



Cytotoxic Activity of Essential Oils from Middle Eastern Medicinal Plants on Malignant Keratinocytes

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Chemical Composition of Essential Oils
2.2. Assessment of Essential Oils’ Effect on Cell Viability
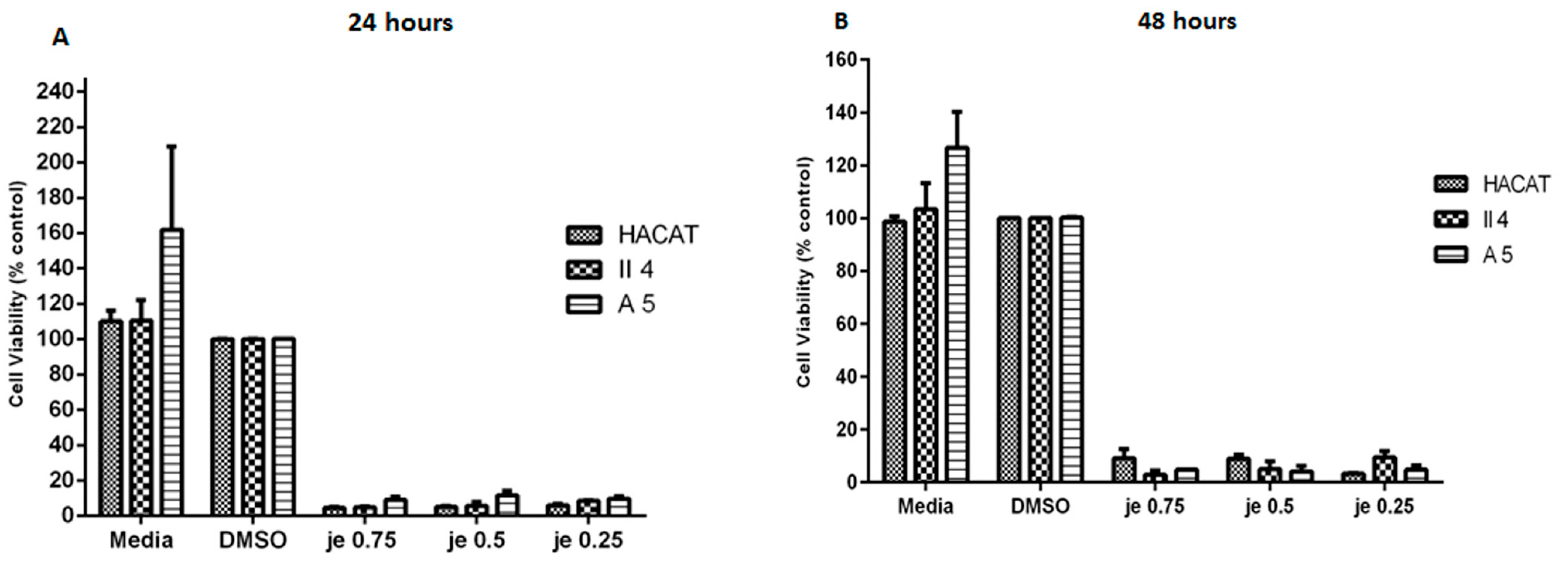
2.2.1. Effect of Juniperus excelsa M. Bieb. (Cupressaceae)
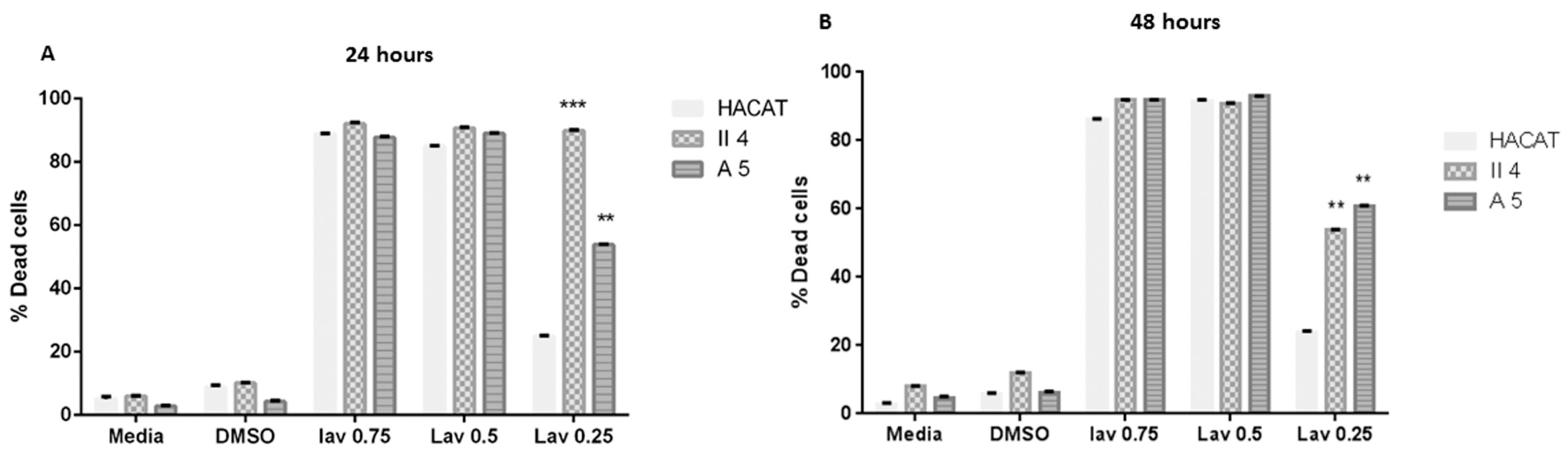
2.2.2. Effect of Lavandula vera DC. (Lamiaceae)
2.2.3. Effect of Salvia fruticosa (Mill.) (Lamiaceae)
2.3. Evaluation of Essential Oils’ Effect on Cell Proliferation
2.3.1. Effect of Juniperus excelsa M. Bieb. (Cupressaceae)
2.3.2. Effect of Lavandula vera DC. (Lamiaceae)
2.3.3. Effect of Salvia fruticosa Mill. (Lamiaceae)
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Collection and Essential Oil Isolation and Analysis
3.3. Essential Oil Dissolution
3.3.1. GC Analyses
3.3.2. GC–MS Analyses
3.3.3. Qualitative and Quantitative Analysis
3.4. Cell Culture
3.5. Cell Viability Analysis: Trypan Blue Exclusion Test
3.6. Cell Proliferation Examination: MTT Assay
3.7. Statistical Data Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gordon, R. Skin Cancer: An Overview of Epidemiology and Risk Factors. Semin. Oncol. Nurs. 2013, 29, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Hyeraci, M.; Papanikolau, E.S.; Grimaldi, M.; Ricci, F.; Pallotta, S.; Monetta, R.; Minafò, Y.A.; Di Lella, G.; Galdo, G.; Abeni, D.; et al. Systemic Photoprotection in Melanoma and Non-Melanoma Skin Cancer. Biomolecules 2023, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Arda, O.; Göksügür, N.; Tüzün, Y. Basic histological structure and functions of facial skin. Clin. Dermatol. 2014, 32, 3–13. [Google Scholar] [CrossRef]
- Abbas, O.; Miller, D.D.; Bhawan, J. Cutaneous Malignant Melanoma. Am. J. Dermatopathol. 2014, 36, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Qadir, M.I.; Ghafoor, S. Malignant Melanoma: Skin Cancer-Diagnosis, Prevention, and Treatment. Crit. Rev. Eukaryot. Gene Expr. 2020, 30, 291–297. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Mehta, A.; Verma, R.S. Essential oils: From prevention to treatment of skin cancer. Drug Discov. Today 2019, 24, 644–655. [Google Scholar] [CrossRef]
- Aziz, Z.A.A.; Ahmad, A.; Setapar, S.H.M.; Karakucuk, A.; Azim, M.M.; Lokhat, D.; Rafatullah, M.; Ganash, M.; Kamal, M.A.; Ashraf, G.M. Essential Oils: Extraction Techniques, Pharmaceutical And Therapeutic Potential—A Review. Curr. Drug Metab. 2018, 19, 1100–1110. [Google Scholar] [CrossRef]
- Emami, S.A.; Abedindo, B.F.; Hassanzadeh-Khayyat, M. Antioxidant Activity of the Essential Oils of Different Parts of Juniperus excelsa M. Bieb. subsp. excelsa and J. excelsa M. Bieb. subsp. polycarpos (K. Koch) Takhtajan (Cupressaceae). Iran. J. Pharm. Res. 2011, 10, 799–810. [Google Scholar]
- Bayala, B.; Bassole, I.H.; Scifo, R.; Gnoula, C.; Morel, L.; Lobaccaro, J.M.; Simpore, J. Anticancer activity of essential oils and their chemical components—A review. Am. J. Cancer Res. 2014, 4, 591–607. [Google Scholar]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of lavender oil and its major components to human skin cells. Cell Prolif. 2004, 37, 221–229. [Google Scholar] [CrossRef]
- Miastkowska, M.; Kantyka, T.; Bielecka, E.; Kałucka, U.; Kamińska, M.; Kucharska, M.; Kilanowicz, A.; Cudzik, D.; Cudzik, K. Enhanced Biological Activity of a Novel Preparation of Lavandula angustifolia Essential Oil. Molecules 2021, 26, 2458. [Google Scholar] [CrossRef] [PubMed]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 16, 55–63. [Google Scholar] [CrossRef]
- Ciążyńska, M.; Olejniczak-Staruch, I.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibińska, M.; Lesiak, A. Ultraviolet Radiation and Chronic Inflammation—Molecules and Mechanisms Involved in Skin Carcinogenesis: A Narrative Review. Life 2021, 11, 326. [Google Scholar] [CrossRef]
- Halliday, G.M. Inflammation, gene mutation and photoimmunosuppression in response to UVR-induced oxidative damage contributes to photocarcinogenesis. Mutat. Res. 2005, 571, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Gieniusz, E.; Skrzydlewska, E.; Łuczaj, W. Current Insights into the Role of UV Radiation-Induced Oxidative Stress in Melanoma Pathogenesis. Int. J. Mol. Sci. 2024, 25, 11651. [Google Scholar] [CrossRef]
- Ahn, C.; Lee, J.; Park, M.; Kim, J.; Yang, J.; Yoo, Y.; Jeung, E. Cytostatic effects of plant essential oils on human skin and lung cells. Exp. Ther. Med. 2020, 19, 2008–2018. [Google Scholar] [CrossRef]
- Ghorbanzadeh, A.; Ghasemnezhad, A.; Sarmast, M.K.; Ebrahimi, S.N. An analysis of variations in morphological characteristics, essential oil content, and genetic sequencing among and within major Iranian Juniper (Juniperus spp.) populations. Phytochemistry 2021, 186, 112737. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Ren, J.-N.; Li, X.; Fan, G.; Qu, S.-S.; Song, Y.; Li, Y.; Pan, S.-Y. Recent updates on bioactive properties of linalool. Food Funct. 2021, 12, 10370–10389. [Google Scholar] [CrossRef]
- Tayarani-Najaran, Z.; Amiri, A.; Karimi, G.; Emami, S.A.; Asili, J.; Mousavi, S.H. Comparative studies of cytotoxic and apoptotic properties of different extracts and the essential oil of Lavandula angustifolia on malignant and normal cells. Nutr. Cancer 2014, 66, 424–434. [Google Scholar] [CrossRef]
- Bethesda (MD): National Institute of Child Health and Human Development. Available online: https://www.ncbi.nlm.nih.gov/books/NBK501865/ (accessed on 25 April 2025).
- Li, K.; Yang, W.; Li, Z.; Jia, W.; Li, J.; Zhang, P.; Xiao, T. Bitter apricot essential oil induces apoptosis of human HaCaT keratinocytes. Int. Immunopharmacol. 2016, 34, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bahri, S.; Jameleddine, S.; Shlyonsky, V. Relevance of carnosic acid to the treatment of several health disorders: Molecular targets and mechanisms. Biomed. Pharmacother. 2016, 84, 569–582. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Lima, M.; de Medeiros, A.A.; Silva, K.S.; Cardoso, G.; de Oliveira Lima, E.; de Oliveira Pereira, F. Investigation of the antifungal potential of linalool against clinical isolates of fluconazole resistant Trichophyton rubrum. J. Mycol. Med. 2017, 27, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Iobbi, V.; Parisi, V.; Bernabè, G.; De Tommasi, N.; Bisio, A.; Brun, P. Anti-Biofilm Activity of Carnosic Acid from Salvia rosmarinus against Methicillin-Resistant Staphylococcus aureus. Plants 2023, 12, 3679. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, H.M.A.; Wilkinson, J.M. Biological activities of Lavender essential oil. Phytother. Res. 2002, 16, 301–308. [Google Scholar] [CrossRef]
- Dhar, P.; Chan, P.; Cohen, D.T.; Khawam, F.; Gibbons, S.; Snyder-Leiby, T.; Dickstein, E.; Rai, P.K.; Watal, G. Synthesis, Anti-microbial Evaluation, and Structure-Activity Relationship of α-Pinene Derivatives. J. Agric. Food Chem. 2014, 62, 3548–3552. [Google Scholar] [CrossRef]
- Singh, J.; Luqman, S.; Meena, A. Carvacrol as a Prospective Regulator of Cancer Targets/Signalling Pathways. Curr. Mol. Pharmacol. 2023, 16, 542–558. [Google Scholar] [CrossRef]
- Birtić, S.; Dussort, P.; Pierre, F.-X.; Bily, A.C.; Roller, M. Carnosic acid. Phytochemistry 2015, 115, 9–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Name | Juniperus excelsa | Salvia fruticosa | Lavandula vera | ||
|---|---|---|---|---|---|
| Yield % (v/w) | 1.17 | 0.9 | 1.25 | ||
| Ri | Compound ID | Identification | |||
| 938 | α-Pinene | Ri, MS, CoGC | 86.8 | 0.8 | 0.1 |
| 953 | Camphene | Ri, MS, CoGC | - | - | 0.1 |
| 980 | β-Pinene | Ri, MS, CoGC | 2.5 | 3.2 | |
| 993 | Myrcene | Ri, MS, CoGC | 3.2 | 1.4 | 0.4 |
| 1013 | δ-3-Carene | Ri, MS | 2.4 | 2.5 | |
| 1030 | Limonene | Ri, MS, CoGC | 2.2 | - | |
| 1034 | 1,8-Cineole | Ri, MS, CoGC | - | 48.7 | 2.8 |
| 1057 | γ-Terpinene | Ri, MS, CoGC | 0.3 | - | |
| 1074 | Linalool oxide | Ri, MS | 0.5 | ||
| 1098 | Linalool | Ri, MS, CoGC | 42.5 | ||
| 1105 | α-Thujone | Ri, MS | - | 0.4 | |
| 1138 | Menthone | Ri, MS | - | - | |
| 1145 | Camphor | Ri, MS, CoGC | trace < 0.05 | 1.1 | 10.4 |
| 1156 | Isoborneol | 1.1 | |||
| 1166 | Borneol | 5.6 | |||
| 1168 | Lavandulol | Ri, MS | - | - | 0.2 |
| 1176 | Terpinen-4-ol | 10.5 | |||
| 1189 | alpha-terpineol | Ri, MS | 0.4 | - | 0.9 |
| 1191 | Hexyl butyrate | 2.5 | |||
| 1217 | Verbenone | Ri, MS | 0.1 | - | |
| 1233 | Pulegone | Ri, MS, CoGC | - | - | |
| 1264 | Linalyl acetate | 8.6 | |||
| 1289 | Lavandulyl acetate | 1.9 | |||
| 1329 | Piperitone | Ri, MS, CoGC | - | - | |
| 1343 | Piperitenone | Ri, MS | - | - | |
| 1365 | Neryl acetate | 0.3 | |||
| 1369 | Piperitenone oxide | Ri, MS | |||
| 1383 | Geranyl acetate | 1.1 | |||
| 1404 | (Z)-Caryophyllene | 1.1 | |||
| 1415 | β-Caryophyllene | Ri, MS, CoGC | - | 30.8 | |
| 1437 | Aromadendrene | Ri, MS | - | 3.3 | |
| 1452 | β-Farnesene | 3.1 | |||
| 1455 | α-Humulene | Ri, MS | - | 2.8 | |
| 1463 | allo-Aromadendrene | Ri, MS | - | 0.3 | |
| 1491 | Bicyclogermacrene | Ri, MS | - | - | |
| 1500 | Lavandulyl isovalerate | 1.6 | |||
| 1515 | δ-Cadinene | Ri, MS | trace < 0.05 | 0.1 | |
| 1566 | α-Nerolidol | 0.2 | |||
| 1577 | Caryophyllene oxide | 0.5 | |||
| Total | 97.8 | 95.5 | 87.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othman, R.; Moarbes, V.; Zaatar, M.T.; Antonios, D.; Roufayel, R.; Beyrouthy, M.; Fajloun, Z.; Sabatier, J.-M.; Karam, M. Cytotoxic Activity of Essential Oils from Middle Eastern Medicinal Plants on Malignant Keratinocytes. Molecules 2025, 30, 2844. https://doi.org/10.3390/molecules30132844
Othman R, Moarbes V, Zaatar MT, Antonios D, Roufayel R, Beyrouthy M, Fajloun Z, Sabatier J-M, Karam M. Cytotoxic Activity of Essential Oils from Middle Eastern Medicinal Plants on Malignant Keratinocytes. Molecules. 2025; 30(13):2844. https://doi.org/10.3390/molecules30132844
Chicago/Turabian StyleOthman, Rima, Vanessa Moarbes, Muriel Tahtouh Zaatar, Diane Antonios, Rabih Roufayel, Marc Beyrouthy, Ziad Fajloun, Jean-Marc Sabatier, and Marc Karam. 2025. "Cytotoxic Activity of Essential Oils from Middle Eastern Medicinal Plants on Malignant Keratinocytes" Molecules 30, no. 13: 2844. https://doi.org/10.3390/molecules30132844
APA StyleOthman, R., Moarbes, V., Zaatar, M. T., Antonios, D., Roufayel, R., Beyrouthy, M., Fajloun, Z., Sabatier, J.-M., & Karam, M. (2025). Cytotoxic Activity of Essential Oils from Middle Eastern Medicinal Plants on Malignant Keratinocytes. Molecules, 30(13), 2844. https://doi.org/10.3390/molecules30132844