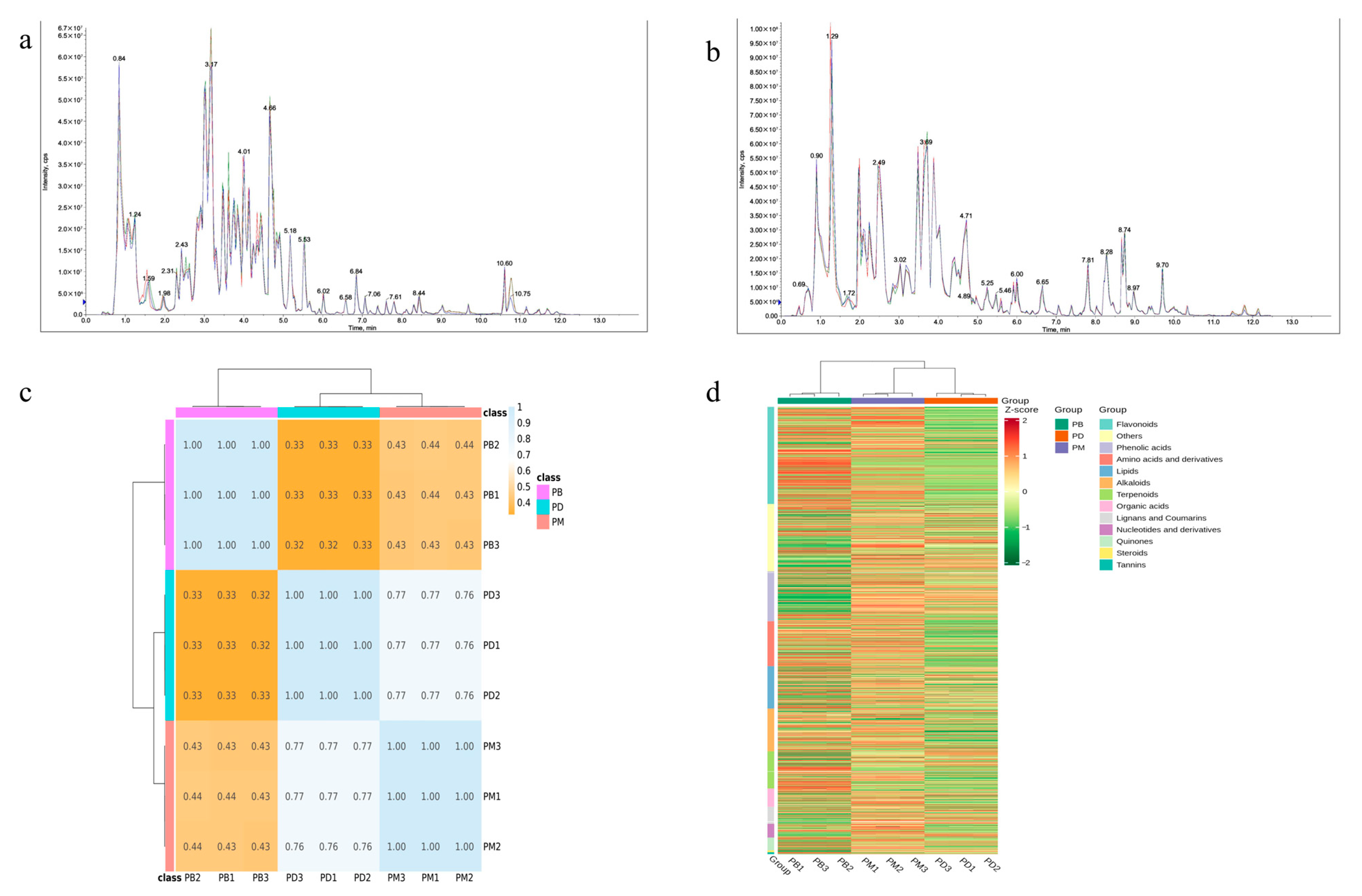
2.1. Metabolic Profiling Analysis
2.2. PCA Analysis
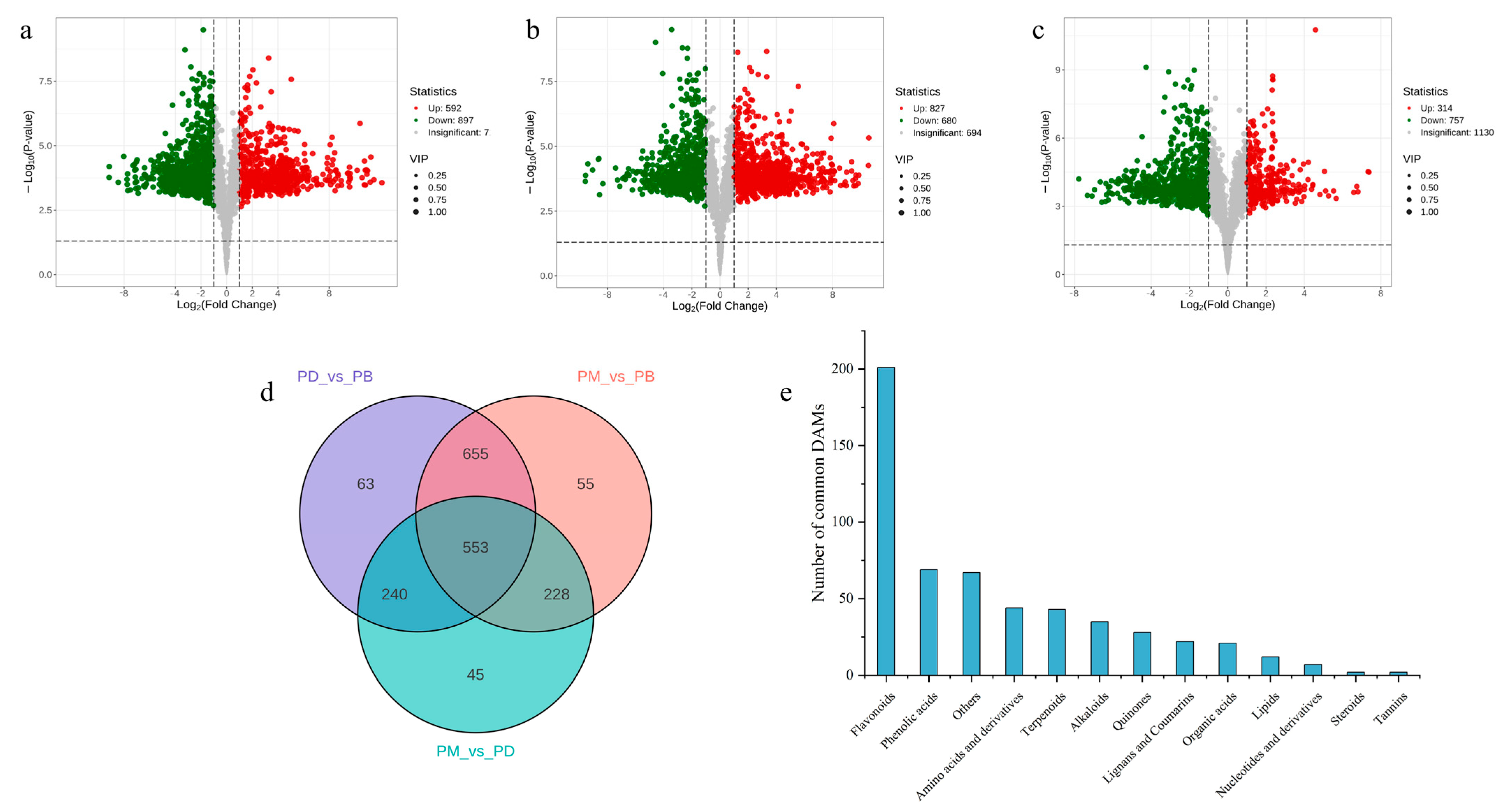
2.3. Screening of Differential Metabolites
2.4. Analysis of Potential Antioxidant Metabolites
2.5. Antioxidant Activity Analysis
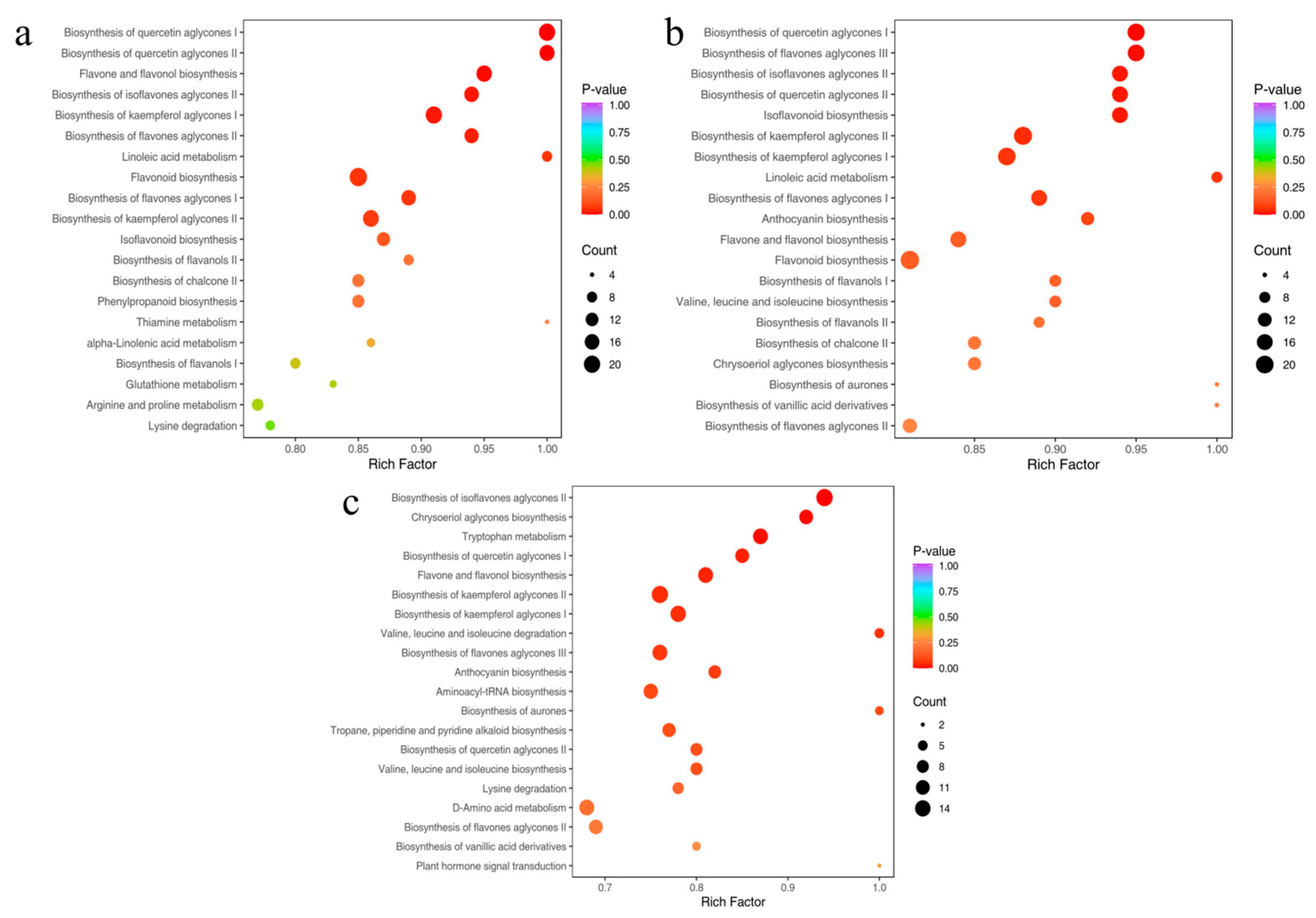
2.6. Enrichment Analysis and KEGG Pathways
2.7. Identification of Core Metabolic Pathways
3. Discussion
3.1. Flavonoids and Phenolic Acids May Serve as Primary Drug-like Components
3.2. Flavonoids May Drive Interspecific Differences in Antioxidant Capacity
3.3. Key Enzyme Expression Differences Drive Flavonoid Metabolic Divergence
4. Materials and Methods
4.1. Materials
4.2. In Vitro Antioxidant Assays
4.2.1. Sample Preparation
4.2.2. DPPH Radical Scavenging Assay
4.2.3. Hydroxyl Radical (·OH) Scavenging Assay
4.2.4. Superoxide Radical (O2−·) Scavenging Assay
4.3. Metabolic Analysis
4.3.1. Sample Pretreatment
4.3.2. UPLC Conditions and ESI-MS/MS System Conditions
4.3.3. Qualitative and Quantitative Determination of Secondary Metabolites
4.3.4. Metabolome Data Processing and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Y.J.; Huang, J.L.; Hu, H.; Zhang, S.B. Research progress on conservation and utilization of germplasm resources of Paphiopedilum species in China. J. West. For. Sci. 2021, 50, 108–112, 119. [Google Scholar]
- Zhang, Y.; An, M.T.; Wu, J.Y.; Yang, Y.B.; Li, Z.; Ye, C. Survival status and protection effectiveness of Paphiopedilum subgenus. Brachypetalum in China. China Environ. Sci. 2022, 42, 3461–3472. [Google Scholar]
- Zhang, J.R.; Fang, L.; Zeng, J.J.; Li, L.; Wu, K.L.; Zeng, S.J. Species and geographical distribution of Paphiopedilum Plants. J. Trop. Crops 2024, 45, 1572–1584. [Google Scholar]
- Yin, Y.Y.; Fang, L.; Li, L.; Chen, Y.; Fu, W.Q.; Wu, K.L.; Zeng, S.J. Research progress on flowering regulation of Paphiopedilum species. J. Trop. Crops 2022, 43, 769–778. [Google Scholar]
- He, J.Z.; Bu, Z.Y.; Li, J.L.; Li, X.L. Introduction and cultivation of Paphiopedilum species. Guangdong Agric. Sci. 2013, 40, 35–37. [Google Scholar]
- Wei, G.W.; Tang, J.M.; Huang, F.K.; Zou, R.; Cen, H.F.; Wei, X.; Wei, L.J.; Ding, T.; Ye, K.Y.; Zeng, D.J.; et al. Medicinal Plant Resources of Hechi; Guangxi Science and Technology Publishing House: Nanning, China, 2025. [Google Scholar]
- Qin, Y. Investigation and Evaluation of Medicinal Plant Resources in Debao County. Master’s Thesis, Guangxi Normal University, Guilin, China, 2019. [Google Scholar]
- Li, T.; Su, C.; Wang, Q.Y. Pharmacognostic identification of Paphiopedilum micranthum. West China J. Pharm. Sci. 2018, 33, 213–215. [Google Scholar]
- Wang, M.; Li, S.; Chen, L.; Li, J.; Rao, W.; Liu, H.; Chen, J.; Ren, H. Conservation and reintroduction of the rare and endangered orchid Paphiopedilum armeniacum. Ecosyst. Health Sustain. Taylor Fr. 2021, 7, 1903817. [Google Scholar] [CrossRef]
- Xiu, M.; Zhang, X.S.; Wei, X.L.; Hong, S.B.; Zheng, Y.H.; Zhang, K.B.; Chen, J.S.; Chen, C.J.; Lin, Y.Y. Adaptability study on introduction and cultivation of Paphiopedilum species in Chaoshan Area. Guangdong Agric. Sci. 2021, 48, 47–56. [Google Scholar]
- Zhang, L.L.; Fang, L.; Li, L.; Wu, K.L.; Zeng, J.Y.; Zhang, J.R.; Kang, M.; Zeng, S.J. Research progress on breeding techniques of Paphiopedilum. Guangxi Sci. 2023, 30, 1052–1059. [Google Scholar]
- Shi, J.Z. Wild Paphiopedilum Plant Resources and Genetic Diversity in the Beipan River Basin. Master’s Thesis, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- Liao, H.Y.; Fan, J.Z.; Li, X.L.; Cui, X.Q.; Wang, X.G.; Ning, Y.F. Germplasm identification and genetic diversity analysis of Paphiopedilum concolor group based on iPBS and ISSR Molecular markers. J. South. Agric. 2023, 54, 2844–2853. [Google Scholar]
- Sun, Y.; Zou, P.; Jiang, N.; Fang, Y.; Liu, G. Comparative analysis of the complete chloroplast genomes of nine Paphiopedilum Species. Front. Genet. 2022, 12, 772415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.P.; Zhang, S.B. Genome size and labellum epidermal cell size are evolutionarily correlated with floral longevity in Paphiopedilum Species. Front. Plant Sci. 2021, 12, 793516. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.T.; Tran, N.; Nguyen, T.D.; Vu, Q.L.; Bui, M.H.; Le, M.T.; Le, L. Complete chloroplast genome of Paphiopedilum delenatii and phylogenetic relationships among Orchidaceae. Plants 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Liu, W.J.; Li, W.; Wu, J.H. Comparative analysis of active constituents and antioxidant activities of Dendrobium officinale in different artificial cultivation models. J. Food Saf. Qual. 2022, 13, 2665–2671. [Google Scholar]
- Chang, M.H.; Jing, Y.Z.; Wang, L. Rapid propagation technique system of Dendrobium officinale. Chin. Tradit. Herb. Drugs 2012, 43, 1412–1417. [Google Scholar]
- Yang, Y. Study on the Quality on Different Planting Patterns of Dendrobium candidum. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2016. [Google Scholar]
- Yang, M.Z.; Shan, Y.Y.; Chen, X.M.; Zhang, C.F.; Li, Z.J. Current development situation of Dendrobium industry in China. Mod. Chin. Med. 2022, 24, 1395–1402. [Google Scholar]
- Barry, H. Free radicals and antioxidants-quo vadis? Trends Pharmacol. Sci. 2010, 32, 125–130. [Google Scholar]
- Helmut, S.; Carsten, B.; Dean, P.J. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar]
- Khan, I.; Ahmad, S. The Impact of Natural Antioxidants on Human Health. In Functional Food Products and Sustainable Health; Springer: Singapore, 2020. [Google Scholar]
- Aguayo-Morales, H.; Poblano, J.; Berlanga, L.; Castillo-Tobías, I.; Silva-Belmares, S.Y.; Cobos-Puc, L.E. Plant antioxidants: Therapeutic potential in cardiovascular diseases. Compounds 2024, 4, 479–502. [Google Scholar] [CrossRef]
- Kozlov, A.V.; Javadov, S.; Sommer, N. Cellular ROS and antioxidants: Physiological and pathological role. Antioxidants 2024, 13, 602. [Google Scholar] [CrossRef]
- Liu, Y.C.; Yang, M.R.; Li, Y.Q.; Liu, Y.Y.; Su, H.Y.; Zhang, W.P.; Liu, Y.; Zhang, Q.L. A multifunctional living hydrogel for the synergistic management of infected diabetic wounds. Mater. Today Bio 2025, 32, 101787. [Google Scholar] [CrossRef]
- Che, H.; Xu, J.; Wu, D.; Chen, S.; Liu, C.; Zhao, C.; Peng, K. Reactive oxygen species-responsive polydopamine-PtCuTe nanoparticle-loaded microneedle system for promoting the healing of infected skin wounds. J. Control. Release 2024, 376, 999–1013. [Google Scholar] [CrossRef]
- Zhuo, D.; Qiang, L.; Meng, D.; Li, X.Y.; He, A.; Xiu, W.J.; Wang, M.J.; Zhang, Y.; Ding, M.; Dong, H.; et al. Radical-scavenging violet phosphorus nanosheets for attenuating hyperinflammation and promoting infected wound healing. Adv. Sci. 2024, 11, e2407545. [Google Scholar]
- Liu, Q.; Wang, C.; Hu, L.; Zhang, Z.; Sun, Q.; Wang, S.; Fan, Y.; Pan, P. Self-healing conductive hydrogels with dynamic dual network structure accelerate infected wound healing via photothermal antimicrobial and regulating inflammatory response. ACS Appl. Mater. Interfaces 2024, 16, 30776–30792. [Google Scholar] [CrossRef]
- Lackner, C.; Tiniakos, D. Fibrosis and alcohol-related liver disease. J. Hepatol. 2019, 70, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumertt, F. Liver fibrosis: Mechanistic concepts and therapeutic perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Beck, M.A.; Levander, O.A. Host nutritional status and its effect on a viral pathogen. J. Infect. Dis. 2000, 182, S93–S96. [Google Scholar] [CrossRef]
- Griffin, D.E. Measles virus-induced suppression of immune responses. Immunol. Rev. 2010, 236, 176–189. [Google Scholar] [CrossRef]
- Shomali, A.; Das, S.; Arif, N.; Sarraf, M.; Zahra, N.; Yadav, V.; Ahmad, P. Diverse physiological roles of flavonoids in plant environmental stress responses and tolerance. Plants 2022, 11, 3158. [Google Scholar] [CrossRef]
- Long, J.; Qin, F.X.; Zhou, J.W.; Ran, J.C.; Qin, L.J. Microenvironment characteristics of the endangered plant Paphiopedilum barbigerum. J. Guizhou Norm. Univ. (Nat. Sci. Ed.) 2009, 27, 15–20. [Google Scholar]
- Kwansu, K.; Sunoh, R.; Haegon, C. Influence of drought stress on chemical composition of sesame seed. Bull. Math. Biol. 2006, 62, 399–428. [Google Scholar]
- Ding, L. Effects of Drought Stress on Antioxidant System and Photosynthetic Characteristics of Cucumber Seedlings. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2013. [Google Scholar]
- Lu, Y.; Ding, Z.; Zhang, D.; Zhu, F.; Gao, B. Integrated metabolomic and transcriptomic analysis reveals the pharmacological effects and differential mechanisms of isoflavone biosynthesis in four species of Glycyrrhiza. Int. J. Mol. Sci. 2025, 26, 2539. [Google Scholar] [CrossRef]
- Sun, F.; Wang, J.; Geng, S.; Liang, Y.; Gong, Z.; Yang, N.; Zheng, J. Comprehensive transcriptomic and metabolomic analysis revealed drought tolerance regulatory pathways in upland cotton. Front. Plant Sci. 2025, 16, 1571944. [Google Scholar] [CrossRef]
- Song, Y.; John Martin, J.J.; Liu, X.; Li, X.; Hou, M.; Zhang, R.; Cao, H. Unraveling the response of secondary metabolites to cold tolerance in oil palm by integration of physiology and metabolomic analyses. BMC Plant Biol. 2025, 25, 279. [Google Scholar] [CrossRef]
- Cui, X.; Qin, X.; Liu, Y.; Zhang, Y.; Bao, H.; Hu, Y.; Shen, X. Analysis of flavonoid metabolism during the process of petal discoloration in three Malus crabapple cultivars. ACS Omega 2022, 7, 37304–37314. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental factors regulate plant secondary metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef]
- Gunasekaran, D.; Tahir, N.I.; Akbar, M.A.; Basir, S.; Ismail, I.; Talip, N.; Bunawan, H. Discovery of anthocyanin biosynthetic pathway in cosmos caudatus kunth. using omics analysis. Agronomy 2021, 11, 661. [Google Scholar]
- Wang, Y.S.; Xu, Y.J.; Gao, L.P.; Yu, O.; Wang, X.Z.; He, X.J.; Jiang, X.L.; Liu, Y.J.; Tao, X. Functional analysis of flavonoid 3′, 5′-hydroxylase from tea plant (Camellia sinensis): Critical role in the accumulation of catechins. BMC Plant Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef]
- Yu, D.; Zhang, K.; Wu, J.; Li, X.; Zhou, G.; Wan, Y. Integrated metabolomic and transcriptomic analysis revealed the flavonoid biosynthesis and regulation in Areca catechu. Phytochem. Anal. 2023, 34, 372–380. [Google Scholar] [CrossRef]
- Parisi, O.; Aiello, D.; Casula, M.; Puoci, F.; Malivindi, R.; Scrivano, L.; Testa, F. Mesoporous nanocrystalline TiO2 loaded with ferulic acid for sunscreen and photo-protection: Safety and efcacy assessment. RSC Adv. 2016, 6, 83767–83775. [Google Scholar] [CrossRef]
- Yang, Y.S.; Chai, S.F.; Tang, J.M.; Luo, Y.J.; Yang, X.X.; Wei, X. Study on extraction process optimization of total polysaccharides from Habenaria dentata and Its Antioxidant Activity. J. Guangxi Acad. Sci. 2022, 38, 188–196. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | PD vs. PB | PM vs. PB | PM vs. PD | |||
|---|---|---|---|---|---|---|
| Up | Down | Up | Down | Up | Down | |
| Flavonoids | 103 | 301 | 192 | 216 | 214 | 80 |
| Phenolic acids | 136 | 62 | 47 | 151 | 80 | 32 |
| Terpenoids | 51 | 87 | 58 | 63 | 42 | 36 |
| Alkaloids | 42 | 72 | 69 | 41 | 77 | 27 |
| Sum | 332 | 522 | 366 | 471 | 413 | 175 |
| Sample | DPPH (IC50, mg/mL) | O2−· (IC50, mg/mL) | OH (IC50, mg/mL) |
|---|---|---|---|
| PB | 0.194 | 0.060 | 0.044 |
| PM | 0.204 | 0.068 | 0.048 |
| PD | 0.490 | 0.082 | 0.058 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sang, J.; Yang, Y.; Xian, K.; Su, J.; Tang, J.; Fu, C.; Tang, F.; Wei, X. Comparative Metabolomic Analysis of Three Medicinal Paphiopedilum Species Reveals Divergence in Antioxidant Capacity and Functional Compound Profiles. Molecules 2025, 30, 2961. https://doi.org/10.3390/molecules30142961
Sang J, Yang Y, Xian K, Su J, Tang J, Fu C, Tang F, Wei X. Comparative Metabolomic Analysis of Three Medicinal Paphiopedilum Species Reveals Divergence in Antioxidant Capacity and Functional Compound Profiles. Molecules. 2025; 30(14):2961. https://doi.org/10.3390/molecules30142961
Chicago/Turabian StyleSang, Jinhan, Yishan Yang, Kanghua Xian, Jiang Su, Jianmin Tang, Chuanming Fu, Fengluan Tang, and Xiao Wei. 2025. "Comparative Metabolomic Analysis of Three Medicinal Paphiopedilum Species Reveals Divergence in Antioxidant Capacity and Functional Compound Profiles" Molecules 30, no. 14: 2961. https://doi.org/10.3390/molecules30142961
APA StyleSang, J., Yang, Y., Xian, K., Su, J., Tang, J., Fu, C., Tang, F., & Wei, X. (2025). Comparative Metabolomic Analysis of Three Medicinal Paphiopedilum Species Reveals Divergence in Antioxidant Capacity and Functional Compound Profiles. Molecules, 30(14), 2961. https://doi.org/10.3390/molecules30142961