How Do Molecular Tweezers Bind to Proteins? Lessons from X-ray Crystallography

 , and
, and
Abstract
1. Introduction
2. Results
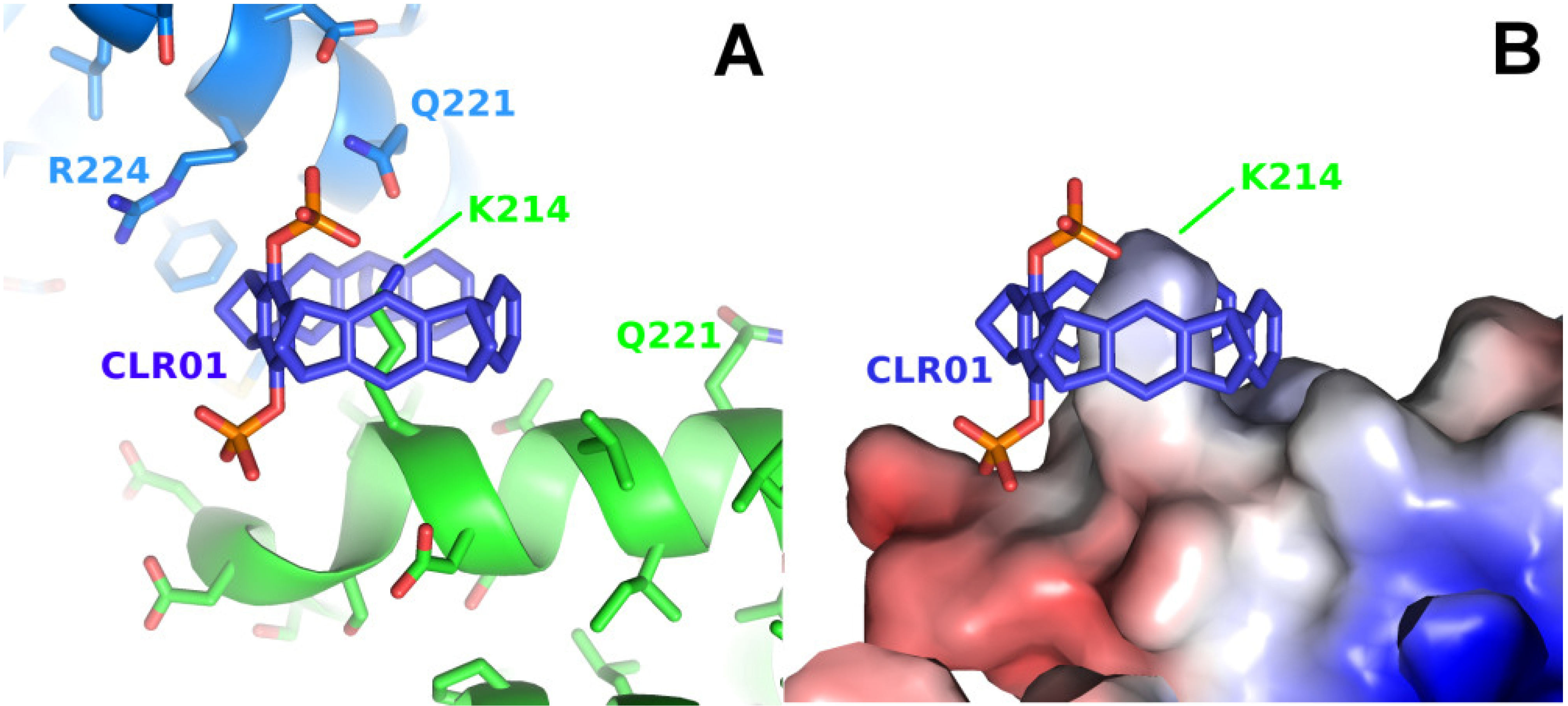
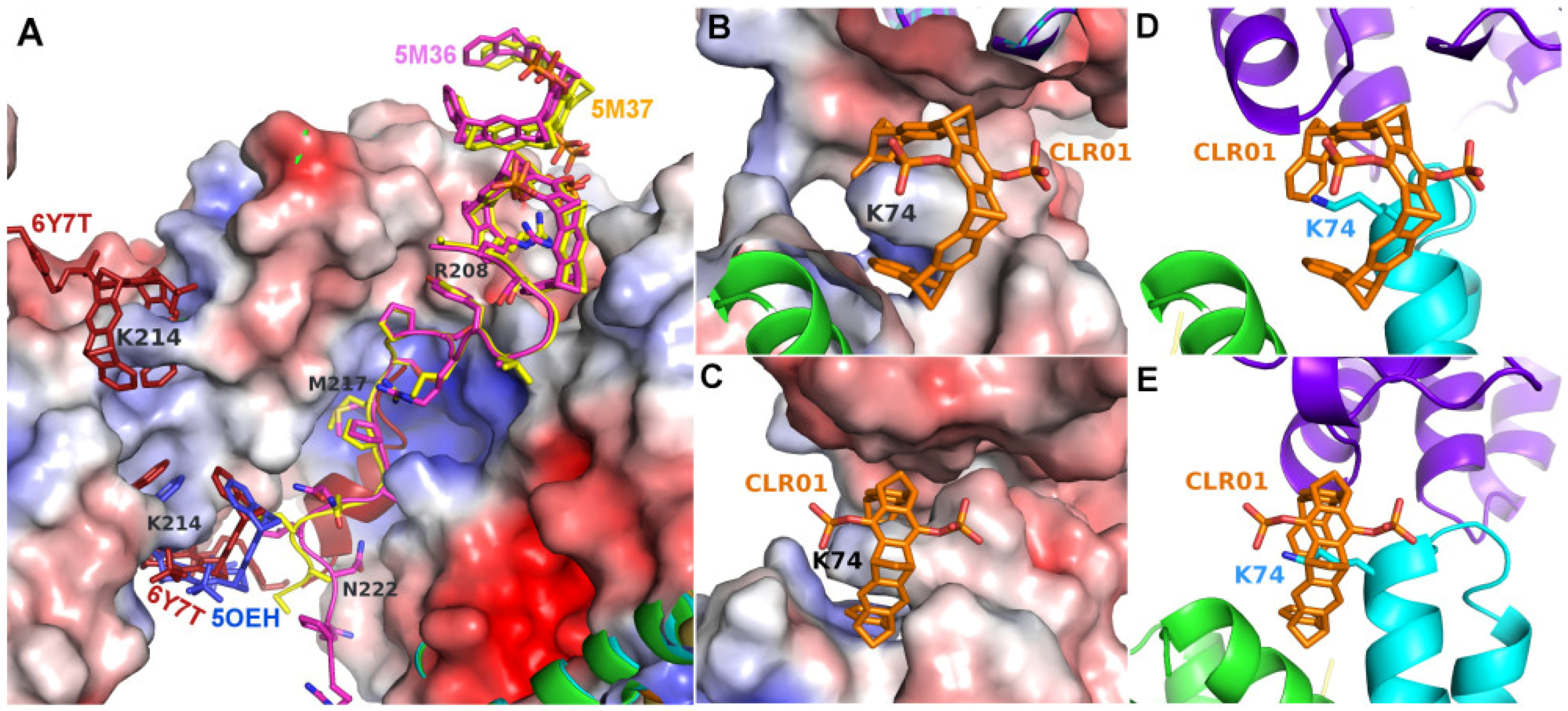
2.1. Analysis of Published Structures: 14-3-3–Tweezer Complexes
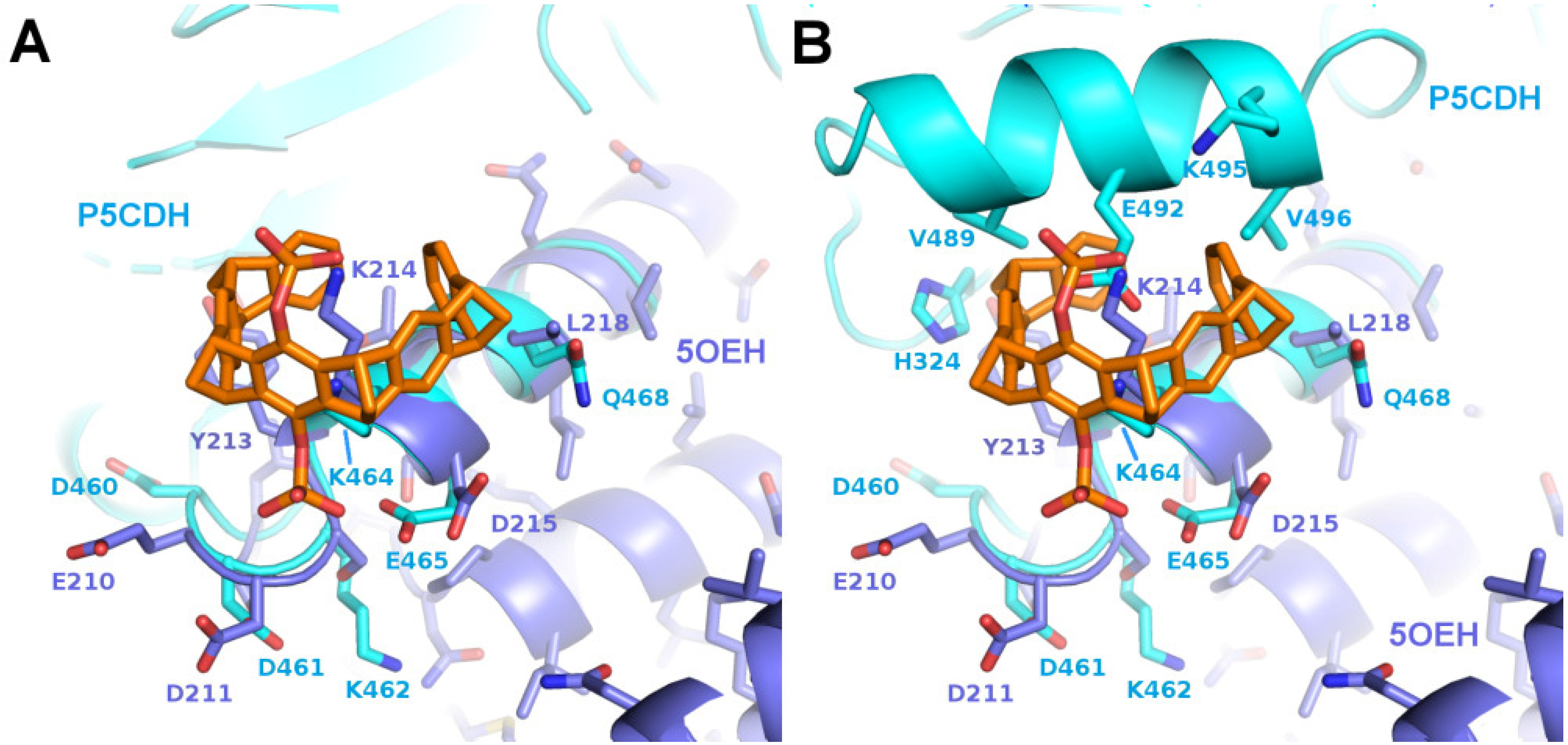
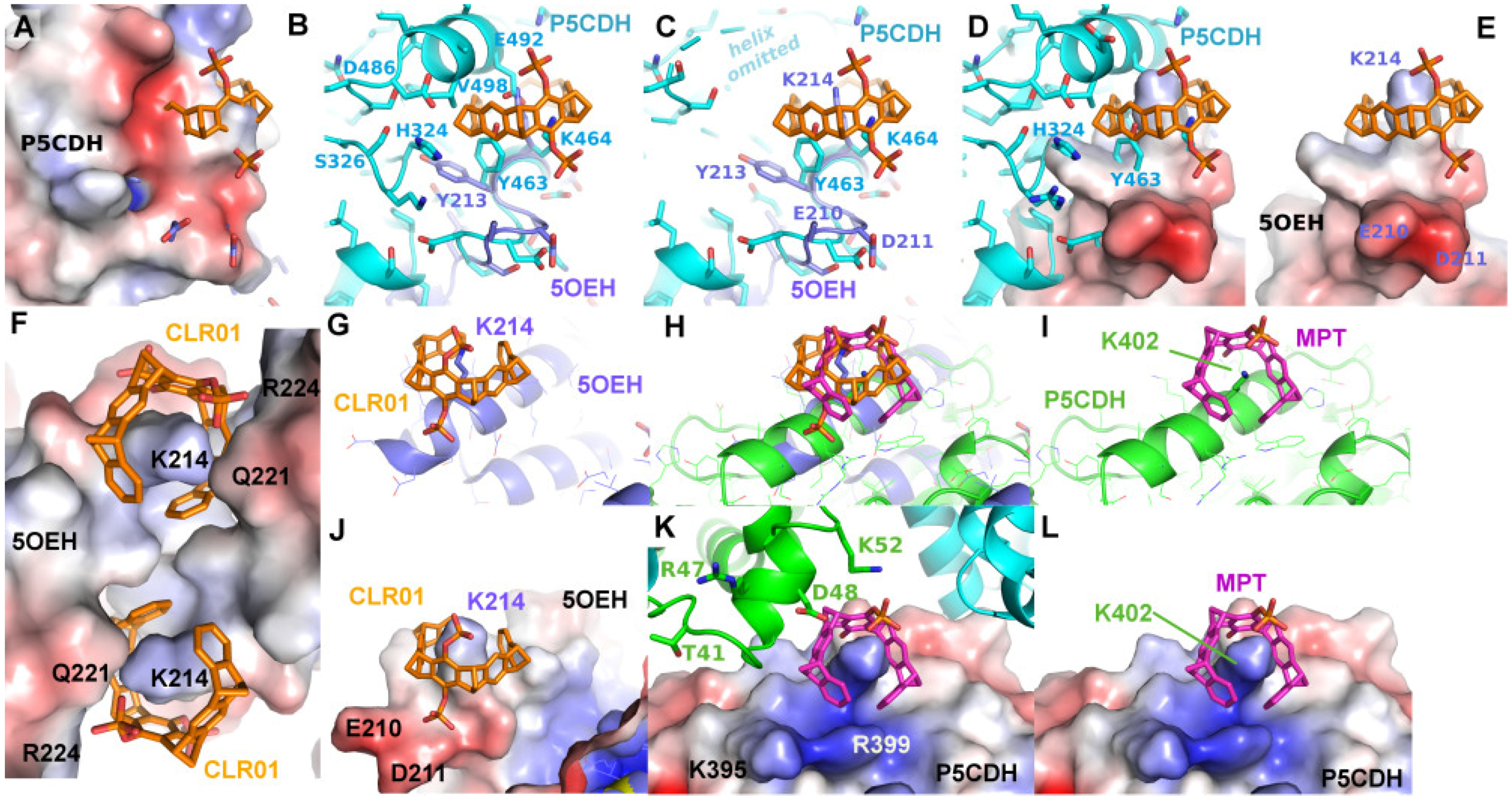
2.2. Binding of CLR01 to Sterically Related Sites of 14-3-3 Proteins—P5CDH as Model System
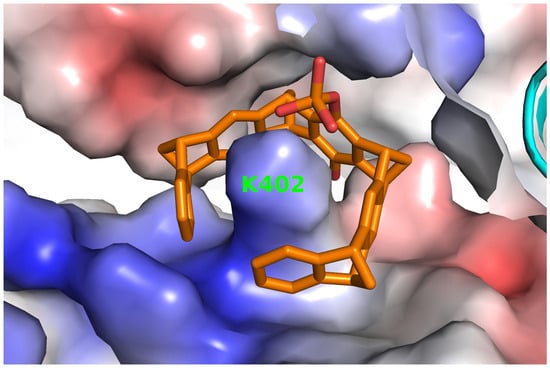
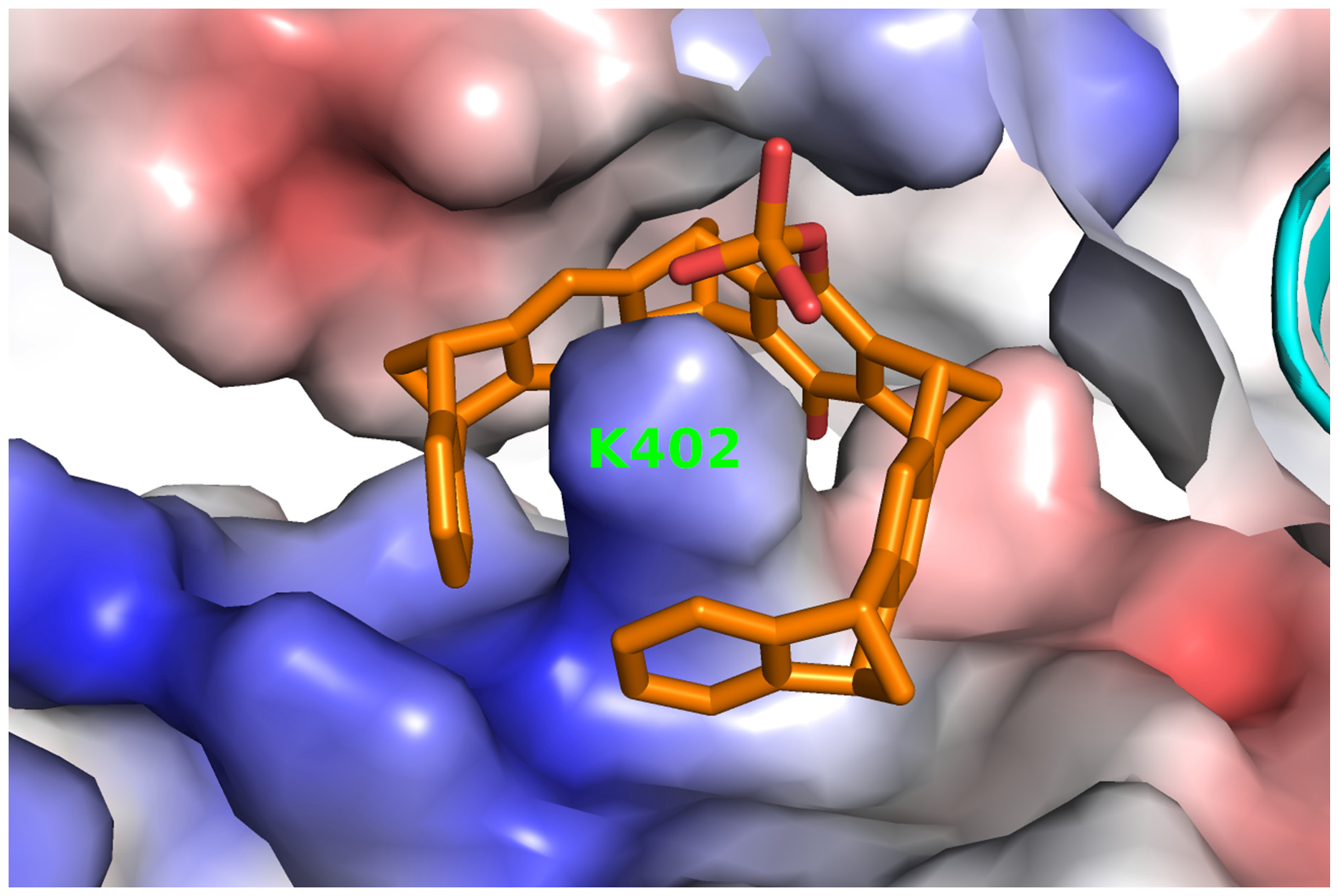
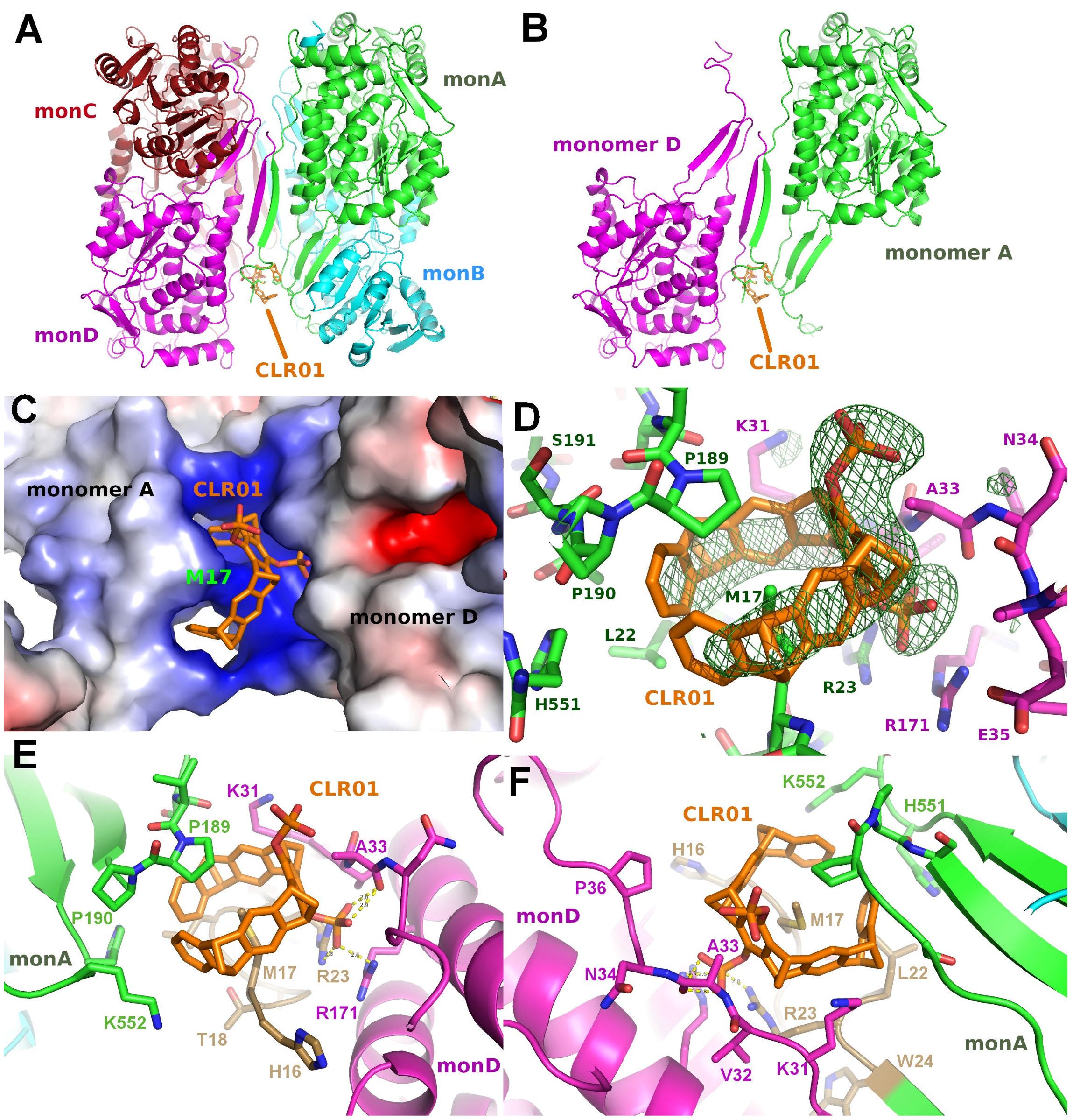
2.3. Structure of P5CDH with the Diphosphate Tweezer CLR01
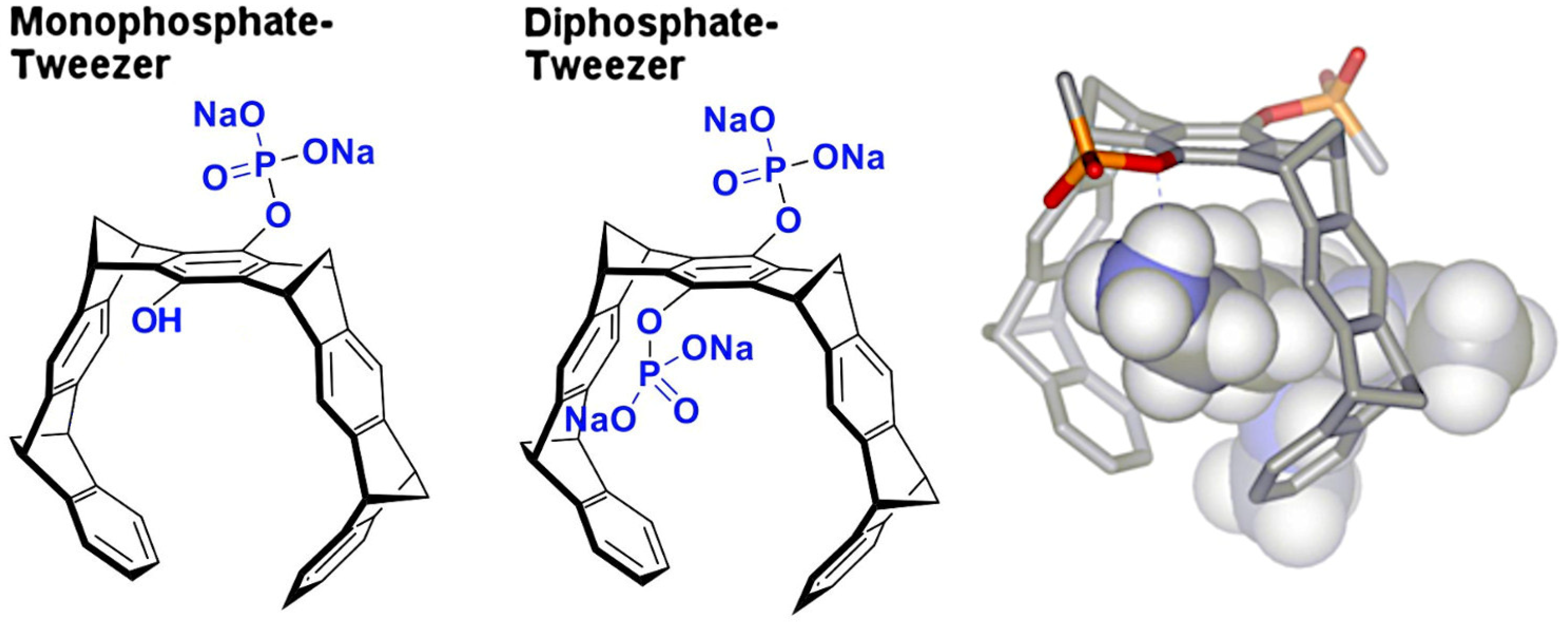
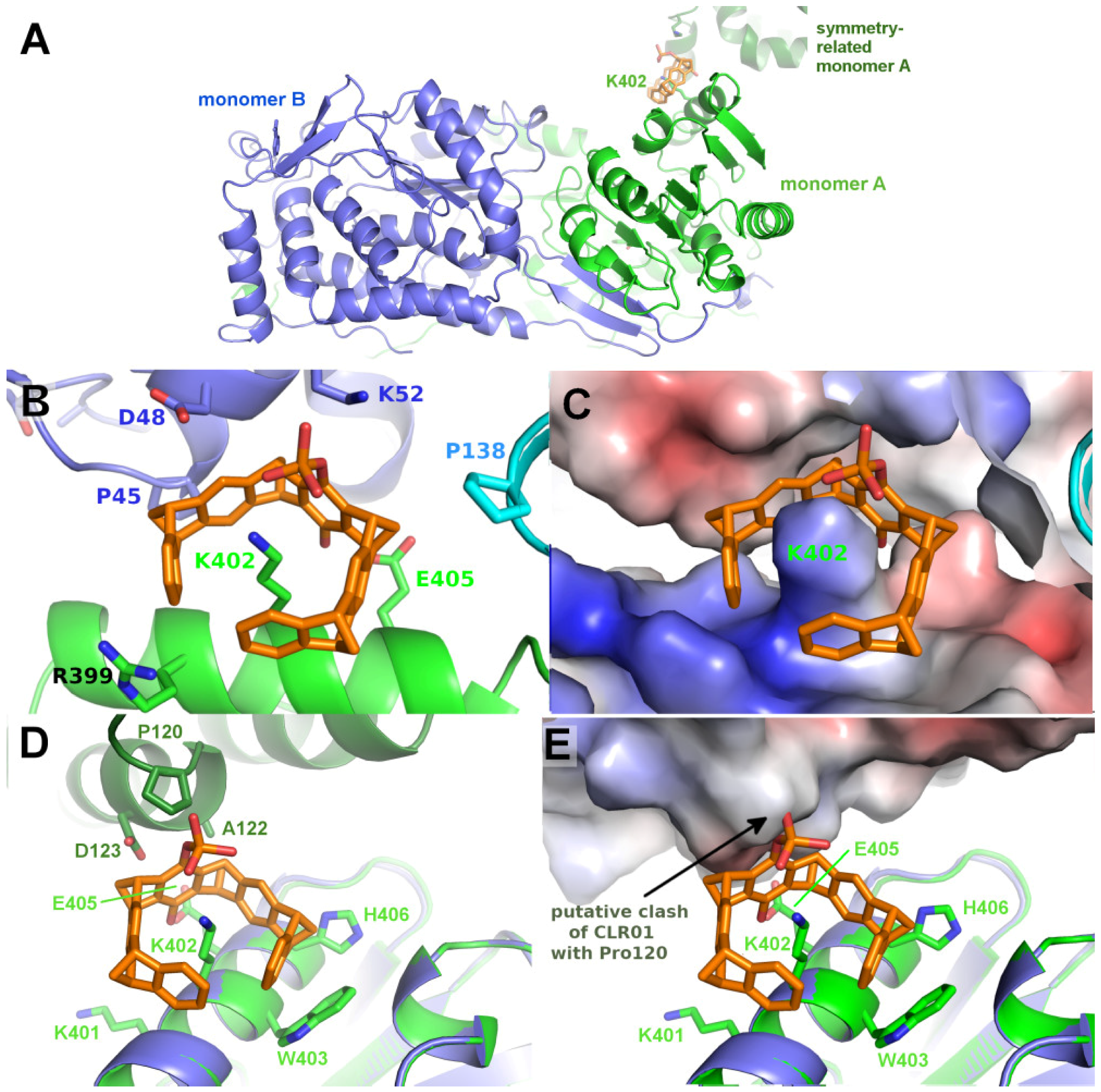
2.4. Structure of P5CDH with the Monophosphate Tweezer
3. Discussion
4. Conclusions
5. Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References and Notes
- Huttlin, E.L.; Bruckner, R.J.; Paulo, J.A.; Cannon, J.R.; Ting, L.; Baltier, K.; Colby, G.; Gebreab, F.; Gygi, M.P.; Parzen, H.; et al. Architecture of the human interactome defines protein communities and disease networks. Nature 2017, 545, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Milroy, L.-G.; Grossmann, T.N.; Hennig, S.; Brunsveld, L.; Ottmann, C. Modulators of protein-protein interactions. Chem. Rev. 2014, 114, 4695–4748. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.E.; Bayly, A.R.; Abell, C.; Skidmore, J. Small molecules, big targets: Drug discovery faces the protein-protein interaction challenge. Nat. Rev. Drug Discov. 2016, 15, 533–550. [Google Scholar] [CrossRef] [PubMed]
- van Dun, S.; Ottmann, C.; Milroy, L.-G.; Brunsveld, L. Supramolecular Chemistry Targeting Proteins. J. Am. Chem. Soc. 2017, 139, 13960–13968. [Google Scholar] [CrossRef]
- Yang, X.; Lennard, K.R.; He, C.; Walker, M.C.; Ball, A.T.; Doigneaux, C.; Tavassoli, A.; van der Donk, W.A. A lanthipeptide library used to identify a protein–protein interaction inhibitor. Nat. Chem. Biol. 2018, 4, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Nobel prizes in Supramolecular Chemistry—1987: Charles J. Pedersen, Donald J. Cram, Jean-Marie Lehn; 2016: Bernard L. Feringa, Sir J. Fraser Stoddart, Jean-Pierre Sauvage.
- Hof, F. Host-guest chemistry that directly targets lysine methylation: Synthetic host molecules as alternatives to bio-reagents. Chem. Commun. 2016, 52, 10093–10108. [Google Scholar] [CrossRef] [PubMed]
- Hatai, J.; Schmuck, C. Diverse Properties of Guanidiniocarbonyl Pyrrole-Based Molecules: Artificial Analogues of Arginine. Acc. Chem. Res. 2019, 52, 1709–1720. [Google Scholar] [CrossRef] [PubMed]
- Sonzini, S.; Marcozzi, A.; Gubeli, R.J.; van der Walle, C.F.; Ravn, P.; Herrmann, A.; Scherman, O.A. High Affinity Recognition of a Selected Amino Acid Epitope within a Protein by Cucurbit [8] uril Complexation. Angew. Chem. Int. Ed. 2016, 55, 14000–14004. [Google Scholar] [CrossRef] [PubMed]
- Vilenchik, L.Z.; Griffith, J.P.; St. Clair, N.; Navia, M.A.; Margolin, A.L. Protein Crystals as Novel Microporous Materials. J. Am. Chem. Soc. 1998, 120, 4290–4294. [Google Scholar] [CrossRef]
- Vink, P.J.; de Briels, J.M.; Schrader, T.; Milroy, L.-G.; Brunsveld, L.; Ottmann, C. A Binary Bivalent Supramolecular Assembly Platform Based on Cucurbit8uril and Dimeric Adapter Protein 14-3-3. Angew. Chem. Int. Ed. 2017, 56, 8998–9002. [Google Scholar] [CrossRef]
- Ehlers, M.; Grad, J.-N.; Mittal, S.; Bier, D.; Mertel, M.; Ohl, L.; Bartel, M.; Briels, J.; Heimann, M.; Ottmann, C.; et al. Rational Design, Binding Studies, and Crystal-Structure Evaluation of the First Ligand Targeting the Dimerization Interface of the 14-3-3ζ Adapter Protein. Chembiochem 2018, 19, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Sijbesma, E.; Visser, E.; Plitzko, K.; Thiel, P.; Milroy, L.-G.; Kaiser, M.; Brunsveld, L.; Ottmann, C. Structure-based evolution of a promiscuous inhibitor to a selective stabilizer of protein-protein interactions. Nat. Commun. 2020, 11, 3954. [Google Scholar] [CrossRef] [PubMed]
- Crowley, P.B. Protein–Calixarene Complexation: From Recognition to Assembly. Acc. Chem. Res. 2022, 55, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, M.; Schrader, T.; Klärner, F.-G. A Molecular Tweezer for Lysine and Arginine. J. Am. Chem. Soc. 2005, 127, 14415–14421. [Google Scholar] [CrossRef] [PubMed]
- Dutt, C.S.; Wilch, C.; Gersthagen, T.; Wölper, C.; Sowislok, A.A.; Klärner, F.-G.; Schrader, T. Linker effects on Amino Acid and Peptide Recognition by Molecular Tweezers. Eur. J. Org. Chem. 2013, 7705–7714. [Google Scholar] [CrossRef]
- Meiners, A.; Bäcker, S.; Hadrović, I.; Heid, C.; Beuck, C.; Ruiz-Blanco, Y.B.; Mieres-Perez, J.; Pörschke, M.; Grad, J.-N.; Vallet, C.; et al. Specific inhibition of the Survivin–CRM1 interaction by peptide-modified molecular tweezers. Nat. Commun. 2021, 12, 1505. [Google Scholar] [CrossRef] [PubMed]
- Schrader, T.; Bitan, G.; Klärner, F.-G. Molecular tweezers for lysine and arginine—Powerful inhibitors of pathologic protein aggregation. Chem. Commun. 2016, 52, 11318–11334. [Google Scholar] [CrossRef]
- Lump, E.; Castellano, L.M.; Meier, C.; Seeliger, J.; Erwin, N.; Sperlich, B.; Stürzel, C.M.; Usmani, S.; Hammond, R.M.; von Einem, J.; et al. A molecular tweezer antagonizes seminal amyloid and HIV infection. eLife 2015, 4, e05397. [Google Scholar] [CrossRef] [PubMed]
- McGovern, R.; Fernandes, H.; Khan, A.; Power, N.P.; Crowley, P.B. Protein camouflage in cytochrome c–calixarene complexes. Nat. Chem. 2012, 4, 527–533. [Google Scholar] [CrossRef]
- Mallon, M.; Dutt, S.; Schrader, T.; Crowley, P.B. Protein Camouflage: Supramolecular Anion Recognition by Ubiquitin. ChemBioChem 2016, 17, 774–783. [Google Scholar] [CrossRef]
- Trusch, F.; Kowski, K.; Bravo-Rodriguez, K.; Beuck, C.; Sowislok, A.; Wettig, B.; Matena, A.; Sanchez-Garcia, E.; Meyer, H.; Schrader, T.; et al. Molecular tweezers target a protein–protein interface and thereby modulate complex formation. Chem. Commun. 2016, 52, 14141–14144. [Google Scholar] [CrossRef] [PubMed]
- Bier, D.; Rose, R.; Bravo-Rodriguez, K.; Bartel, M.; Ramirez-Anguita, J.M.; Dutt, S.; Wilch, C.; Klärner, F.-G.; Sanchez-Garcia, E.; Schrader, T. Molecular tweezers modulate 14-3-3 protein-protein interactions. Nat. Chem. 2013, 5, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Bier, D.; Mittal, S.; Bravo-Rodriguez, K.; Sowislok, A.; Briels, J.; Heid, C.; Bartel, M.; Wettig, B.; Brunsveld, L.; Sanchez-Garcia, E.; et al. The molecular tweezer CLR01 stabilizes a disordered protein-protein interface. J. Am. Chem. Soc. 2017, 139, 16256–16263. [Google Scholar] [CrossRef] [PubMed]
- Dutt, S.; Wilch, C.; Gersthagen, T.; Talbiersky, P.; Rodriguez, K.; Hanni, M.; Sánchez-García, E.; Ochsenfeld, C.; Klärner, F.-G.; Schrader, T. Molecular Tweezers with Varying Anions: A Comparative Study. J. Org. Chem. 2013, 78, 6721–6734. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bosica, F.; Andrei, S.A.; Neves, J.F.; Brandt, P.; Gunnarsson, A.; Landrieu, I.; Ottmann, C.; O’Mahony, G. Design of Drug-Like Protein-Protein Interaction Stabilizers Guided By Chelation-Controlled Bioactive Conformation Stabilization. Chem. Eur. J. 2020, 26, 7131–7139. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, N.; Fukata, Y.; Okatsu, K.; Yamagata, A.; Liu, Y.; Sanbo, M.; Miyazaki, Y.; Goto, T.; Abe, M.; Kassai, H.; et al. 14-3-3 proteins stabilize LGI1-ADAM22 levels to regulate seizure thresholds in mice. Cell Rep. 2021, 37, 110107. [Google Scholar] [CrossRef]
- Yilmaz, E.; Bier, D.; Guillory, X.; Briels, J.; Ruiz-Blanco, Y.B.; Sanchez-Garcia, E.; Ottmann, C.; Kaiser, M. Mono- and Bivalent 14-3-3 Inhibitors for Characterizing Supramolecular “Lysine Wrapping” of Oligoethylene Glycol (OEG) Moieties in Proteins. Chem. Eur. J. 2018, 24, 13807. [Google Scholar] [CrossRef]
- Molzan, M.; Ottmann, C. Synergistic binding of the phosphorylated S233- and S259-binding sites of C-RAF to one 14-3-3zeta dimer. J. Mol. Biol. 2012, 423, 486–495. [Google Scholar] [CrossRef]
- Guillory, X.; Hadrovic, I.; Vink, P.J.; de Sowislok, A.; Brunsveld, L.; Schrader, T.; Ottmann, C. Supramolecular Enhancement of a Natural 14-3-3 Protein Ligand. J. Am. Chem. Soc. 2021, 143, 13495–13500. [Google Scholar] [CrossRef]
- Srivastava, D.; Singh, R.K.; Moxley, M.A.; Henzl, M.T.; Becker, D.F.; Tanner, J.J. The three-dimensional structural basis of type II hyperprolinemia. J. Mol. Biol. 2012, 420, 176–189. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Søndergaard, C.R.; Garrett, A.E.; Carstensen, T.; Pollastri, G.; Nielsen, J.E. Structural artifacts in protein-ligand X-ray structures: Implications for the development of docking scoring functions. J. Med. Chem 2009, 52, 5673–5684. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.E.; Cowan, J.A. Mg2+ chemistry and biochemistry. Biometals 2002, 15, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Romani, A.; Scarpa, A. Regulation of cell magnesium. Arch. Biochem. Biophys. 1992, 298, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, A.; Brinley, F.J. In situ measurements of free cytosolic magnesium ions. Fed. Proc. 1981, 40, 2646–2652. [Google Scholar] [PubMed]
- Pepperell, J.R.; Kommineni, K.; Buradagunta, S.; Smith, P.J.S.; Keefe, D.L. Transmembrane Regulation of Intracellular Calcium by a Plasma Membrane Sodium/Calcium Exchanger in Mouse Ova. Biol. Reprod. 1999, 60, 1137–1143. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kabsch, W. Automatic processing of rotation diffraction data from crystals of initially unknown symmetry and cell constants. J. Appl. Crystallogr. 1993, 26, 795–800. [Google Scholar] [CrossRef]
- Collaborative Computational Project, Number 4. “The CCP4 Suite: Programs for Protein Crystallography”. Acta Crystallogr. 1994, D50, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. 2010, D66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. 2004, D60, 2126–2132. [Google Scholar] [CrossRef]
- Likova, E.; Petkov, P.; Ilieva, N.; Litov, L. The PyMOL Molecular Graphics System, Version 2.0; Schrödinger, LLC: New York, NY, USA, 2015. [Google Scholar]
- Pemberton, T.A.; Tanner, J.J. Structural basis of substrate selectivity of Δ1-pyrroline-5-carboxylate dehydrogenase (ALDH4A1): Semialdehyde chain length. Arch. Biochem. Biophys. 2013, 538, 34–40. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Resol. | M | SG | Unit Cell | Guest | Crystallization Condition |
|---|---|---|---|---|---|---|
| P5CDH apo (3V9J/ 3V9K/3V9L) | 1.3–1.5 Å | 2 | P212121 | 85, 94, 132 Å 90, 90, 90° | - | 20–25% w/v PEG3350, 0.2 M LiSO4, 0.1 M Bis-Tris pH 6.5 |
| P5CDH apo (3V9G) | 2.5 Å | 4 | P65 | 150.7, 150.7, 192.0 Å, 90, 90, 120° | - | 22.5% PEG3350, 0.2 M (NH4)3 sulfate, 0.1 M HEPES pH 7.5 |
| P5CDH-CLR01 | 2.6 Å | 4 | P65 | 148.8, 148.78, 190.0 Å 90, 90, 120° | Met | 0.2 M (NH4)3 citrate, 20% PEG 3350 |
| P5CDH-MPT | 1.2 Å | 2 | P21 | 71.9, 85.1, 85.6 Å 90, 103.2, 90° | Lys | 25%PEG3350, 0.1 M TRIS pH 8.5 |
| P5CHD apo (4OE5) | 1.95 Å | 4 | P21 | 92, 121.3, 93.4 Å 90, 104.2, 90° | - | 20–25% w/v PEG3350, 0.2 M MgCl2, 0.1 M HEPES pH 7–8 |
| 14-3-3-CLR01 (5OEH) | 2.35 Å | 1 | C222 | 60, 157, 77 Å 90, 90, 90° | Lys | 1.26 M tri-sodium citrate, 10%glycerol, 0.09 M Na-HEPES pH 7.5 |
| 14-3-3-CLR01+ non-coval. peptide (5M36/37) | 2.35 Å | 2 | P212121 | 5M36: 71.5, 102.4, 112.8 Å, 90, 90, 90° 5M37: 71.2, 88.2, 112.6 Å 90, 90, 90° | Arg, Lys | 25.5% PEG 4000, 0.17 M (NH4)3 acetate, 0.085 M tri-sodium citrate pH 5.6, 15% glycerol (same condition for 5M35, 5M36 and 5M37) |
| 14-3-3-CLR01-peptide-hybrid (6Y7T) | 2.5 Å | 4 | I2 | 145.2, 63.1, 167.0 Å 90, 101.3,90° | Lys | 20% PEG1000, 0.2 M Na acetate trihydrate, bis-tris-propane pH 7, 10% glycerol |
| P5CDH-CLR01 (PDB 8RKQ) | P5CDH-MPT (PDB 8RKR) | |
|---|---|---|
| Wavelength | 0.97794 | 0.9188 |
| Resolution range | 48.21–2.6 (2.693–2.6) | 42.13–1.2 (1.243–1.2) |
| Space group | P65 | P21 |
| Unit cell | 148.8 148.8 190.0 90 90 120 | 71.9 85.2 85.6 90 103.1 90 |
| Total reflections | 654389 (62098) | 2106606 (208643) |
| Unique reflections | 72711 (7197) | 312301 (31102) |
| Multiplicity | 9.0 (8.6) | 6.7 (6.7) |
| Completeness (%) | 99.56 (99.08) | 99.88 (99.85) |
| Mean I/sigma(I) | 9.48 (0.77) | 11.66 (0.76) |
| Wilson B-factor | 73.60 | 14.31 |
| R-merge | 0.1317 (1.977) | 0.08005 (2.148) |
| R-meas | 0.1399 (2.103) | 0.08677 (2.33) |
| R-pim | 0.04687 (0.7122) | 0.03313 (0.8898) |
| CC1/2 | 0.998 (0.656) | 0.999 (0.344) |
| CC * | 0.999 (0.89) | 1 (0.716) |
| Reflections in refinement | 72706 (7197) | 312278 (31102) |
| Reflections for R-free | 2097 (207) | 3279 (326) |
| R-work | 0.2311 (0.4023) | 0.1725 (0.3326) |
| R-free | 0.2713 (0.4090) | 0.1914 (0.3463) |
| CC(work) | 0.945 (0.640) | 0.973 (0.645) |
| CC(free) | 0.930 (0.514) | 0.974 (0.612) |
| Non-hydrogen atoms | 16936 | 9626 |
| macromolecules | 16829 | 8410 |
| ligands | 52 | 84 |
| solvent | 55 | 1132 |
| Protein residues | 2176 | 1064 |
| RMS(bonds) | 0.014 | 0.014 |
| RMS(angles) | 1.67 | 1.79 |
| Ramachandran: | ||
| favored (%) | 94.51 | 97.92 |
| allowed (%) | 5.49 | 2.08 |
| outliers (%) | 0.00 | 0.00 |
| Rotamer outliers (%) | 3.08 | 0.66 |
| Clashscore | 3.25 | 3.79 |
| Average B-factor | 93.40 | 18.61 |
| macromolecules | 93.45 | 17.46 |
| ligands | 102.35 | 23.00 |
| solvent | 70.24 | 26.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porfetye, A.T.; Stege, P.; Rebollido-Rios, R.; Hoffmann, D.; Schrader, T.; Vetter, I.R. How Do Molecular Tweezers Bind to Proteins? Lessons from X-ray Crystallography. Molecules 2024, 29, 1764. https://doi.org/10.3390/molecules29081764
Porfetye AT, Stege P, Rebollido-Rios R, Hoffmann D, Schrader T, Vetter IR. How Do Molecular Tweezers Bind to Proteins? Lessons from X-ray Crystallography. Molecules. 2024; 29(8):1764. https://doi.org/10.3390/molecules29081764
Chicago/Turabian StylePorfetye, Arthur T., Patricia Stege, Rocio Rebollido-Rios, Daniel Hoffmann, Thomas Schrader, and Ingrid R. Vetter. 2024. "How Do Molecular Tweezers Bind to Proteins? Lessons from X-ray Crystallography" Molecules 29, no. 8: 1764. https://doi.org/10.3390/molecules29081764
APA StylePorfetye, A. T., Stege, P., Rebollido-Rios, R., Hoffmann, D., Schrader, T., & Vetter, I. R. (2024). How Do Molecular Tweezers Bind to Proteins? Lessons from X-ray Crystallography. Molecules, 29(8), 1764. https://doi.org/10.3390/molecules29081764