

Royal Jelly Enhances the Ability of Myoblast C2C12 Cells to Differentiate into Multilineage Cells

 , , , and
, , , and
Abstract

1. Introduction
2. Results
2.1. pRJ Stimulates BMP-Induced Osteoblast Lineage Conversion in C2C12 Cells
2.2. pRJ Stimulates Adipocyte Lineage Conversion in C2C12 Cells
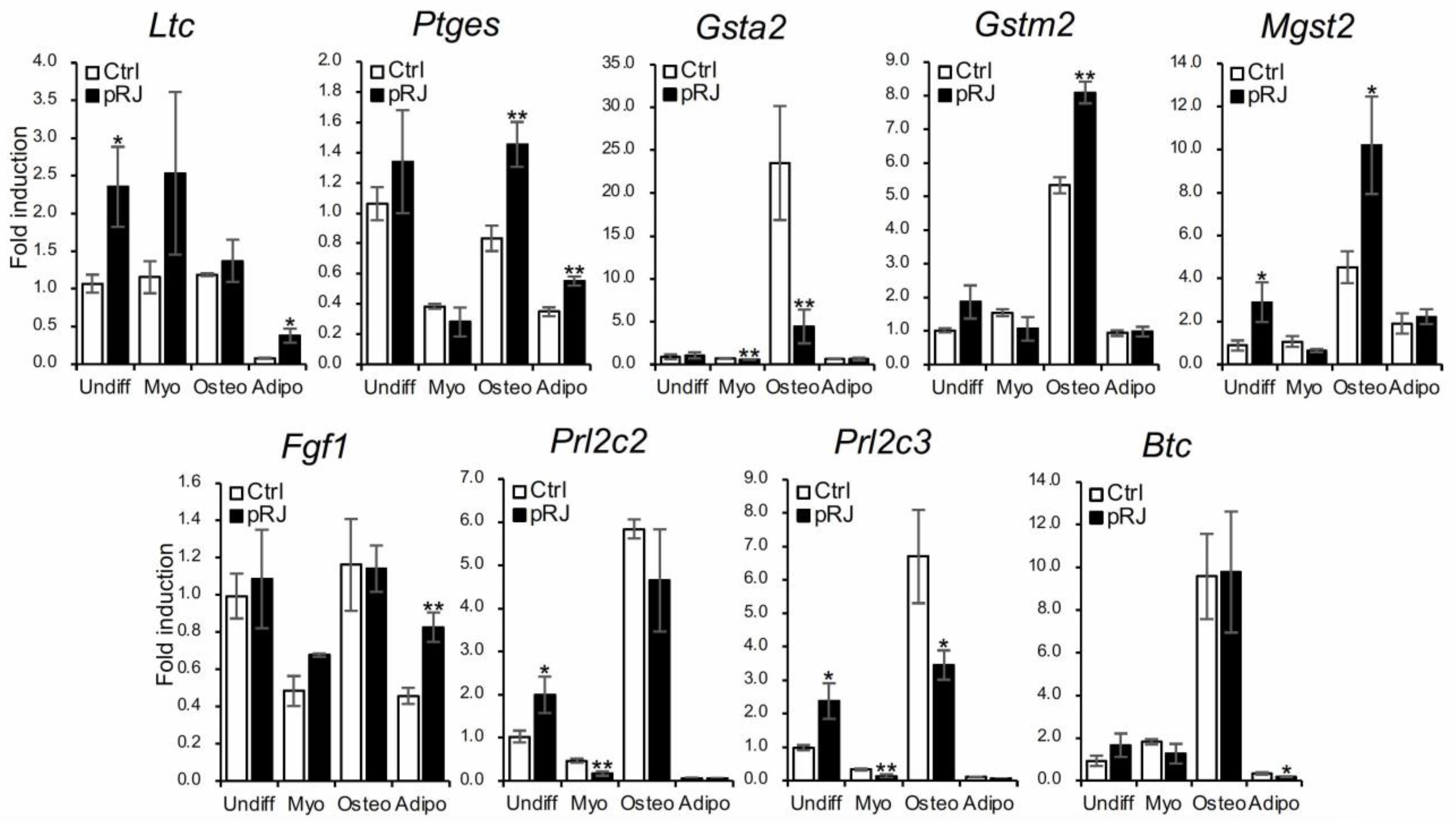
2.3. pRJ Regulated 279 Genes in C2C12 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Assay
4.3. Osteoblast Differentiation and ALP Activity Assay
4.4. Immunocytochemistry Analysis
4.5. Western Blot Analyses
4.6. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.7. Adipogenic Medium and Oil Red O Staining
4.8. Luciferase Assay
4.9. RNA Isolation and RNA-Seq Analysis
4.10. Gene-Expression Analysis
4.11. Determination of Differentially Expressed Genes
4.12. Differential Gene GO Enrichment Analysis
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isidorov, V.A.; Bakier, S.; Grzech, I. Gas chromatographic-mass spectrometric investigation of volatile and extractable compounds of crude royal jelly. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 885–886, 109–116. [Google Scholar] [CrossRef]
- Ahmad, S.; Campos, M.G.; Fratini, F.; Altaye, S.Z.; Li, J. New Insights into the Biological and Pharmaceutical Properties of Royal Jelly. Int. J. Mol. Sci. 2020, 21, 382. [Google Scholar] [CrossRef]
- Khazaei, M.; Ansarian, A.; Ghanbari, E. New Findings on Biological Actions and Clinical Applications of Royal Jelly: A Review. J. Diet. Suppl. 2018, 15, 757–775. [Google Scholar] [CrossRef]
- Okumura, N.; Toda, T.; Ozawa, Y.; Watanabe, K.; Ikuta, T.; Tatefuji, T.; Hashimoto, K.; Shimizu, T. Royal Jelly Delays Motor Functional Impairment During Aging in Genetically Heterogeneous Male Mice. Nutrients 2018, 10, 1191. [Google Scholar] [CrossRef]
- Okamoto, H.; Ohkuma, A.; Kawaguchi, M.; Shigematsu, N.; Okumura, N. Royal Jelly Increases Hematopoietic Stem Cells in Peripheral Blood: A Double-Blind, Placebo-Controlled, Randomized Trial in Healthy Subjects. Evid. Based Complement. Altern. Med. 2023, 2023, 7665515. [Google Scholar] [CrossRef] [PubMed]
- Asama, T.; Matsuzaki, H.; Fukushima, S.; Tatefuji, T.; Hashimoto, K.; Takeda, T. Royal Jelly Supplementation Improves Menopausal Symptoms Such as Backache, Low Back Pain, and Anxiety in Postmenopausal Japanese Women. Evid. Based Complement. Altern. Med. 2018, 2018, 4868412. [Google Scholar] [CrossRef]
- Rosmilah, M.; Shahnaz, M.; Patel, G.; Lock, J.; Rahman, D.; Masita, A.; Noormalin, A. Characterization of major allergens of royal jelly Apis mellifera. Trop. Biomed. 2008, 25, 243–251. [Google Scholar] [PubMed]
- Mizutani, Y.; Shibuya, Y.; Takahashi, T.; Tsunoda, T.; Moriyama, T.; Seishima, M. Major royal jelly protein 3 as a possible allergen in royal jelly-induced anaphylaxis. J. Dermatol. 2011, 38, 1079–1081. [Google Scholar] [CrossRef]
- Blank, S.; Bantleon, F.I.; McIntyre, M.; Ollert, M.; Spillner, E. The major royal jelly proteins 8 and 9 (Api m 11) are glycosylated components of Apis mellifera venom with allergenic potential beyond carbohydrate-based reactivity. Clin. Exp. Allergy 2012, 42, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Yanagihara, M.; Yano, E.; Kimura, G.; Seishima, M.; Tani, H.; Kanno, T.; Nakamura-Hirota, T.; Hashimoto, K.; Tatefuji, T.; et al. Hypoallergenicity and immunological characterization of enzyme-treated royal jelly from Apis mellifera. Biosci. Biotechnol. Biochem. 2013, 77, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, T.; Miyawaki, A.; Matsubara, T.; Okumura, N.; Okamoto, H.; Nakai, N.; Rojasawasthien, T.; Morikawa, K.; Inoue, A.; Goto, A.; et al. Daily Oral Administration of Protease-Treated Royal Jelly Protects Against Denervation-Induced Skeletal Muscle Atrophy. Nutrients 2020, 12, 3089. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Era, T. Pluripotent stem cell as a source of mesenchymal stem cell. Inflamm. Regen. 2013, 33, 19–28. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Cicek, G.; Oz Bagci, F. Effects of royal jelly on the antisenescence, mitochondrial viability and osteogenic differentiation capacity of umbilical cord-derived mesenchymal stem cells. Histochem. Cell Biol. 2023, in press. [Google Scholar] [CrossRef]
- Itoh, T.; Degawa, T.; Ito, Y.; Akao, Y.; Okumura, N.; Bassukas, I.D. Role of Royal Jelly Treated Adipose-Derived Stem Cell-Extracellular Vesicles on Fibroblast Proliferation, Migration, and Collagen Production. Dermatol. Ther. 2023, 2023, 7950026. [Google Scholar] [CrossRef]
- Yaffe, D.; Saxel, O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scime, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Imada, M.; Yanai, T.; Suda, T.; Takahashi, N.; Kamijo, R. Identification of a BMP-responsive element in Id1, the gene for inhibition of myogenesis. Genes Cells 2002, 7, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Addison, W.N.; Fu, M.M.; Yang, H.X.; Lin, Z.; Nagano, K.; Gori, F.; Baron, R. Direct transcriptional repression of Zfp423 by Zfp521 mediates a bone morphogenic protein-dependent osteoblast versus adipocyte lineage commitment switch. Mol. Cell Biol. 2014, 34, 3076–3085. [Google Scholar] [CrossRef]
- Maleszka, R. Epigenetic integration of environmental and genomic signals in honey bees: The critical interplay of nutritional, brain and reproductive networks. Epigenetics 2008, 3, 188–192. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, L.; Chen, W. Recombinant bovine FGF1 promotes muscle satellite cells mitochondrial fission and proliferation in serum-free conditions. Food Res. Int. 2024, 175, 113794. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gaviro, M.V.; Scott, C.E.; Sesay, A.K.; Matheu, A.; Booth, S.; Galichet, C.; Lovell-Badge, R. Betacellulin promotes cell proliferation in the neural stem cell niche and stimulates neurogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Kamakura, M. Royalactin induces queen differentiation in honeybees. Nature 2011, 473, 478–483. [Google Scholar] [CrossRef]
- Wan, D.C.; Morgan, S.L.; Spencley, A.L.; Mariano, N.; Chang, E.Y.; Shankar, G.; Luo, Y.; Li, T.H.; Huh, D.; Huynh, S.K.; et al. Honey bee Royalactin unlocks conserved pluripotency pathway in mammals. Nat. Commun. 2018, 9, 5078. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef]
- Okumura, N.; Ito, T.; Degawa, T.; Moriyama, M.; Moriyama, H. Royal Jelly Protects against Epidermal Stress through Upregulation of the NQO1 Expression. Int. J. Mol. Sci. 2021, 22, 12973. [Google Scholar] [CrossRef]
- Martinello, M.; Mutinelli, F. Antioxidant Activity in Bee Products: A Review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef]
- Kuo, P.L. Myricetin inhibits the induction of anti-Fas IgM-, tumor necrosis factor-alpha- and interleukin-1beta-mediated apoptosis by Fas pathway inhibition in human osteoblastic cell line MG-63. Life Sci. 2005, 77, 2964–2976. [Google Scholar] [CrossRef]
- Spilmont, M.; Leotoing, L.; Davicco, M.J.; Lebecque, P.; Mercier, S.; Miot-Noirault, E.; Pilet, P.; Rios, L.; Wittrant, Y.; Coxam, V. Pomegranate seed oil prevents bone loss in a mice model of osteoporosis, through osteoblastic stimulation, osteoclastic inhibition and decreased inflammatory status. J. Nutr. Biochem. 2013, 24, 1840–1848. [Google Scholar] [CrossRef]
- Shavandi, A.; Hou, Y.; Carne, A.; McConnell, M.; Bekhit, A.E.A. Marine Waste Utilization as a Source of Functional and Health Compounds. Adv. Food Nutr. Res. 2019, 87, 187–254. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Barbieri, E.; Martinelli, C.; Battistelli, M.; Guescini, M.; Vallorani, L.; Casadei, L.; D’Emilio, A.; Falcieri, E.; Piccoli, G.; et al. Creatine supplementation prevents the inhibition of myogenic differentiation in oxidatively injured C2C12 murine myoblasts. Mol. Nutr. Food Res. 2009, 53, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Yamada, M.; Niibe, K.; Zhang, M.; Kondo, T.; Ishibashi, M.; Egusa, H. Preconditioning of bone marrow-derived mesenchymal stem cells with N-acetyl-L-cysteine enhances bone regeneration via reinforced resistance to oxidative stress. Biomaterials 2018, 185, 25–38. [Google Scholar] [CrossRef] [PubMed]
- de Villiers, D.; Potgieter, M.; Ambele, M.A.; Adam, L.; Durandt, C.; Pepper, M.S. The Role of Reactive Oxygen Species in Adipogenic Differentiation. Adv. Exp. Med. Biol. 2018, 1083, 125–144. [Google Scholar] [CrossRef] [PubMed]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Kokabu, S.; Nguyen, T.; Ohte, S.; Sato, T.; Katagiri, T.; Yoda, T.; Rosen, V. TLE3, transducing-like enhancer of split 3, suppresses osteoblast differentiation of bone marrow stromal cells. Biochem. Biophys. Res. Commun. 2013, 438, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | GO Term | GO ID | Gene Count | Gene Symbol |
|---|---|---|---|---|
| Matured muscle Muscle contraction | Actin binding | GO:0003779 | 11 | Cnn1, Myh11, Myh3, Parvg, Avil, Tnni1, Tagln, Lmod1, Myh7, Myh1, Tnnt3 |
| Cytoskeleton | GO:0005856 | 8 | Cnn1, Parvg, Cabyr, Avil, Lmod1, Actg2, Actc1, Gm2420 | |
| Motor activity | GO:0003774 | 4 | Myh11, Myh3, Myh7, Myh1 | |
| Muscle–myosin complex | GO:0005859 | 5 | Myh11, Mylpf, Myh7, Myh1, Myl9 | |
| Structural constituent of muscle | GO:0008307 | 4 | Myh11, Mylpf, Myh7, Myl9 | |
| Troponin complex | GO:0005861 | 3 | Tnnc2, Tnni1, Tnnt3 | |
| Sarcomere assembly | Z disc | GO:0030018 | 3 | Casq1 Cacna1s Actc1 |
| I band | GO:0031674 | 15 | Pvalb, Casq1, Tnnc2, Cabyr, Vldlr, Fbln7, Cabp1, Mylpf, Try5, Pcdhb12, Pcdhb4, Mmp13, Myl9 | |
| Calcium related | Calcium ion binding | GO:0005509 | 5 | Vldlr, Cabp1, Mmp13, Tnnt3, Gm15720 |
| Calcium-dependent protein binding | GO:0048306 | 5 | Cnn1, Myh11, Myh3, Myh7, Myh1 | |
| Calcium-mediated signaling | GO:0005516 | 3 | Edn1, Cabyr, Gm15720 | |
| Immune response | Immune response | GO:0019722 | 3 | Prg4, Mylpf, Cxcl10 |
| Cellular response to interleukin-1 | GO:0071347 | 3 | Edn1, Vldlr, Ccl5 |
| Classification | GO Term | GO ID | Gene Count | Gene Symbol |
|---|---|---|---|---|
| Glutathione related | Glutathione binding | GO:00 | 5 | Ltc4s, Ptges, Gsta2, Gstm2-ps1, Gm3776 |
| Glutathione transferase activity | GO:0004364 | 5 | Ltc4s, Gsta2, Mgst2, Gstm2-ps1, Gm3776 | |
| Glutathione metabolic process | GO:0006749 | 3 | Gsta2, Gstm2-ps1, Gm3776 | |
| Growth factor activity | Growth factor activity | GO:0008083 | 4 | Fgf1, Prl2c3, Prl2c2, Btc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, T.; Rojasawasthien, T.; Takeuchi, S.Y.; Okamoto, H.; Okumura, N.; Shirakawa, T.; Matsubara, T.; Kawamoto, T.; Kokabu, S. Royal Jelly Enhances the Ability of Myoblast C2C12 Cells to Differentiate into Multilineage Cells. Molecules 2024, 29, 1449. https://doi.org/10.3390/molecules29071449
Ito T, Rojasawasthien T, Takeuchi SY, Okamoto H, Okumura N, Shirakawa T, Matsubara T, Kawamoto T, Kokabu S. Royal Jelly Enhances the Ability of Myoblast C2C12 Cells to Differentiate into Multilineage Cells. Molecules. 2024; 29(7):1449. https://doi.org/10.3390/molecules29071449
Chicago/Turabian StyleIto, Takumi, Thira Rojasawasthien, Sachiko Yamashita Takeuchi, Hideto Okamoto, Nobuaki Okumura, Tomohiko Shirakawa, Takuma Matsubara, Tatsuo Kawamoto, and Shoichiro Kokabu. 2024. "Royal Jelly Enhances the Ability of Myoblast C2C12 Cells to Differentiate into Multilineage Cells" Molecules 29, no. 7: 1449. https://doi.org/10.3390/molecules29071449
APA StyleIto, T., Rojasawasthien, T., Takeuchi, S. Y., Okamoto, H., Okumura, N., Shirakawa, T., Matsubara, T., Kawamoto, T., & Kokabu, S. (2024). Royal Jelly Enhances the Ability of Myoblast C2C12 Cells to Differentiate into Multilineage Cells. Molecules, 29(7), 1449. https://doi.org/10.3390/molecules29071449