
Plant-Derived Phenolic Acids Limit the Pathogenesis of Salmonella Typhimurium and Protect Intestinal Epithelial Cells during Their Interactions


Abstract

1. Introduction
2. Results
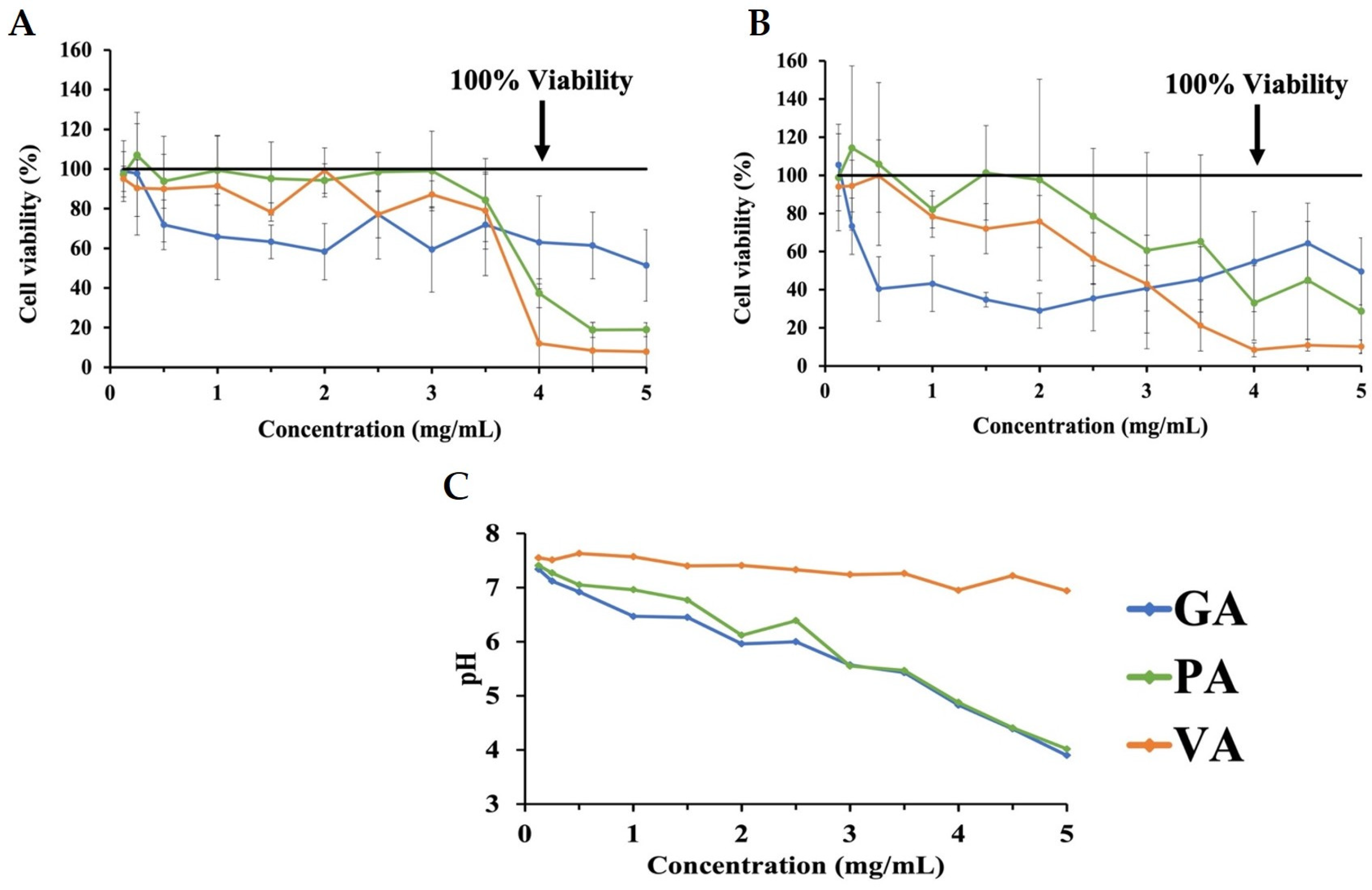
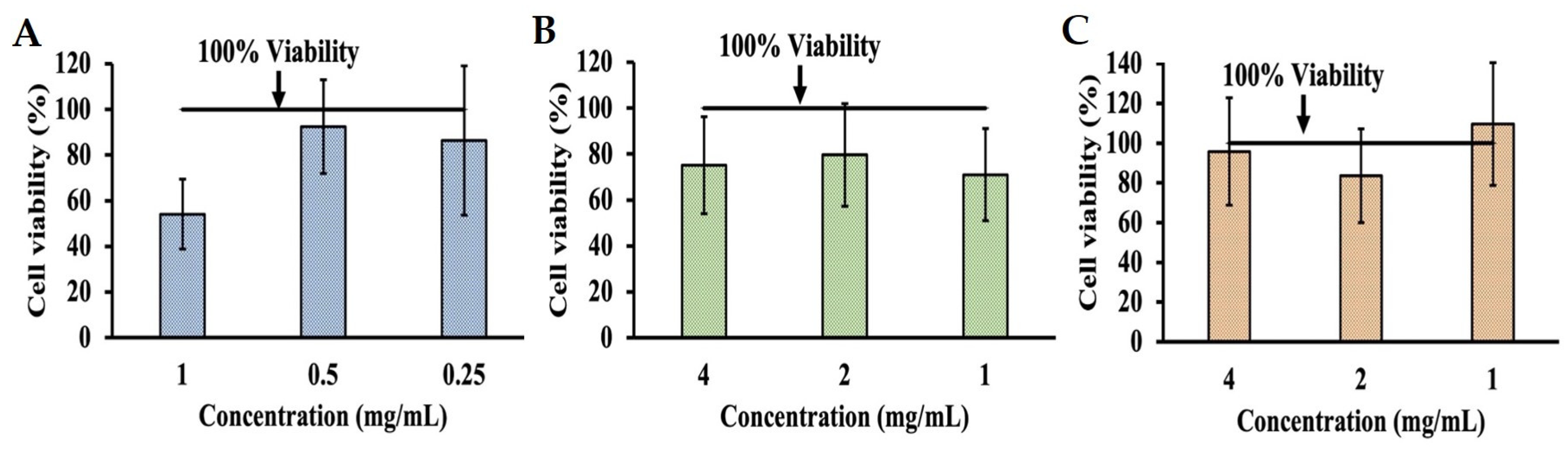
2.1. Evaluation of Cytotoxicity of Phenolic Acids in INT-407 Cells Using MTT Assay
2.2. Determining Effects of pH on Cytotoxicity in INT-407 Cell Using MTT
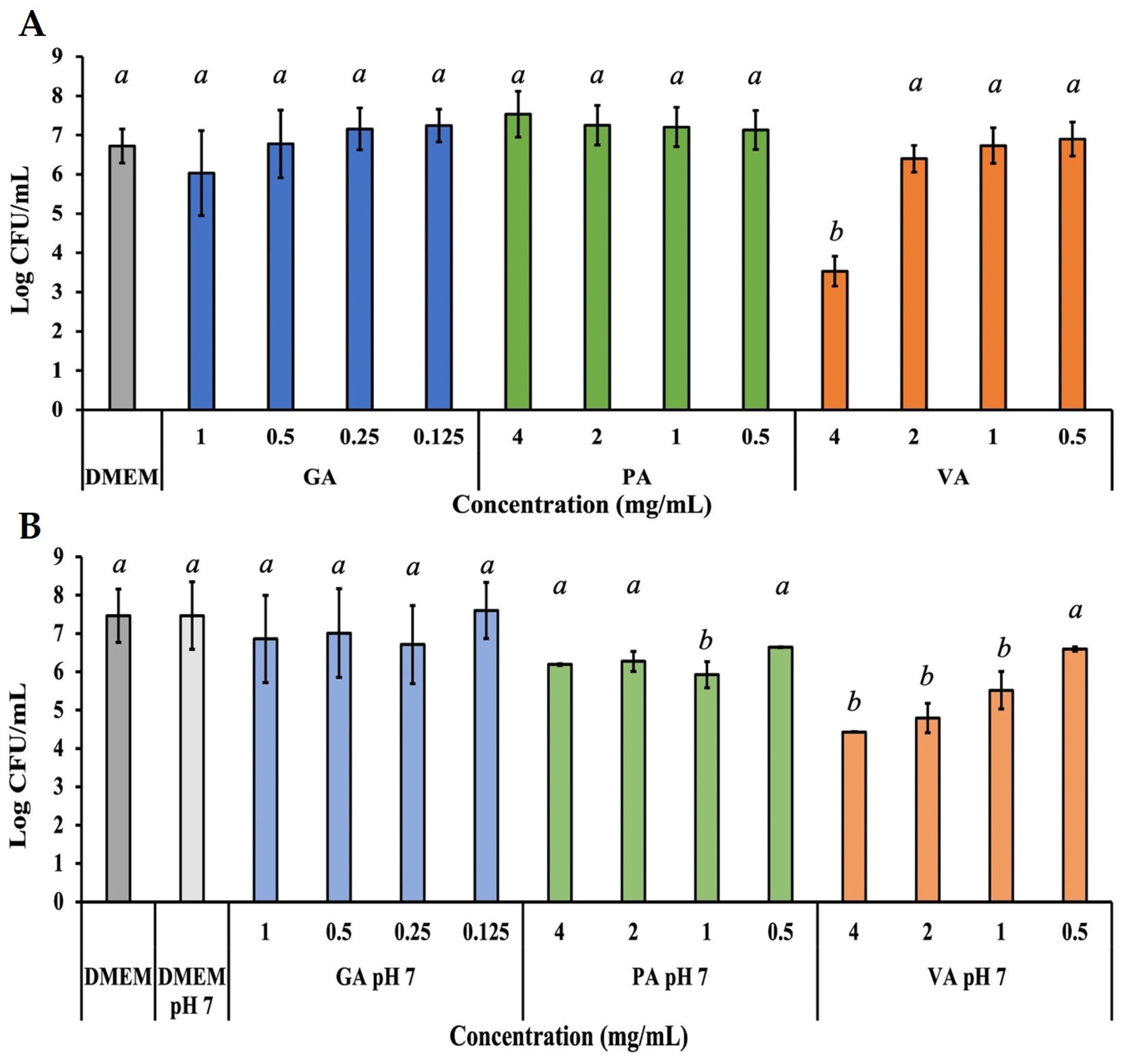
2.3. Antimicrobial Potency of Phenolic Acids in DMEM
2.4. Changes in Host-Pathogen Interactions between ST and INT-407 after Treatment
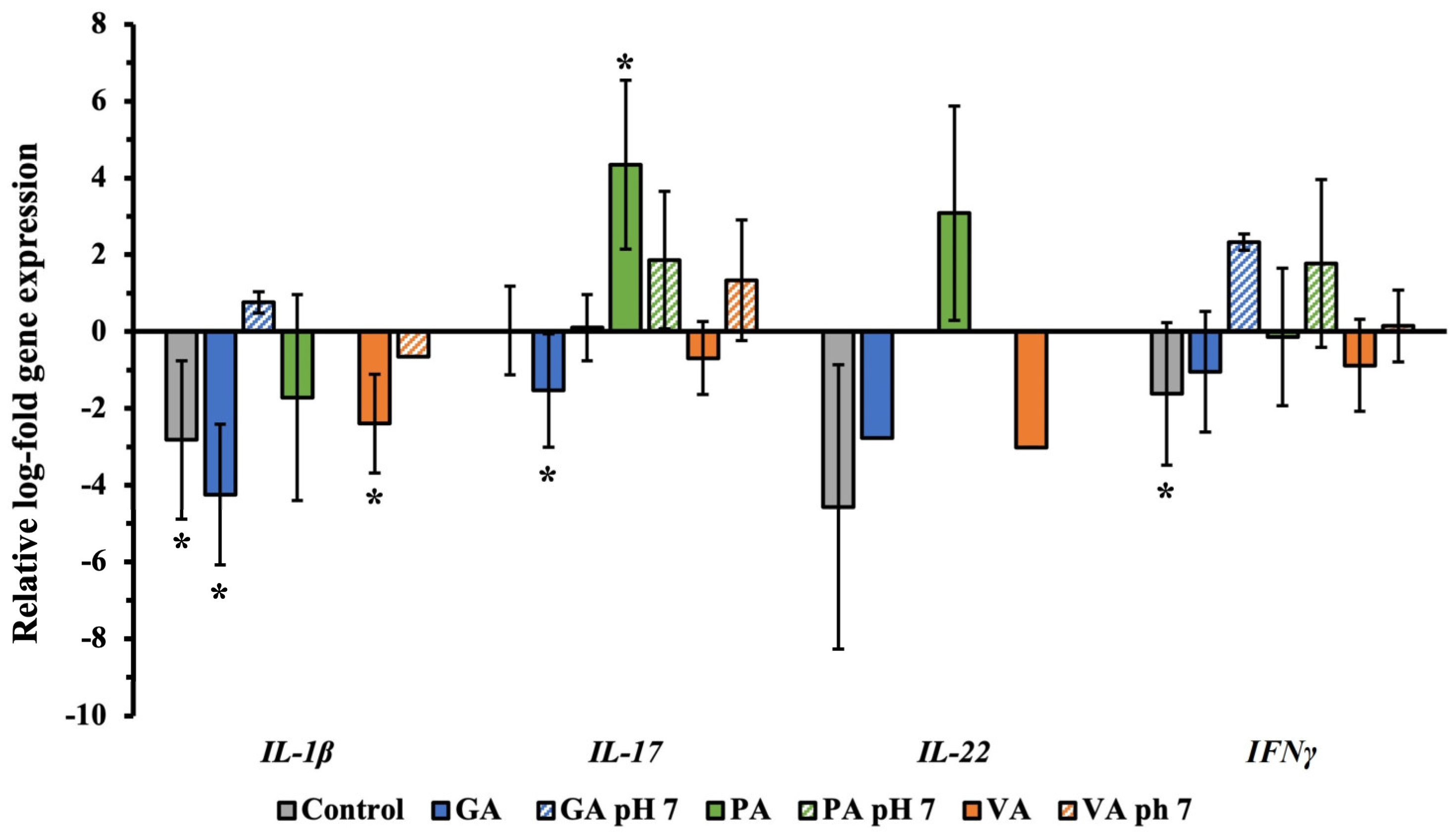
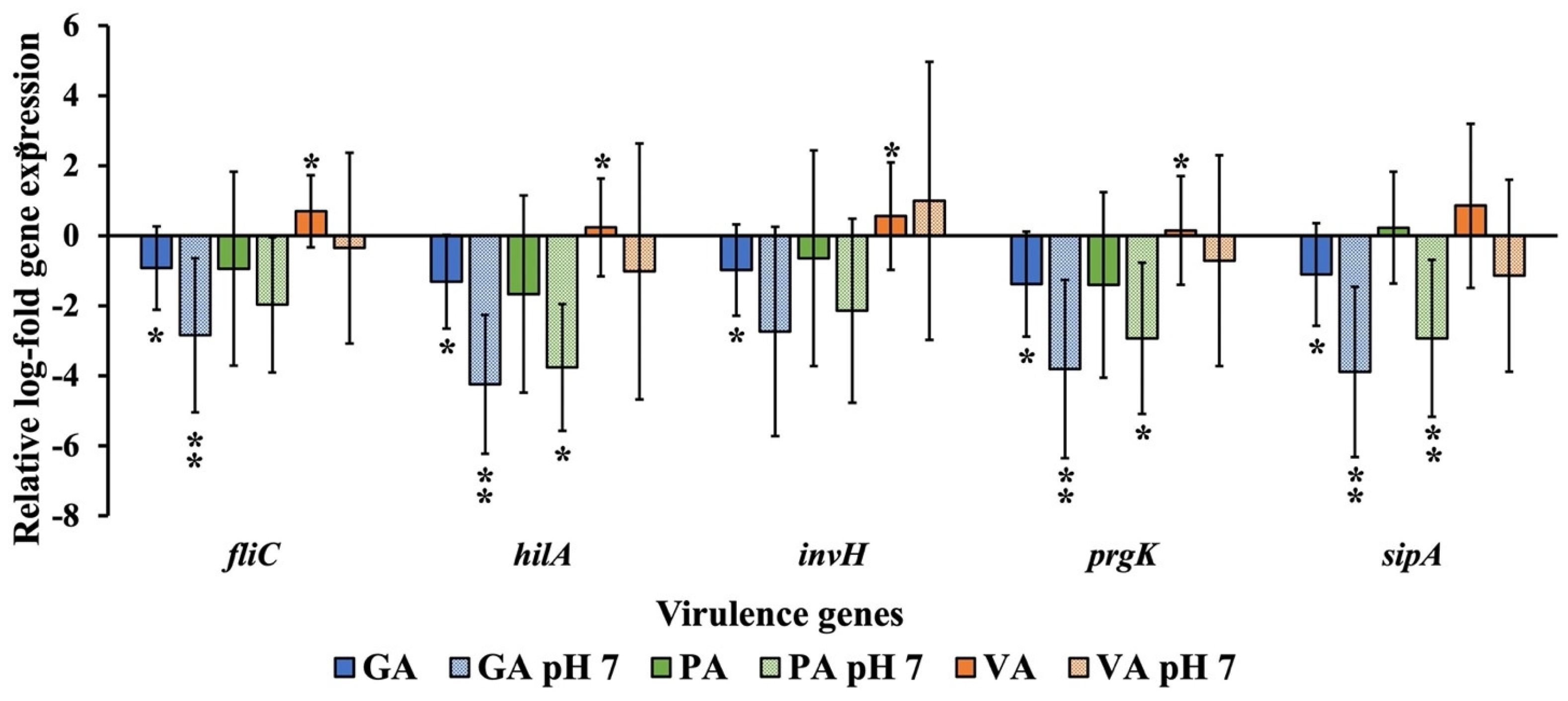
2.5. Relative Gene Expression of Virulence Genes and Inflammatory Cytokine Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Growth Conditions
4.2. Cell Line and Growth Conditions
4.3. Compounds and Stock Solution Preparation
4.4. Phenolic Acid Antimicrobial Potential within DMEM and at Alternate pH Ranges
4.5. MTT Assay for Evaluating Cytotoxicity and Cell Viability
4.6. Adhesion and Invasion Assay
4.7. Assessing Relative Expression in INT-407 Cells and ST
4.8. Measuring Gene Expression through qRT-PCR Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stanaway, J.D.; Parisi, A.; Sarkar, K.; Blacker, B.F.; Reiner, R.C.; Hay, S.I.; Nixon, M.R.; Dolecek, C.; James, S.L.; Mokdad, A.H.; et al. The Global Burden of Non-Typhoidal Salmonella Invasive Disease: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States-Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Interagency Food Safety Analytics Collaboration. Foodborne Illness Source Attribution Estimates for 2020 for Salmonella, Escherichia coli O157, and Listeria Monocytogenes Using Multi-Year Outbreak Surveillance Data, United States; Interagency Food Safety Analytics Collaboration (IFSAC): Stillwater, OK, USA, 2022; p. 157. [Google Scholar]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, Colonization, and Virulence Differences among Serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills to Succeed in the Host: Virulence and Regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-Associated Salmonella enterica Serotypes and Food Commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic Resistance in Salmonella Typhimurium Isolates Recovered from the Food Chain through National Antimicrobial Resistance Monitoring System between 1996 and 2016. Front. Microbiol. 2019, 10, 985. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Surveillance for Foodborne Disease Outbreaks, United States, 2017; Annual Report; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2019. [Google Scholar]
- Chen, H.M.; Wang, Y.; Su, L.H.; Chiu, C.H. Nontyphoid Salmonella Infection: Microbiology, Clinical Features, and Antimicrobial Therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Nyirenda, T.S.; Kalonji, L.M.; Phoba, M.-F.; Tack, B.; Platts-Mills, J.A.; Gordon, M.A.; Kariuki, S.M. Nontyphoidal Salmonella Invasive Disease: Challenges and Solutions. Open Forum Infect. Dis. 2023, 10, S32–S37. [Google Scholar] [CrossRef]
- Salaheen, S.; Tabashsum, Z.; Gaspard, S.; Dattilio, A.; Tran, T.H.; Biswas, D. Reduced Campylobacter jejuni Colonization in Poultry Gut with Bioactive Phenolics. Food Control 2018, 84, 1–7. [Google Scholar] [CrossRef]
- Salaheen, S.; Kim, S.W.; Haley, B.J.; Van Kessel, J.A.S.; Biswas, D. Alternative Growth Promoters Modulate Broiler Gut Microbiome and Enhance Body Weight Gain. Front. Microbiol. 2017, 8, 2088. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Anthony Ryan, C.; Dempsey, E.; Paul Ross, R.; Stanton, C. Impact of Antibiotics on the Human Microbiome and Consequences for Host Health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Ecevit, K.; Barros, A.A.; Silva, J.M.; Reis, R.L. Preventing Microbial Infections with Natural Phenolic Compounds. Futur. Pharmacol. 2022, 2, 460–498. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Nat. Prod. Commun. 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic Acids: Natural Versatile Molecules with Promising Therapeutic Applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Beya, M.M.; Netzel, M.E.; Sultanbawa, Y.; Smyth, H.; Hoffman, L.C. Plant-Based Phenolic Molecules as Natural Preservatives in Comminuted Meats: A Review. Antioxidants 2021, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Martinez, Z.; Bravo, P.; Kennedy, N.F.; Krishna, M.; Hussain, S.; Young, A.C.; Biswas, D. Antimicrobial and Antivirulence Impacts of Phenolics on Salmonella enterica Serovar Typhimurium. Antibiotics 2020, 9, 668. [Google Scholar] [CrossRef]
- Ménard, S.; Lacroix-Lamandé, S.; Ehrhardt, K.; Yan, J.; Grassl, G.A.; Wiedemann, A. Cross-Talk Between the Intestinal Epithelium and Salmonella Typhimurium. Front. Microbiol. 2022, 13, 906238. [Google Scholar] [CrossRef]
- Diacovich, L.; Lorenzi, L.; Tomassetti, M.; Méresse, S.; Gramajo, H. The Infectious Intracellular Lifestyle of Salmonella enterica Relies on the Adaptation to Nutritional Conditions within the Salmonella-Containing Vacuole. Virulence 2016, 8, 975–992. [Google Scholar] [CrossRef]
- Chong, A.; Starr, T.; Finn, C.E.; Steele-Mortimer, O. A Role for the Salmonella Type III Secretion System 1 in Bacterial Adaptation to the Cytosol of Epithelial Cells. Mol. Microbiol. 2019, 112, 1270–1283. [Google Scholar] [CrossRef]
- Bao, H.; Wang, S.; Zhao, J.H.; Liu, S.L. Salmonella Secretion Systems: Differential Roles in Pathogen-Host Interactions. Microbiol. Res. 2020, 241, 1265911. [Google Scholar] [CrossRef] [PubMed]
- Furter, M.; Sellin, M.E.; Hansson, G.C.; Hardt, W.D. Mucus Architecture and Near-Surface Swimming Affect Distinct Salmonella Typhimurium Infection Patterns along the Murine Intestinal Tract. Cell Rep. 2019, 27, 2665–2678.e3. [Google Scholar] [CrossRef] [PubMed]
- Birhanu, B.T.; Park, N.H.; Lee, S.J.; Hossain, M.A.; Park, S.C. Inhibition of Salmonella Typhimurium Adhesion, Invasion, and Intracellular Survival via Treatment with Methyl Gallate Alone and in Combination with Marbofloxacin. Vet. Res. 2018, 49, 101. [Google Scholar] [CrossRef] [PubMed]
- Lara-Tejero, M.; Galán, J.E. Salmonella enterica serovar Typhimurium Pathogenicity Island 1-Encoded Type III Secretion System Translocases Mediate Intimate Attachment to Nonphagocytic Cells. Infect. Immun. 2009, 77, 2635–2642. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef]
- Cos, P.; Rajan, P.; Vedernikova, I.; Calomme, M.; Pieters, L.; Vlietinck, A.J.; Augustyns, K.; Haemers, A.; Berghe, D. Vanden In Vitro Antioxidant Profile of Phenolic Acid Derivatives. Free. Radic. Res. 2002, 36, 711–716. [Google Scholar] [CrossRef]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Rivas, C.L.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial Activity of Phenolic Acids against Commensal, Probiotic and Pathogenic Bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Daliri, E.B.M.; Elahi, F.; Chelliah, R.; Lee, B.H.; Oh, D.H. New Insights on the Use of Polyphenols as Natural Preservatives and Their Emerging Safety Concerns. Front. Sustain. Food Syst. 2020, 4, 525810. [Google Scholar] [CrossRef]
- Salaheen, S.; Peng, M.; Joo, J.; Teramoto, H.; Biswas, D. Eradication and Sensitization of Methicillin Resistant Staphylococcus aureus to Methicillin with Bioactive Extracts of Berry Pomace. Front. Microbiol. 2017, 8, 253. [Google Scholar] [CrossRef]
- Hossain, M.A.; Park, H.C.; Lee, K.J.; Park, S.W.; Park, S.C.; Kang, J. In Vitro Synergistic Potentials of Novel Antibacterial Combination Therapies against Salmonella enterica serovar Typhimurium. BMC Microbiol. 2020, 20, 118. [Google Scholar] [CrossRef] [PubMed]
- Balta, I.; Linton, M.; Pinkerton, L.; Kelly, C.; Stef, L.; Pet, I.; Stef, D.; Criste, A.; Gundogdu, O.; Corcionivoschi, N. The Effect of Natural Antimicrobials against Campylobacter spp. and Its Similarities to Salmonella spp., Listeria spp., Escherichia coli, Vibrio spp., Clostridium spp. and Staphylococcus spp. Food Control 2021, 121, 107745. [Google Scholar] [CrossRef]
- Birhanu, B.T.; Lee, E.B.; Lee, S.J.; Park, S.C. Targeting Salmonella Typhimurium Invasion and Intracellular Survival Using Pyrogallol. Front. Microbiol. 2021, 12, 631426. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, A.; Hardt, W.-D. The Interplay between Salmonella enterica serovar Typhimurium and the Intestinal Mucosa during Oral Infection. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Ribet, D.; Cossart, P. How Bacterial Pathogens Colonize Their Hosts and Invade Deeper Tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Tadala, L.; Langenbach, D.; Dannborg, M.; Cervantes-Rivera, R.; Sharma, A.; Vieth, K.; Rieckmann, L.M.; Wanders, A.; Cisneros, D.A.; Puhar, A. Infection-Induced Membrane Ruffling Initiates Danger and Immune Signaling via the Mechanosensor PIEZO1. Cell Rep. 2022, 40, 111173. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Martinez, Z.; Tabashsum, Z.; Aditya, A.; Suh, G.; Wall, M.; Hshieh, K.; Biswas, D. Purified Plant-Derived Phenolic Acids Inhibit Salmonella Typhimurium without Alteration of Microbiota in a Simulated Chicken Cecum Condition. Microorganisms 2023, 11, 957. [Google Scholar] [CrossRef]
- Spiegel, M.; Cel, K.; Sroka, Z. The Mechanistic Insights into the Role of PH and Solvent on Antiradical and Prooxidant Properties of Polyphenols—Nine Compounds Case Study. Food Chem. 2023, 407, 134677. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhang, Q.Q.; Zhang, J.; Song, J.L.; Li, J.C.; Han, K.; Huang, J.T.; Jiang, C.S.; Zhang, H. Synthesis and Evaluation of 1,2,4-Oxadiazole Derivatives as Potential Anti-Inflammatory Agents by Inhibiting NF-ΚB Signaling Pathway in LPS-Stimulated RAW 264.7 Cells. Bioorganic Med. Chem. Lett. 2020, 30, 127373. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, M.; Kan, Z.; Zhang, G.; Li, Y.; Zhou, L.; Wang, C. Identification of Novel Type Three Secretion System (T3SS) Inhibitors by Computational Methods and Anti-Salmonella Evaluations. Front. Pharmacol. 2021, 12, 764191. [Google Scholar] [CrossRef]
- Lathrop, S.K.; Cooper, K.G.; Binder, K.A.; Starr, T.; Mampilli, V.; Detweiler, C.S.; Steele-Mortimer, O. Salmonella Typhimurium Infection of Human Monocyte-Derived Macrophages. Curr. Protoc. Microbiol. 2018, 50, e56. [Google Scholar] [CrossRef]
- Lhocine, N.; Arena, E.T.; Bomme, P.; Ubelmann, F.; Prévost, M.C.; Robine, S.; Sansonetti, P.J. Apical Invasion of Intestinal Epithelial Cells by Salmonella Typhimurium Requires Villin to Remodel the Brush Border Actin Cytoskeleton. Cell Host Microbe 2015, 17, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Dostal, A.; Gagnon, M.; Chassard, C.; Zimmermann, M.B.; O’Mahony, L.; Lacroix, C. Salmonella Adhesion, Invasion and Cellular Immune Responses Are Differentially Affected by Iron Concentrations in a Combined in Vitro Gut Fermentation-Cell Model. PLoS ONE 2014, 9, e93549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Wang, T.; Deng, X.; Chu, X. Natural Compound Sanguinarine Chloride Targets the Type III Secretion System of Salmonella enterica serovar Typhimurium. Biochem. Biophys. Rep. 2018, 14, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Allam, U.S.; Gopala Krishna, M.; Sen, M.; Thomas, R.; Lahiri, A.; Gnanadhas, D.P.; Chakravortty, D. Acidic PH Induced STM1485 Gene Is Essential for Intracellular Replication of Salmonella. Virulence 2012, 3, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Beagley, K.W.; France, M.P.; Shen, J.; Husband, A.J. Interferon-γ Plays a Critical Role in Intestinal Immunity against Salmonella Typhimurium Infection. Immunology 2000, 99, 464–472. [Google Scholar] [CrossRef]
- Mayuzumi, H.; Inagaki-Ohara, K.; Uyttenhove, C.; Okamoto, Y.; Matsuzaki, G. Interleukin-17A Is Required to Suppress Invasion of Salmonella enterica serovar Typhimurium to Enteric Mucosa. Immunology 2010, 131, 377–385. [Google Scholar] [CrossRef]
- Lo, B.C.; Shin, S.B.; Canals Hernaez, D.; Refaeli, I.; Yu, H.B.; Goebeler, V.; Cait, A.; Mohn, W.W.; Vallance, B.A.; McNagny, K.M. IL-22 Preserves Gut Epithelial Integrity and Promotes Disease Remission during Chronic Salmonella Infection. J. Immunol. 2019, 202, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Lutay, N.; Ambite, I.; Hernandez, J.G.; Rydström, G.; Ragnarsdóttir, B.; Puthia, M.; Nadeem, A.; Zhang, J.; Storm, P.; Dobrindt, U.; et al. Bacterial Control of Host Gene Expression through RNA Polymerase II. J. Clin. Investig. 2013, 123, 2366–2379. [Google Scholar] [CrossRef]
- Diamond, C.E.; Leong, K.W.K.; Vacca, M.; Rivers-Auty, J.; Brough, D.; Mortellaro, A. Salmonella Typhimurium-Induced IL-1 Release from Primary Human Monocytes Requires NLRP3 and Can Occur in the Absence of Pyroptosis. Sci. Rep. 2017, 7, 6861. [Google Scholar] [CrossRef]
- Bent, R.; Moll, L.; Grabbe, S.; Bros, M. Interleukin-1 Beta—A Friend or Foe in Malignancies? Int. J. Mol. Sci. 2018, 19, 2155. [Google Scholar] [CrossRef]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef]
- Jepson, M.A.; Kenny, B.; Leard, A.D. Role of SipA in the Early Stages of Salmonella Typhimurium Entry into Epithelial Cells. Cell. Microbiol. 2001, 3, 417–426. [Google Scholar] [CrossRef]
- Kimbrough, T.G.; Miller, S.I. Contribution of Salmonella Typhimurium Type III Secretion Components to Needle Complex Formation. Proc. Natl. Acad. Sci. USA 2000, 97, 11008–11013. [Google Scholar] [CrossRef]
- Pati, N.B.; Vishwakarma, V.; Jaiswal, S.; Periaswamy, B.; Hardt, W.D.; Suar, M. Deletion of InvH Gene in Salmonella enterica Serovar Typhimurium Limits the Secretion of Sip Effector Proteins. Microbes Infect. 2013, 15, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Cummings, L.A.; Wilkerson, W.D.; Bergsbaken, T.; Cookson, B.T. In Vivo, FliC Expression by Salmonella enterica Serovar Typhimurium Is Heterogeneous, Regulated by ClpX, and Anatomically Restricted. Mol. Microbiol. 2006, 61, 795–809. [Google Scholar] [CrossRef] [PubMed]
- CLSI M100; Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed. Clinical & Laboratory Standards: Wayne, PA, USA, 2023.
- Salaheen, S.; Jaiswal, E.; Joo, J.; Peng, M.; Ho, R.; OConnor, D.; Adlerz, K.; Aranda-Espinoza, J.H.; Biswas, D. Bioactive Extracts from Berry Byproducts on the Pathogenicity of Salmonella Typhimurium. Int. J. Food Microbiol. 2016, 237, 128–135. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot095505. [Google Scholar] [CrossRef]
- Tolosa, L.; Donato, M.T.; Gómez-Lechón, M.J. General Cytotoxicity Assessment by Means of the MTT Assay. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 1250. [Google Scholar]
- Cannella, V.; Altomare, R.; Chiaramonte, G.; Di Bella, S.; Mira, F.; Russotto, L.; Pisano, P.; Guercio, A. Cytotoxicity Evaluation of Endodontic Pins on L929 Cell Line. BioMed Res. Int. 2019, 2019, 3469525. [Google Scholar] [CrossRef]
- Peng, M.; Reichmann, G.; Biswas, D. Lactobacillus casei and Its Byproducts Alter the Virulence Factors of Foodborne Bacterial Pathogens. J. Funct. Foods 2015, 15, 418–428. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). Cold Spring Harb. Protoc. 2010, 6, pdb.prot5439. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.K.; Kaiser, P.; Rothwell, L.; Humphrey, T.; Barrow, P.A.; Jones, M.A. Campylobacter jejuni-Induced Cytokine Responses in Avian Cells. Infect. Immun. 2005, 73, 2094–2100. [Google Scholar] [CrossRef]
- Rothwell, L.; Young, J.R.; Zoorob, R.; Whittaker, C.A.; Hesketh, P.; Archer, A.; Smith, A.L.; Kaiser, P. Cloning and Characterization of Chicken IL-10 and Its Role in the Immune Response to Eimeria Maxima. J. Immunol. 2004, 173, 2675–2682. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Lee, S.H.; Dalloul, R.A.; Lillehoj, E.P. Analysis of Chicken Cytokine and Chemokine Gene Expression Following Eimeria Acervulina and Eimeria Tenella Infections. Vet. Immunol. Immunopathol. 2006, 114, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Tabashsum, Z.; Patel, P.; Bernhardt, C.; Biswas, D. Linoleic Acids Overproducing Lactobacillus casei Limits Growth, Survival, and Virulence of Salmonella Typhimurium and Enterohaemorrhagic Escherichia coli. Front. Microbiol. 2018, 9, 2663. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Concentration (mg/mL) | |||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DMEM | Gallic Acid | Protocatechuic Acid | Vanillic Acid | |||||||||||||||||||||||||||||||||||||
| 0 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 | 4.5 | 5 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 | 4.5 | 5 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 1.5 | 2 | 2.5 | 3 | 3.5 | 4 | 4.5 | 5 | |
| 3 | + | + | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | + | + | + | + | - | - | - | - | - | - |
| 4 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | - | - | - | - | - | - | - | + | + | + | + | + | + | + | - | - | - | - | - | - |
| 5 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | - | - | - | - | - | + | + | + | + | + | + | + | - | - | - | - | - | - |
| 6 | + | + | + | + | + | + | - | - | - | - | - | - | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | - |
| 7 | + | + | + | - | - | - | - | - | - | - | - | - | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | + |
| 8 | + | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | + | + | - | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | - |
| 9 | + | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | + | - | - | - | - | + | - | - | - | - | + | + | + | + | + | + | + | + | + | + | - | - | - |
| 10 | + | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + | + | + | - | - | - | - | + | - | - | - | - | + | + | + | + | + | + | + | + | + | + | - | - | - |
| Gene | Cytokine | Primer Sequence (5′-3′) |
|---|---|---|
| GAPDH | dehydrogenase | F: GGTGGTGCTAAGCGTGTTAT |
| R: ACCTCTGTCATCTCTCCACA | ||
| IFNγ | interferon gamma | F: GTGAAGAAGGTGAAAGATATCATGGA |
| R: GCTTTGCGCTGGATTCTCA | ||
| IL-17 | neutrophil activation | F: GCAGATGACGGTACATCCAA |
| R: CCAGATCAGGCTGTGCTTTA | ||
| IL-1β | induces pyroptosis | F: GCCATGGACAAGCTGAGGAAG |
| R: GTGCTGATGTACCAGTTGGG | ||
| IL-22 | host defense at mucosa | F: CTCCGATCCCTTATTCTCCTC |
| R: AAGCGGTTGTGGTCCTCAT |
| Gene | Protein | Primer Sequence (5′-3′) |
|---|---|---|
| 16S-rRNA | 16S ribosomal RNA protein | F: GTAGTACGATGGCGAAACTGC |
| R: CTTCTCGACCCGAGGGACTT | ||
| hilA | SPI-1 transcriptional regulator | F: AATGGTCACAGGCTGAGGTG |
| R: ACATCGTCGCGACTTGTGAA | ||
| fliC | flagellum subunit | F: GCAGATGACGGTACATCCAA |
| R: CCAGATCAGGCTGTGCTTTA | ||
| invH | adherence and invasion | F: GGTGCCCCTCCCTTCCT |
| R: TGCGTTGGCCAGTTGCT | ||
| sipA | actin binding protein | F: CGTCTTCGCCTCAGGAGAAT |
| R: TGCCGGGCTCTTTCGTT | ||
| prgK | base structure formation of T3SS | F: GGGTGGAAATAGCGCAGATG |
| R: TCAGCTCGCGGAGACGATA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado-Martinez, Z.; Tabashsum, Z.; Aditya, A.; Hshieh, K.; Suh, G.; Wall, M.; Scriba, A.; Sellers, G.; Canagarajah, C.; Kapadia, S.; et al. Plant-Derived Phenolic Acids Limit the Pathogenesis of Salmonella Typhimurium and Protect Intestinal Epithelial Cells during Their Interactions. Molecules 2024, 29, 1364. https://doi.org/10.3390/molecules29061364
Alvarado-Martinez Z, Tabashsum Z, Aditya A, Hshieh K, Suh G, Wall M, Scriba A, Sellers G, Canagarajah C, Kapadia S, et al. Plant-Derived Phenolic Acids Limit the Pathogenesis of Salmonella Typhimurium and Protect Intestinal Epithelial Cells during Their Interactions. Molecules. 2024; 29(6):1364. https://doi.org/10.3390/molecules29061364
Chicago/Turabian StyleAlvarado-Martinez, Zabdiel, Zajeba Tabashsum, Arpita Aditya, Katherine Hshieh, Grace Suh, Matthew Wall, Aaron Scriba, George Sellers, Christa Canagarajah, Sarika Kapadia, and et al. 2024. "Plant-Derived Phenolic Acids Limit the Pathogenesis of Salmonella Typhimurium and Protect Intestinal Epithelial Cells during Their Interactions" Molecules 29, no. 6: 1364. https://doi.org/10.3390/molecules29061364
APA StyleAlvarado-Martinez, Z., Tabashsum, Z., Aditya, A., Hshieh, K., Suh, G., Wall, M., Scriba, A., Sellers, G., Canagarajah, C., Kapadia, S., & Biswas, D. (2024). Plant-Derived Phenolic Acids Limit the Pathogenesis of Salmonella Typhimurium and Protect Intestinal Epithelial Cells during Their Interactions. Molecules, 29(6), 1364. https://doi.org/10.3390/molecules29061364