
A Cell-Penetrating Peptide Improves Anti-HER2 Single-Chain Variable Fragment Internalization and Antitumor Activity against HER2-Positive Breast Cancer In Vitro and In Vivo

Abstract

1. Introduction
2. Results and Discussion
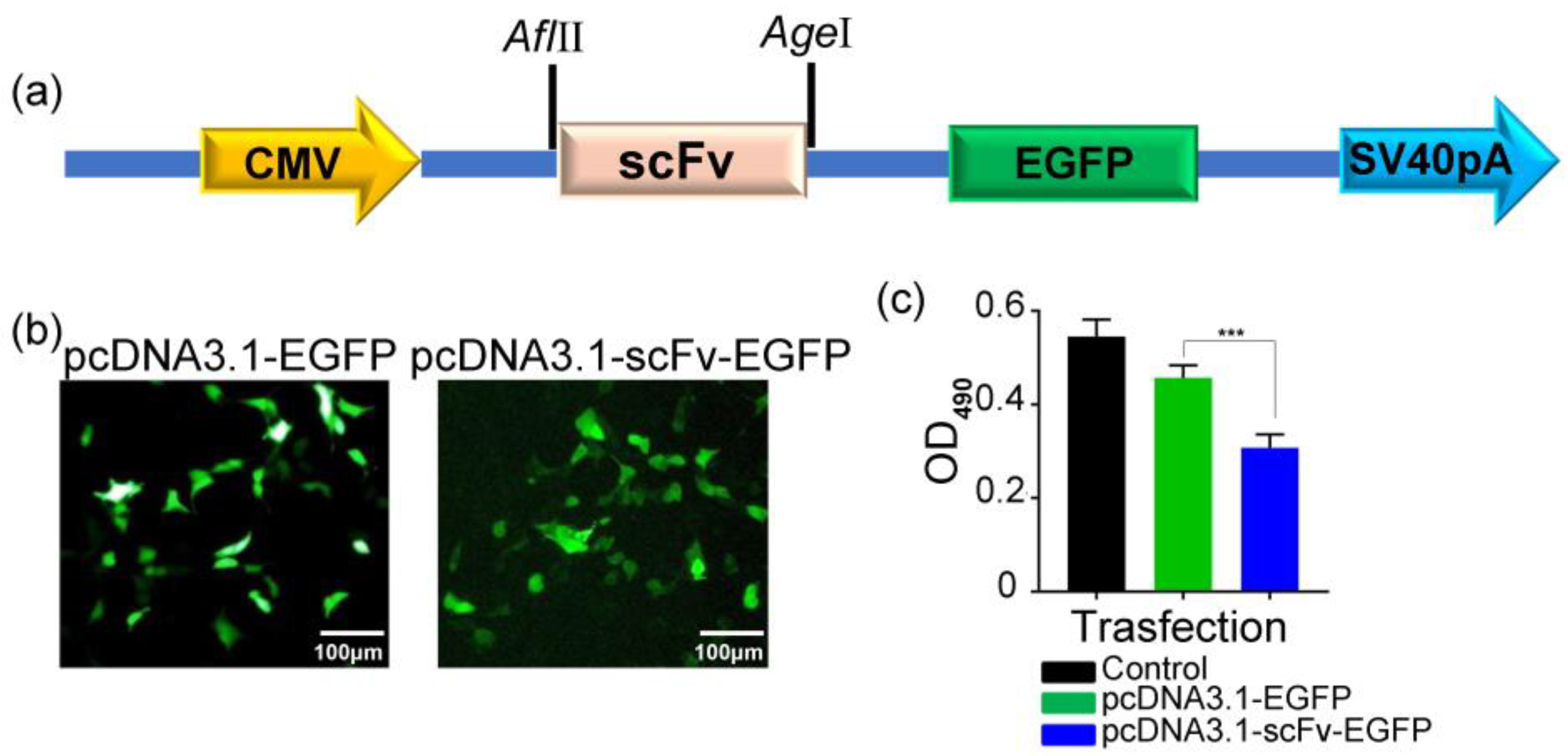
2.1. Construction, Expression, and Purification of Recombinant scFv and LP-scFv
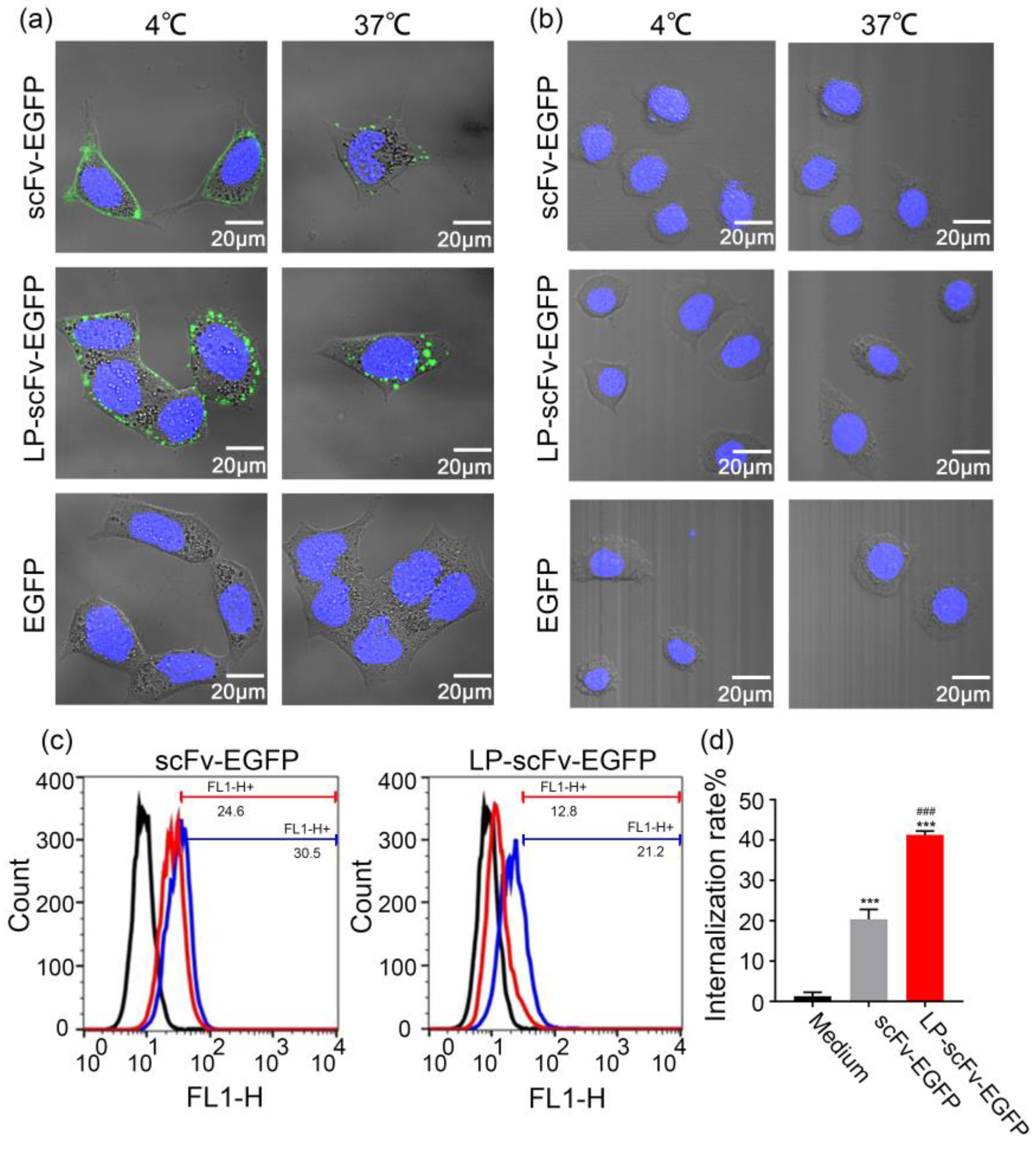
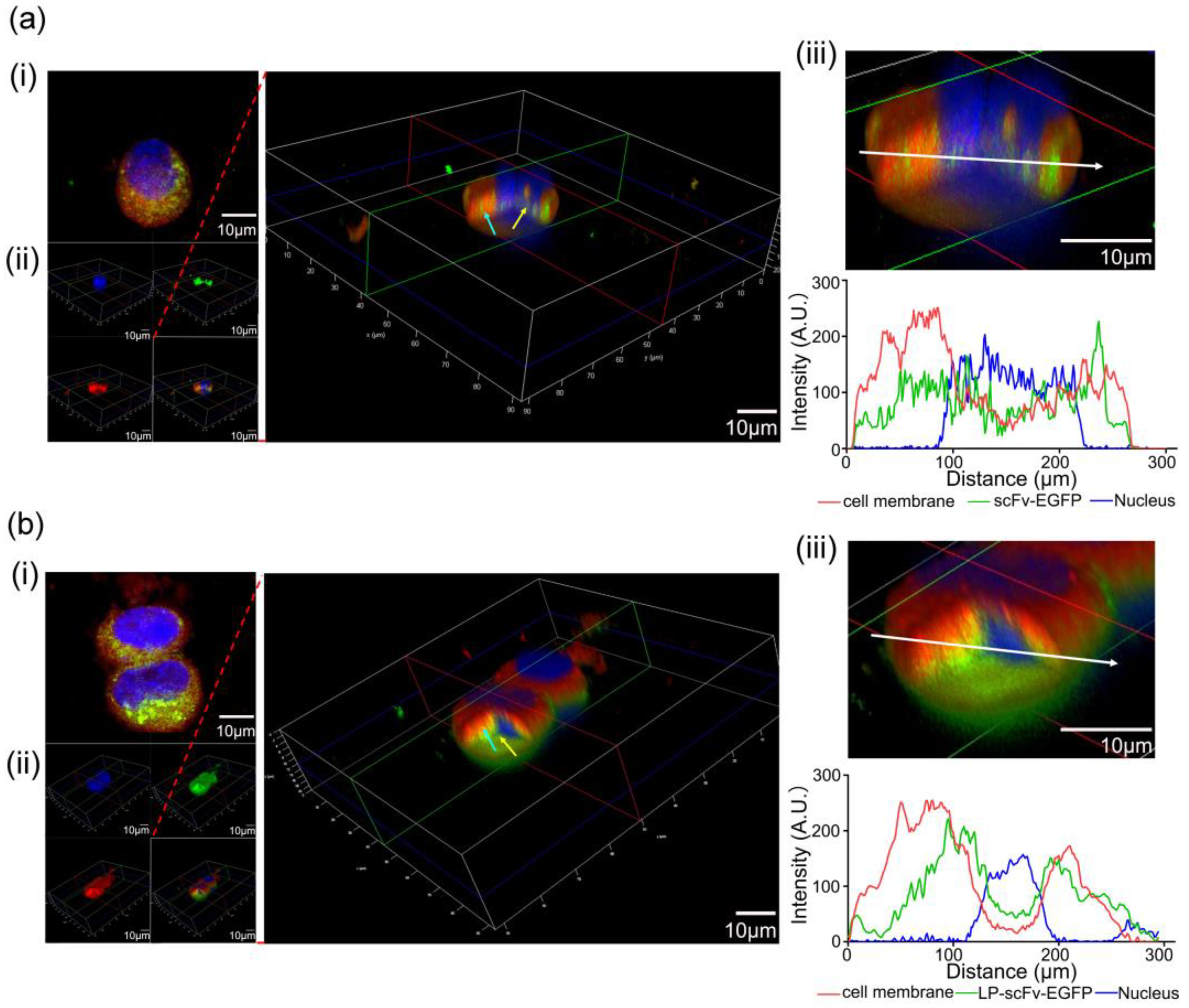
2.2. ScFv and LP-scFv Binding and Internalization
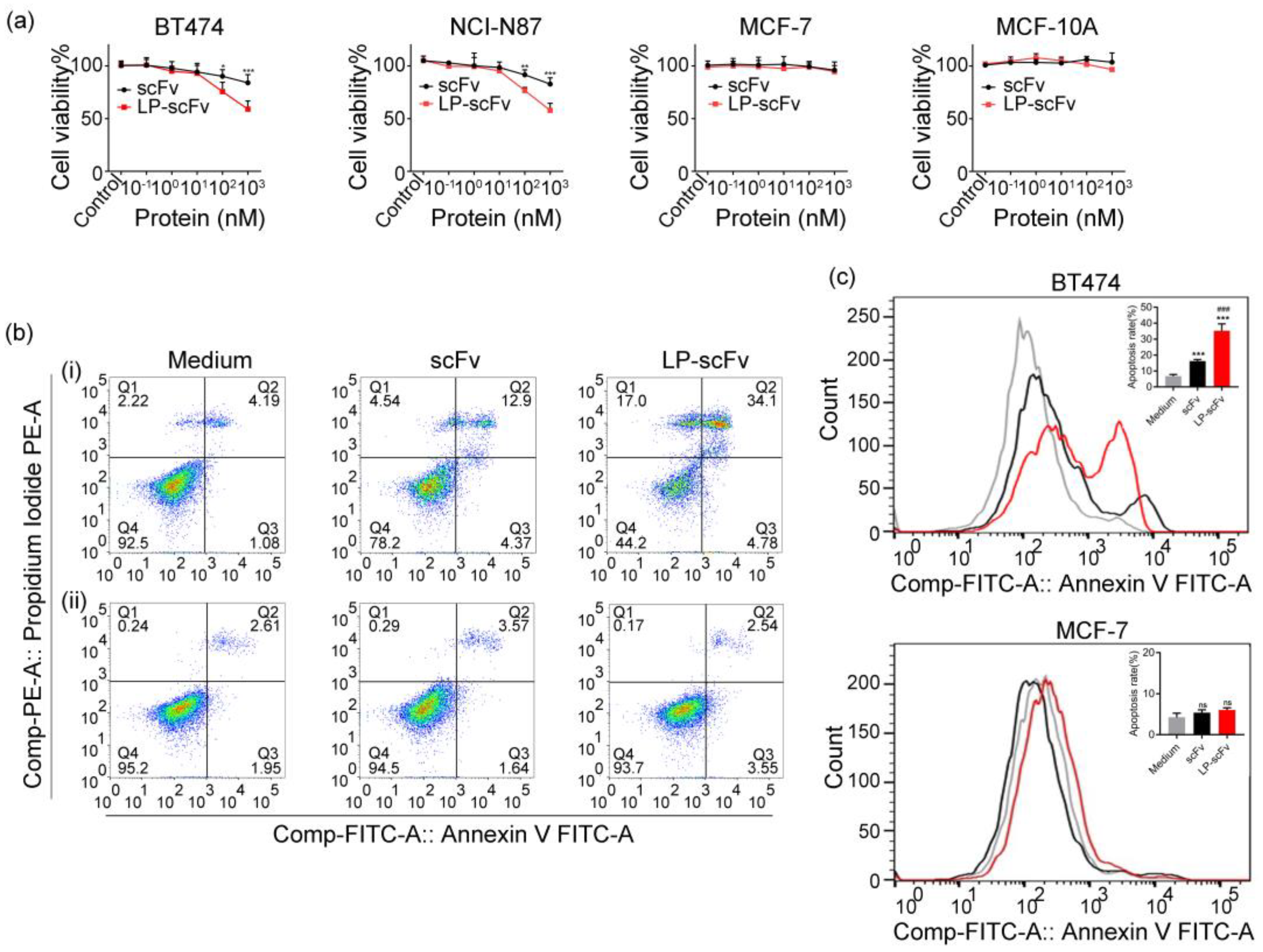
2.3. Cytotoxicity and Apoptosis Induction
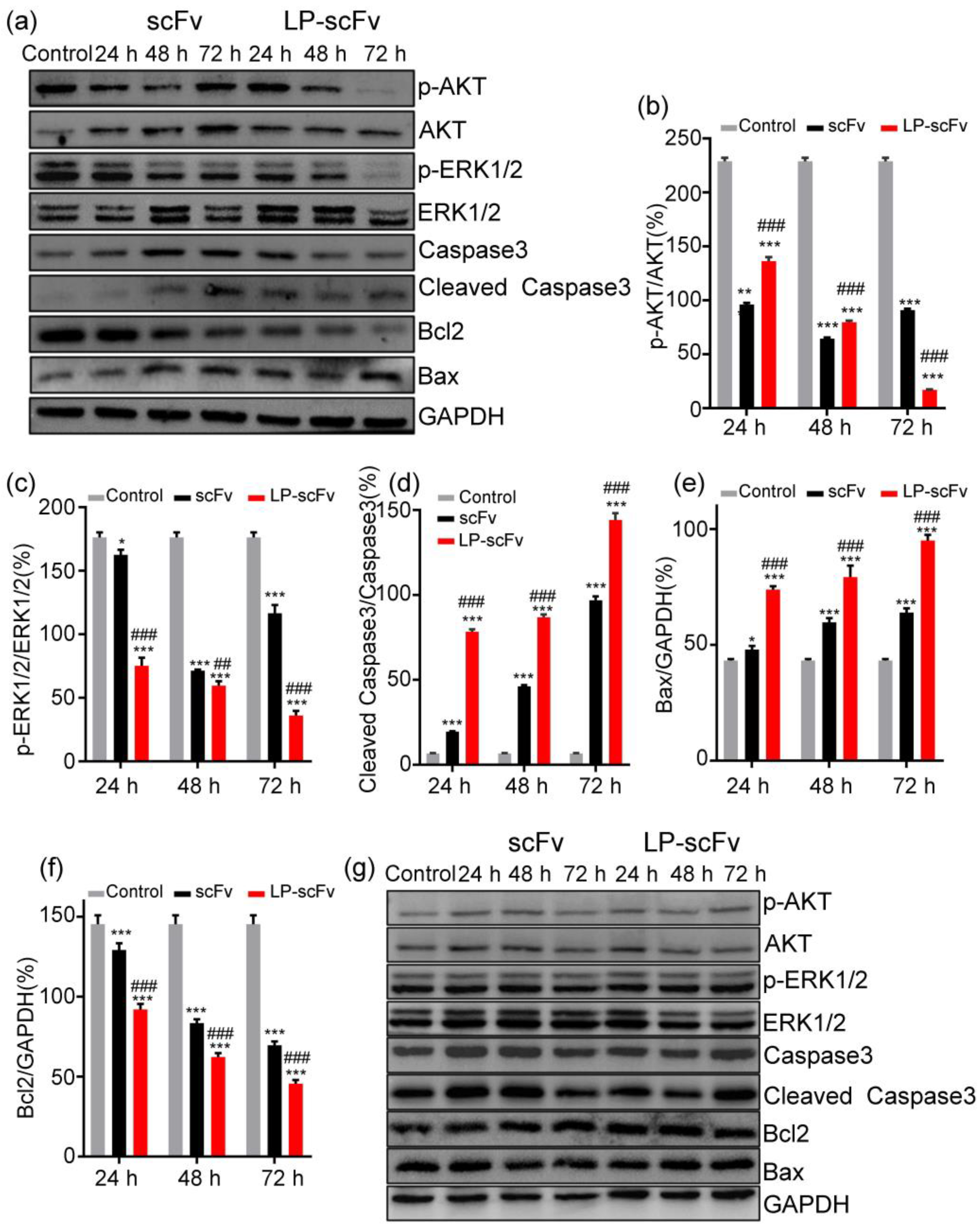
2.4. Mechanism of LP-scFv and scFv Inhibiting the Proliferation of HER2-Positive BT474 Cells
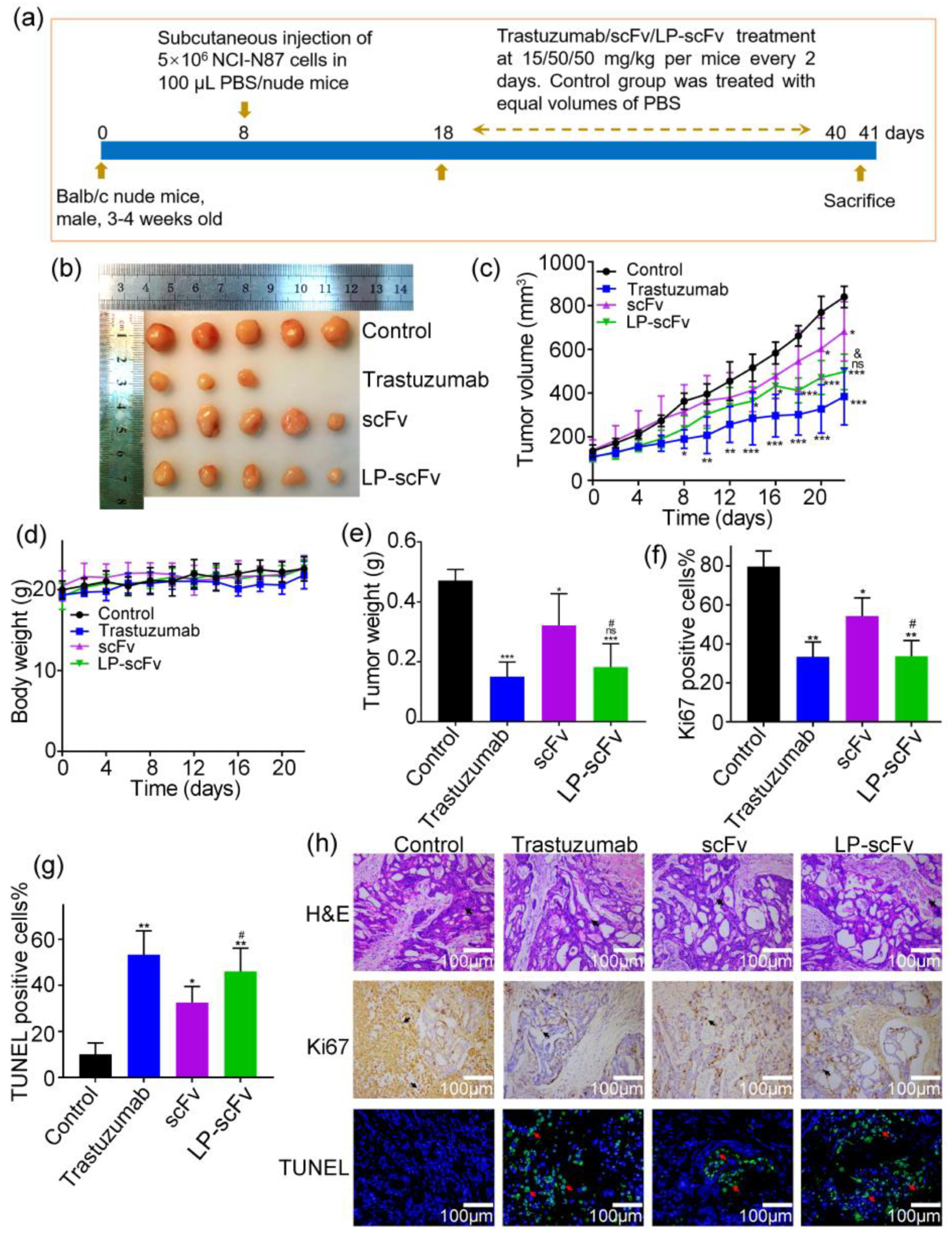
2.5. Detection of scFv and LP-scFv’s Antitumor Activity In Vivo
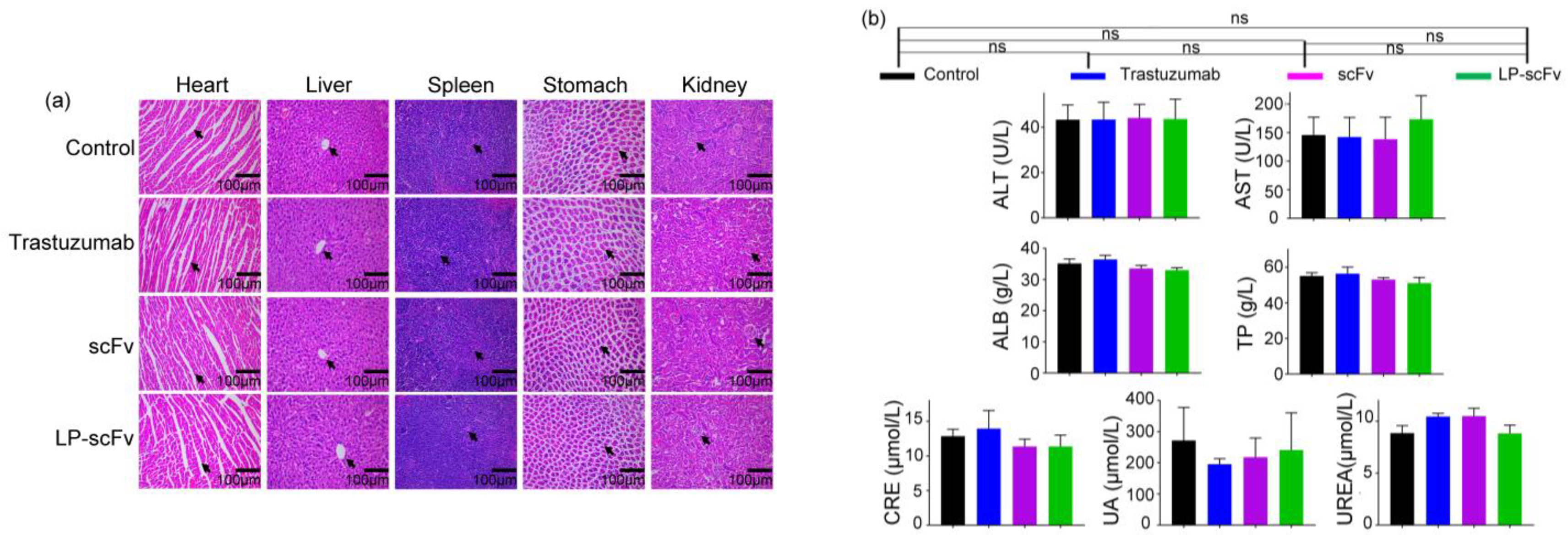
2.6. Safety Evaluation of scFv and LP-scFv In Vivo
3. Materials and Methods
3.1. Expression and Purification of LP-EGFP, scFv, and LP-scFv
3.2. Assessment of scFv and LP-scFv HER2 Affinity
3.3. Cell Culture
3.4. Binding and Internalization Analysis
3.5. MTT Assay
3.6. Measurement of Cell Apoptosis
3.7. Western Blotting
3.8. Cell Transfection
3.9. Assessment of Antitumor Effect in Xenograft Model Mice
3.10. Statistical Analysis
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Stocker, A.; Trojan, A.; Elfgen, C.; Hilbers, M.L.; Moskovszky, L.; Varga, Z. Differential prognostic value of positive HER2 status determined by immunohistochemistry or fluorescence in situ hybridization in breast cancer. Breast Cancer Res. Treat. 2020, 183, 311–319. [Google Scholar] [CrossRef]
- Swain, S.M.; Shastry, M.; Hamilton, E. Targeting HER2-positive breast cancer: Advances and future directions. Nat. Rev. Drug Discov. 2023, 22, 101–126. [Google Scholar] [CrossRef]
- DeBusk, K.; Abeysinghe, S.; Vickers, A.; Nangia, A.; Bell, J.; Ike, C.; Forero-Torres, A.; Blahna, M.T. Efficacy of tucatinib for HER2-positive metastatic breast cancer after HER2-targeted therapy: A network meta-analysis. Future Oncol. 2021, 17, 4635–4647. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.; Shastry, M.; Shiller, S.M.; Ren, R. Targeting HER2 heterogeneity in breast cancer. Cancer Treat. Rev. 2021, 100, 102286. [Google Scholar] [CrossRef] [PubMed]
- Vaisman-Mentesh, A.; Gutierrez-Gonzalez, M.; DeKosky, B.J.; Wine, Y. The molecular mechanisms that underlie the immune biology of anti-drug antibody formation following treatment with monoclonal antibodies. Front. Immunol. 2020, 11, 1951. [Google Scholar] [CrossRef]
- Deonarain, M.P.; Xue, Q. Tackling solid tumour therapy with small-format drug conjugates. Antib. Ther. 2020, 3, 237–245. [Google Scholar] [CrossRef]
- Muñoz-López, P.; Ribas-Aparicio, R.M.; Becerra-Báez, E.I.; Fraga-Pérez, K.; Flores-Martínez, L.F.; Mateos-Chávez, A.A.; Luria-Pérez, R. Single-chain fragment variable: Recent progress in cancer diagnosis and therapy. Cancers 2022, 14, 4206. [Google Scholar] [CrossRef]
- Bennion, B.J.; Be, N.A.; McNerney, M.W.; Lao, V.; Carlson, E.M.; Valdez, C.A.; Malfatti, M.A.; Enright, H.A.; Nguyen, T.H.; Lightstone, F.C.; et al. Predicting a drug’s membrane permeability: A computational model validated with in vitro permeability assay data. J. Phys. Chem. B 2017, 121, 5228–5237. [Google Scholar] [CrossRef]
- Gaston, J.; Maestrali, N.; Lalle, G.; Gagnaire, M.; Masiero, A.; Dumas, B.; Dabdoubi, T.; Radošević, K.; Berne, P.F. Intracellular delivery of therapeutic antibodies into specific cells using antibody-peptide fusions. Sci. Rep. 2019, 9, 18688. [Google Scholar] [CrossRef]
- Habault, J.; Poyet, J.L. Recent advances in cell penetrating peptide-based anticancer therapies. Molecules 2019, 24, 927. [Google Scholar] [CrossRef] [PubMed]
- Kardani, K.; Bolhassani, A. Cppsite 2.0: An available aatabase of experimentally validated cell-penetrating peptides predicting their secondary and tertiary structures. J. Mol. Biol. 2021, 433, 166703. [Google Scholar] [CrossRef]
- Sun, Z.; Huang, J.; Fishelson, Z.; Wang, C.; Zhang, S. Cell-Penetrating Peptide-Based Delivery of Macromolecular Drugs: Development, Strategies, and Progress. Biomedicines 2023, 11, 1971. [Google Scholar] [CrossRef] [PubMed]
- Polli, J.R.; Balthasar, J.P. Cell penetrating peptides conjugated to anti-carcinoembryonic antigen “catch-and-release” monoclonal antibodies alter plasma and tissue pharmacokinetics in colorectal cancer xenograft mice. Bioconjug. Chem. 2022, 33, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Tietz, O.; Cortezon-Tamarit, F.; Chalk, R.; Able, S.; Vallis, K.A. Tricyclic cell-penetrating peptides for efficient delivery of functional antibodies into cancer cells. Nat. Chem. 2022, 14, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Kurrikoff, K.; Langel, Ü.; Almeida, A.J.; Vale, N. A second life for MAP, a model amphipathic peptide. Int. J. Mol. Sci. 2022, 23, 8322. [Google Scholar] [CrossRef]
- Kenien, R.; Shen, W.C.; Zaro, J.L. Vesicle-to-cytosol transport of disulfide-linked cargo mediated by an amphipathic cell-penetrating peptide. J. Drug. Target 2012, 20, 793–800. [Google Scholar] [CrossRef]
- Del Genio, V.; Falanga, A.; Allard-Vannier, E.; Hervé-Aubert, K.; Leone, M.; Bellavita, R.; Uzbekov, R.; Chourpa, I.; Galdiero, S. Design and validation of nanofibers made of self-assembled peptides to become multifunctional stimuli-sensitive nanovectors of anticancer drug doxorubicin. Pharmaceutics 2022, 14, 1544. [Google Scholar] [CrossRef]
- Govindarajan, S.; Sivakumar, J.; Garimidi, P.; Rangaraj, N.; Kumar, J.M.; Rao, N.M.; Gopal, V. Targeting human epidermal growth factor receptor 2 by a cell-penetrating peptide–affibody bioconjugate. Biomaterials 2012, 33, 2570–2582. [Google Scholar] [CrossRef]
- Sauter, M.; Strieker, M.; Kleist, C.; Wischnjow, A.; Daniel, V.; Altmann, A.; Haberkorn, U.; Mier, W. Improving antibody-based therapies by chemical engineering of antibodies with multimeric cell-penetrating peptides for elevated intracellular delivery. J. Control. Release 2020, 322, 200–208. [Google Scholar] [CrossRef]
- Bae, J.; Song, Y. Engineering a cell-penetrating hyperstable antibody scFv(Ras)—An extraordinary approach to cancer therapeutics. Synth. Syst. Biotechnol. 2021, 6, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, V.; Beaudoin, S.; Jean, S.; Leyton, J.V. A novel proteomic method reveals NLS tagging of T-DM1 contravenes classical nuclear transport in a model of HER2-positive breast cancer. Mol. Ther. Methods Clin. Dev. 2020, 19, 99–119. [Google Scholar] [CrossRef] [PubMed]
- Roise, D.; Horvath, S.J.; Tomich, J.M.; Richards, J.H.; Schatz, G. A chemically synthesized pre-sequence of an imported mitochondrial protein can form an amphiphilic helix and perturb natural and artificial phospholipid bilayers. EMBO J. 1986, 5, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing mitochondrial proteins: Machineries and mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef]
- Wiedemann, N.; Pfanner, N. Mitochondrial machineries for protein import and assembly. Annu. Rev. Biochem. 2017, 86, 685–714. [Google Scholar] [CrossRef]
- Busch, J.D.; Fielden, L.F.; Pfanner, N.; Wiedemann, N. Mitochondrial protein transport: Versatility of translocases and mechanisms. Mol. Cell 2023, 83, 890–910. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.E.; Zahid, M. Cell penetrating peptides, novel vectors for gene therapy. Pharmaceutics 2020, 12, 225. [Google Scholar] [CrossRef]
- Zhou, W.; Lin, S.; Chen, R.; Liu, J.; Li, Y. Characterization of antibody-C1q interactions by biolayer interferometry. Anal. Biochem. 2018, 549, 143–148. [Google Scholar] [CrossRef]
- Láng, J.A.; Balogh, Z.C.; Nyitrai, M.F.; Juhász, C.; Gilicze, A.K.B.; Iliás, A.; Zólyomi, Z.; Bodor, C.; Rábai, E. In vitro functional characterization of biosimilar therapeutic antibodies. Drug Discov. Today Technol. 2020, 37, 41–50. [Google Scholar] [CrossRef]
- Li, Z.H.; Zhang, Q.; Wang, H.B.; Zhang, Y.N.; Ding, D.; Pan, L.Q.; Miao, D.; Xu, S.; Zhang, C.; Luo, P.H.; et al. Preclinical studies of targeted therapies for CD20-positive B lymphoid malignancies by Ofatumumab conjugated with auristatin. Investig. New Drugs 2014, 32, 75–86. [Google Scholar] [CrossRef]
- Austin, C.D.; De Maziere, A.M.; Pisacane, P.I.; van Dijk, S.M.; Eigenbrot, C.; Sliwkowski, M.X.; Klumperman, J.; Scheller, R.H. Endocytosis and sorting of ErbB2 and the site of action of cancer therapeutics trastuzumab and geldanamycin. Mol. Biol. Cell 2004, 15, 5268–5282. [Google Scholar] [CrossRef]
- Wang, Y.N.; Lee, H.H.; Lee, H.J.; Du, Y.; Yamaguchi, H.; Hung, M.C. Membrane-bound trafficking regulates nuclear transport of integral epidermal growth factor receptor (EGFR) and ErbB-2. J. Biol. Chem. 2012, 287, 16869–16879. [Google Scholar] [CrossRef] [PubMed]
- Elizalde, P.V.; Russo, R.I.C.; Chervo, M.F.; Schillaci, R. ErbB-2 nuclear function in breast cancer growth, metastasis and resistance to therapy. Endocr. Relat. Cancer 2016, 23, T243–T257. [Google Scholar] [CrossRef] [PubMed]
- Agus, D.B.; Akita, R.W.; Fox, W.D.; Lewis, G.D.; Higgins, B.; Pisacane, P.I.; Lofgren, J.A.; Tindell, C.; Evans, D.P.; Maiese, K.; et al. Targeting ligand-activated ErbB2 signaling inhibits breast and prostate tumor growth. Cancer Cell 2002, 2, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Maadi, H.; Soheilifar, M.H.; Choi, W.S.; Moshtaghian, A.; Wang, Z. Trastuzumab mechanism of action; 20 years of research to unravel a dilemma. Cancers 2021, 13, 3540. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Bang, Y.J. HER2-targeted therapies—A role beyond breast cancer. Nat. Rev. Clin. Oncol. 2020, 17, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.K.; Lee, S.Y.; Lee, Y.H.; Hwang, I.S.; Persson, H.; Rockberg, J.; Borrebaeck, C.; Park, D.; Kim, K.T.; Uhlen, M.; et al. Combination of novel HER2-targeting antibody 1E11 with trastuzumab shows synergistic antitumor activity in HER2-positive gastric cancer. Mol. Oncol. 2015, 9, 398–408. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, M.Y.; Hong, S. Synergistic effects of metformin and trastuzumab on HER2 positive gastroesophageal adenocarcinoma cells in vitro and in vivo. Cancers 2023, 15, 4768. [Google Scholar] [CrossRef]
- Sobecki, M.; Mrouj, K.; Camasses, A.; Parisis, N.; Nicolas, E.; Llères, D.; Gerbe, F.; Prieto, S.; Krasinska, L.; David, A.; et al. The cell proliferation antigen Ki-67 organises heterochromatin. eLife 2016, 5, e13722. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Galkowski, D.; Zhao, H. Analysis of apoptosis by cytometry using TUNEL assay. Methods 2008, 44, 250–254. [Google Scholar] [CrossRef]
- Chung, W.P.; Huang, W.L.; Lee, C.H.; Hsu, H.P.; Huang, W.L.; Liu, Y.Y.; Su, W.C. PI3K inhibitors in trastuzumab-resistant HER2-positive breast cancer cells with PI3K pathway alterations. Am. J. Cancer Res. 2022, 12, 3067–3082. [Google Scholar] [PubMed]
- Fujimoto-Ouchi, K.; Sekiguchi, F.; Yasuno, H.; Moriya, Y.; Mori, K.; Tanaka, Y. Antitumor activity of trastuzumab in combination with chemotherapy in human gastric cancer xenograft models. Cancer Chemother. Pharmacol. 2007, 59, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Yamashita-Kashima, Y.; Iijima, S.; Yorozu, K.; Furugaki, K.; Kurasawa, M.; Ohta, M.; Fujimoto-Ouchi, K. Pertuzumab in combination with Trastuzumab shows significantly enhanced antitumor activity in HER2-positive human gastric cancer xenograft models. Clin. Cancer Res. 2011, 17, 5060–5070. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Lee, F.; Gao, S.; Tan, M.-H.; Kurisawa, M. Hyaluronidase-incorporated hyaluronic acid–tyramine hydrogels for the sustained release of trastuzumab. J. Control. Release 2015, 216, 47–55. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Antibody | kon (M−1S−1) | koff (S−1) | KD (nM) | χ2 | R2 |
|---|---|---|---|---|---|---|
| HER2 | scFv | (1.05 ± 0.15) × 105 | (2.28 ± 0.31) × 10−3 | 21.6 ± 2.1 | 0.04 | 0.97 |
| HER2 | LP-scFv | (4.49 ± 0.49) × 104 | (8.63 ± 2.24) × 10−4 | 19.2 ± 3.4 | 0.05 | 0.97 |
| Group Name | n | Injection Interval (Day) | Experiment Time (Weeks) | Treatment |
|---|---|---|---|---|
| Control | 5 | 2 | 3 | PBS (control, subcutaneous administration) |
| Trastuzumab | 3 | 2 | 3 | PBS + Trastuzumab (15 mg/kg, subcutaneous administration) |
| scFv | 5 | 2 | 3 | PBS + scFv (50 mg/kg, subcutaneous administration) |
| LP-scFv | 5 | 2 | 3 | PBS + LP-scFv (50 mg/kg, subcutaneous administration) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhou, Y.; Su, Z.; Li, X.; Zhang, L.; Li, S. A Cell-Penetrating Peptide Improves Anti-HER2 Single-Chain Variable Fragment Internalization and Antitumor Activity against HER2-Positive Breast Cancer In Vitro and In Vivo. Molecules 2024, 29, 1247. https://doi.org/10.3390/molecules29061247
Li J, Zhou Y, Su Z, Li X, Zhang L, Li S. A Cell-Penetrating Peptide Improves Anti-HER2 Single-Chain Variable Fragment Internalization and Antitumor Activity against HER2-Positive Breast Cancer In Vitro and In Vivo. Molecules. 2024; 29(6):1247. https://doi.org/10.3390/molecules29061247
Chicago/Turabian StyleLi, Junmin, Yanting Zhou, Zhuowei Su, Xue Li, Lei Zhang, and Shan Li. 2024. "A Cell-Penetrating Peptide Improves Anti-HER2 Single-Chain Variable Fragment Internalization and Antitumor Activity against HER2-Positive Breast Cancer In Vitro and In Vivo" Molecules 29, no. 6: 1247. https://doi.org/10.3390/molecules29061247
APA StyleLi, J., Zhou, Y., Su, Z., Li, X., Zhang, L., & Li, S. (2024). A Cell-Penetrating Peptide Improves Anti-HER2 Single-Chain Variable Fragment Internalization and Antitumor Activity against HER2-Positive Breast Cancer In Vitro and In Vivo. Molecules, 29(6), 1247. https://doi.org/10.3390/molecules29061247