
Untargeted Metabolomics Based on UPLC-Q-Exactive-Orbitrap-MS/MS Revealed the Differences and Correlations between Different Parts of the Root of Paeonia lactiflora Pall
 , , , , , , and
, , , , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
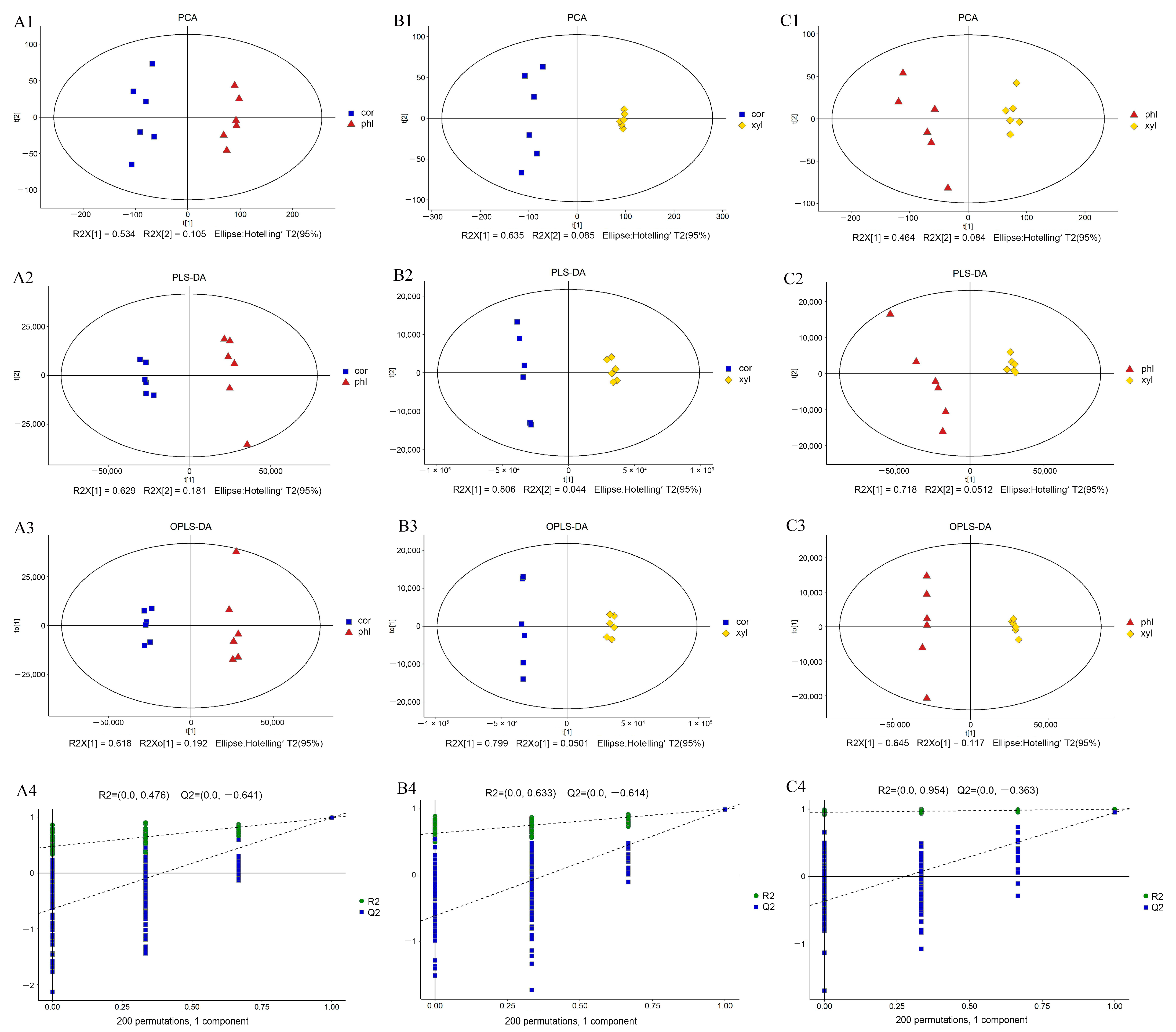
2.1. Multivariate Statistical Analysis
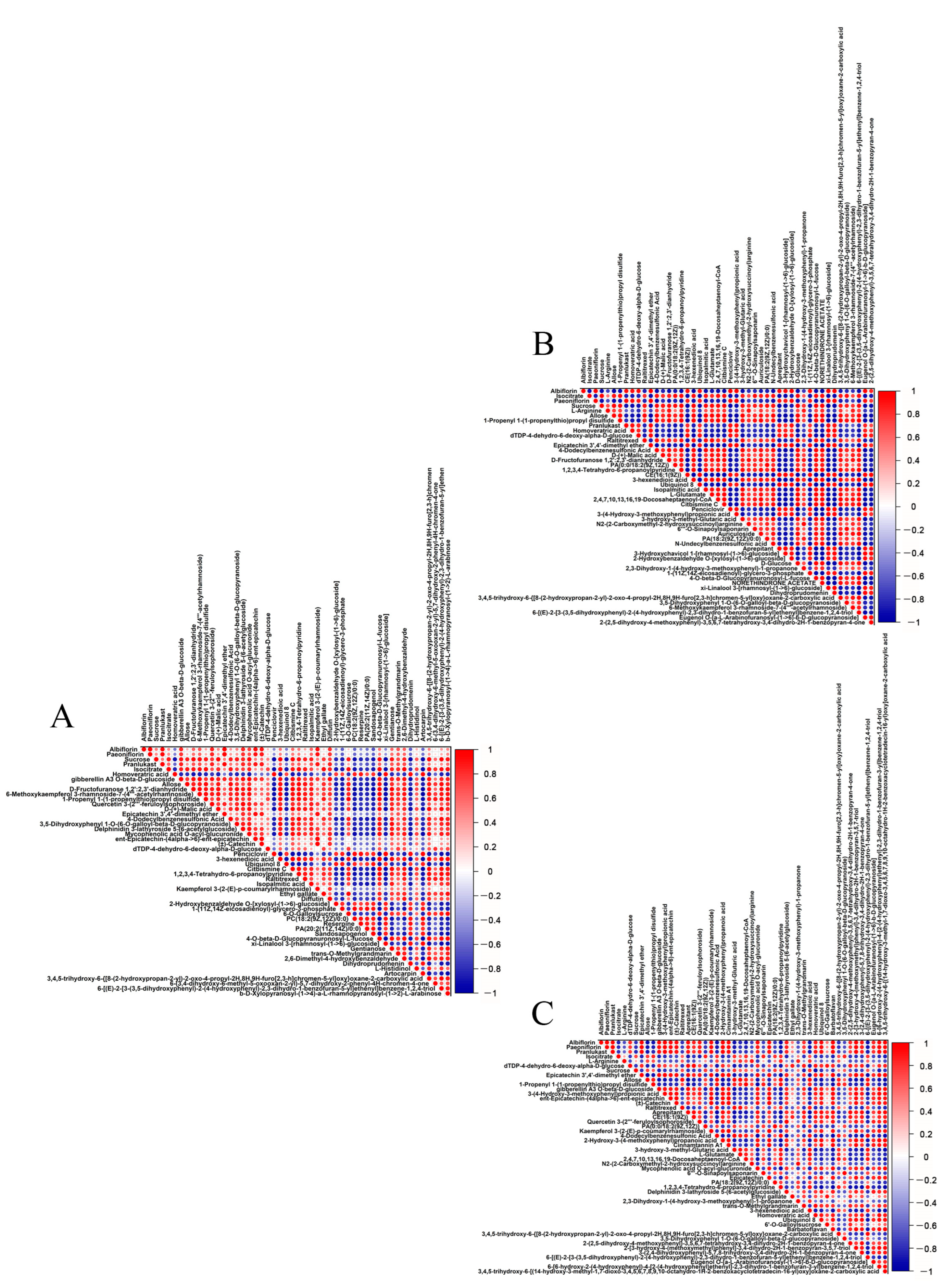
2.2. Analysis of Differential Metabolites
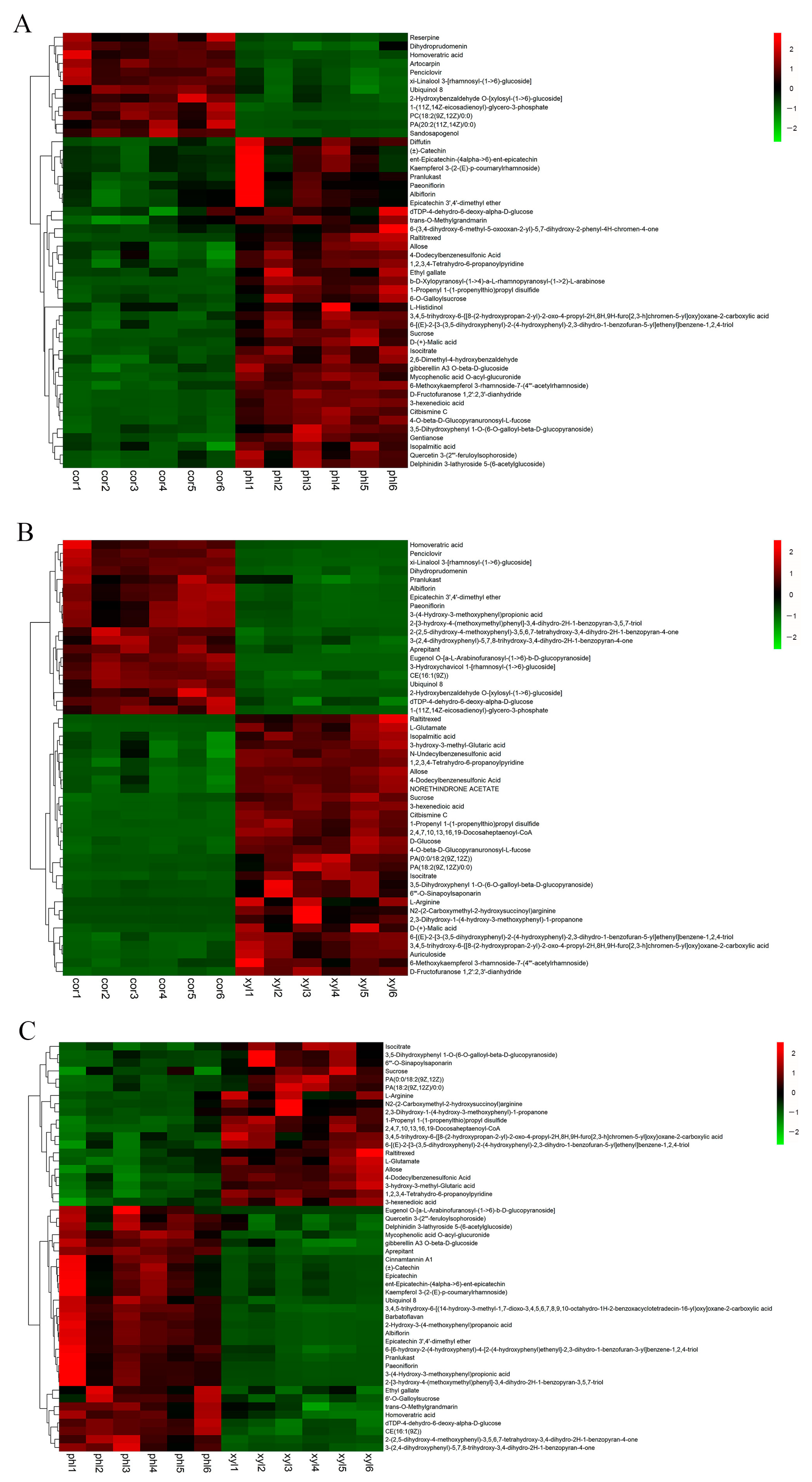
2.3. Analysis of the Top 50 Differential Metabolites
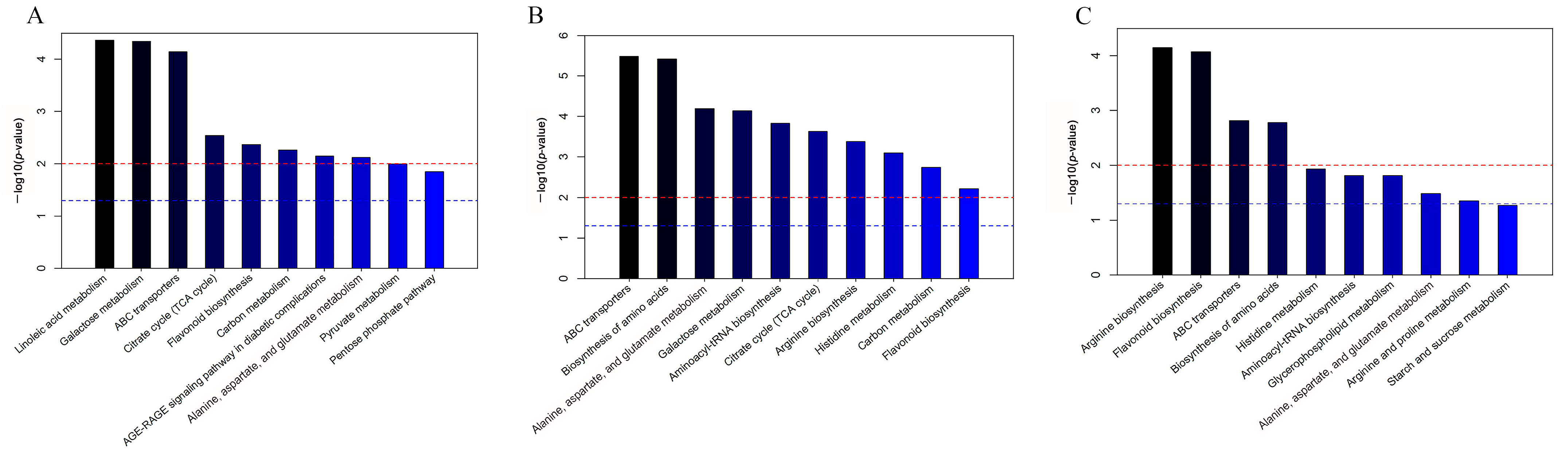
2.4. Enrichment Analysis of Metabolic Pathways
3. Materials and Methods
3.1. Reagents and Materials
3.2. Sample Preparation
3.3. UPLC-MS/MS Analysis
3.4. Data Preprocessing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Tan, Y.Q.; Chen, H.W.; Li, J.; Wu, Q.J. Efficacy, Chemical Constituents, and Pharmacological Actions of Radix Paeoniae Rubra and Radix Paeoniae Alba. Front. Pharmacol. 2020, 11, 1054. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shen, J.; Wang, Z.; Liu, S.; Liu, Q.; Li, Y.; He, C.; Xiao, P. Genus Paeonia: A comprehensive review on traditional uses, phytochemistry, pharmacological activities, clinical application, and toxicology. J. Ethnopharmacol. 2021, 269, 113708. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2020; pp. 108–165. [Google Scholar]
- Zhu, S.; Shirakawa, A.; Shi, Y.; Yu, X.; Tamura, T.; Shibahara, N.; Yoshimatsu, K.; Komatsu, K. Impact of different post-harvest processing methods on the chemical compositions of peony root. J. Nat. Med. 2018, 72, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.L.; Yang, P.W.; Qian, D.W.; Bian, X.K.; Wang, Y.F.; Zhu, Z.H.; Guo, S.; Wang, T.J.; Yan, H.; Duan, J.A. Effects and evaluation of different processing and drying methods on components in Paeoniae Radix Alba. Zhongguo Zhong Yao Za Zhi 2021, 46, 5839–5847. [Google Scholar] [PubMed]
- Wang, Q.; Liang, Z.; Peng, Y.; Hou, J.L.; Wei, S.L.; Zhao, Z.Z.; Wang, W.Q. Whole transverse section and specific-tissue analysis of secondary metabolites in seven different grades of root of Paeonia lactiflora using laser microdissection and liquid chromatography-quadrupole/time of flight-mass spectrometry. J. Pharm. Biomed. Anal. 2015, 103, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Liu, W.; Liu, X.; Qin, C.; He, L.; Wang, P.; Kong, L.; Chen, X.; Liu, Z.; Ma, W. DUS evaluation of nine intersubgeneric hybrids of Paeonia lactiflora and fingerprint analysis of the chemical components in the roots. Front. Chem. 2023, 11, 1158727. [Google Scholar] [CrossRef]
- Ueno, Y.; Suzuki, R.; Kitamura, M. (1)H-NMR-Based Metabolomics for the Classification of the Roots of Paeonia lactiflora, a Constituent of Kampo Medicines. Chem. Pharm. Bull. 2022, 70, 859–862. [Google Scholar] [CrossRef]
- Chen, W.J.; Zheng, Y.N.; Zhao, L.; Song, S.H.; Long, F.; Pei, Z.Q.; Tang, C.; Xu, Z.G.; Lv, G.H. Distribution of bioactive compounds in different tissues of Paeonia lactiflora roots by DESI-MSI and UPLC. Zhongguo Zhong Yao Za Zhi 2022, 47, 4333–4340. [Google Scholar]
- Shen, M.; Zhang, Q.; Qin, L.; Yan, B. Single Standard Substance for the Simultaneous Determination of Eleven Components in the Extract of Paeoniae Radix Alba (Root of Paeonia lactiflora Pall.). J. Anal. Methods Chem. 2021, 2021, 8860776. [Google Scholar] [CrossRef]
- Trovato, E.; Vento, F.; Creti, D.; Dugo, P.; Mondello, L. Elucidation of Analytical-Compositional Fingerprinting of Three Different Species of Chili Pepper by Using Headspace Solid-Phase Microextraction Coupled with Gas Chromatography-Mass Spectrometry Analysis, and Sensory Profile Evaluation. Molecules 2022, 27, 2355. [Google Scholar] [CrossRef]
- Kazemzadeh, M.; Hisey, C.L.; Zargar-Shoshtari, K.; Xu, W.; Broderick, N.G.R. Deep convolutional neural networks as a unified solution for Raman spectroscopy-based classification in biomedical applications. Opt. Commun. 2022, 510, 127977. [Google Scholar] [CrossRef]
- Di Minno, A.; Gelzo, M.; Stornaiuolo, M.; Ruoppolo, M.; Castaldo, G. The evolving landscape of untargeted metabolomics. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 1645–1652. [Google Scholar] [CrossRef]
- Zeki, Ö.C.; Eylem, C.C.; Reçber, T.; Kır, S.; Nemutlu, E. Integration of GC-MS and LC-MS for untargeted metabolomics profiling. J. Pharm. Biomed. Anal. 2020, 190, 113509. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Hong, Y.; Chen, Z. Identification and quantification of the bioactive components in Osmanthus fragrans roots by HPLC-MS/MS. J. Pharm. Anal. 2021, 11, 299–307. [Google Scholar] [CrossRef] [PubMed]
- He, D.Y.; Dai, S.M. Anti-inflammatory and immunomodulatory effects of Paeonia lactiflora pall., a traditional chinese herbal medicine. Front. Pharmacol. 2011, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Li, L.X.; Wu, W.Z.; Pan, T.J.; Yang, Z.S.; Yang, Y.K. Anti-Tumor Effects of Paeoniflorin on Epithelial-To-Mesenchymal Transition in Human Colorectal Cancer Cells. Med. Sci. Monit. 2018, 24, 6405–6413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wei, W. Anti-inflammatory and immunoregulatory effects of paeoniflorin and total glucosides of paeony. Pharmacol. Ther. 2020, 207, 107452. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, Y.; Zuo, W.T.; Gao, Y.R.; Li, R.Z.; Yu, C.X.; Liu, Z.Y.; Zheng, Y.; Shen, Y.Y.; Duan, L.S. Integration of Metabolome and Transcriptome Studies Reveals Flavonoids, Abscisic Acid, and Nitric Oxide Comodulating the Freezing Tolerance in Liriope spicata. Front. Plant Sci. 2021, 12, 764625. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Suo, J.; Liu, S.; Xu, J.; Yang, T.; Xiang, N.; Wu, Y.; Lu, B.; Qin, R.; Liu, H.; et al. Combined transcriptomic and metabolomic analysis reveals the potential mechanism of seed germination and young seedling growth in Tamarix hispida. BMC Genom. 2022, 23, 109. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Reyes, I.; Cardona, L.R.; Kong, H.; Vasan, K.; McElroy, G.S.; Werner, M.; Kihshen, H.; Reczek, C.R.; Weinberg, S.E.; Gao, P.; et al. Mitochondrial ubiquinol oxidation is necessary for tumour growth. Nature 2020, 585, 288–292. [Google Scholar] [CrossRef]
- Nava Lauson, C.B.; Tiberti, S.; Corsetto, P.A.; Conte, F.; Tyagi, P.; Machwirth, M.; Ebert, S.; Loffreda, A.; Scheller, L.; Sheta, D.; et al. Linoleic acid potentiates CD8(+) T cell metabolic fitness and antitumor immunity. Cell Metab. 2023, 35, 633–650.e9. [Google Scholar] [CrossRef]
- Liu, Y.J.; Li, H.; Tian, Y.; Han, J.; Wang, X.Y.; Li, X.Y.; Tian, C.; Zhang, P.H.; Hao, Y.; Gao, F.; et al. PCTR1 ameliorates lipopolysaccharide-induced acute inflammation and multiple organ damage via regulation of linoleic acid metabolism by promoting FADS1/FASDS2/ELOV2 expression and reducing PLA2 expression. Lab. Investig. 2020, 100, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, N.; Imi, Y.; Amano, R.; Shinohara, M.; Okada, K.; Hosokawa, Y.; Imamori, M.; Tomimoto, C.; Kunisawa, J.; Kishino, S.; et al. A gut microbial metabolite of linoleic acid ameliorates liver fibrosis by inhibiting TGF-β signaling in hepatic stellate cells. Sci. Rep. 2023, 13, 18983. [Google Scholar] [CrossRef] [PubMed]
- Santoro, N.; Caprio, S.; Feldstein, A.E. Oxidized metabolites of linoleic acid as biomarkers of liver injury in nonalcoholic steatohepatitis. Clin. Lipidol. 2013, 8, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gan, J.; Li, J.; Wang, Y.; Zhang, J.; Song, L.; Yang, Z.; Guo, M.; Jiang, X. Shengmai Yin formula exerts cardioprotective effects on rats with chronic heart failure via regulating Linoleic Acid metabolism. Prostaglandins Other Lipid Mediat. 2022, 158, 106608. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: Time for a closer look. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129898. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.S.; Li, M.L.; Chen, J.S.; Zhou, L.; Zhou, W. Application of Mono- and Disaccharides in Drug Targeting and Efficacy. ChemMedChem 2018, 13, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Liu, S.; Jiang, X.; Zhang, X.; Wei, F.; Hu, J. The Qingchangligan Formula Alleviates Acute Liver Failure by Regulating Galactose Metabolism and Gut Microbiota. Front. Cell. Infect. Microbiol. 2021, 11, 771483. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Bai, J.; Du, Y.; Tan, P.; Zheng, T.; Chen, Y.; Cheng, Y.; Cai, T.; Huang, M.; Fu, W.; et al. Thiamine pretreatment improves endotoxemia-related liver injury and cholestatic complications by regulating galactose metabolism and inhibiting macrophage activation. Int. Immunopharmacol. 2022, 108, 108892. [Google Scholar] [CrossRef]
- Böger, R.H. The pharmacodynamics of L-arginine. Altern. Ther. Health Med. 2014, 20, 48–54. [Google Scholar] [CrossRef]
- Szefel, J.; Danielak, A.; Kruszewski, W.J. Metabolic pathways of L-arginine and therapeutic consequences in tumors. Adv. Med. Sci. 2019, 64, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Sanders, P.W. L-arginine and arginine analogs in progressive renal failure. Blood Purif. 1995, 13, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lv, S.; Zhao, L.; Gao, T.; Yu, C.; Hu, J.; Ma, F. Advances in the study of the function and mechanism of the action of flavonoids in plants under environmental stresses. Planta 2023, 257, 108. [Google Scholar] [CrossRef] [PubMed]
- Kongpichitchoke, T.; Hsu, J.L.; Huang, T.C. Number of Hydroxyl Groups on the B-Ring of Flavonoids Affects Their Antioxidant Activity and Interaction with Phorbol Ester Binding Site of PKCδ C1B Domain: In Vitro and in Silico Studies. J. Agric. Food Chem. 2015, 63, 4580–4586. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, J.P.; Oresnik, I.J. The Rhizobium-Legume Symbiosis: Co-opting Successful Stress Management. Front. Plant Sci. 2021, 12, 796045. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Pei, Y.; Huang, W.; Ding, J.; Siemann, E. Increasing flavonoid concentrations in root exudates enhance associations between arbuscular mycorrhizal fungi and an invasive plant. ISME J. 2021, 15, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Nogia, P.; Pati, P.K. Plant Secondary Metabolite Transporters: Diversity, Functionality, and Their Modulation. Front. Plant Sci. 2021, 12, 758202. [Google Scholar] [CrossRef]
- Tullus, A.; Rusalepp, L.; Lutter, R.; Rosenvald, K.; Kaasik, A.; Rytter, L.; Kontunen-Soppela, S.; Oksanen, E. Climate and Competitive Status Modulate the Variation in Secondary Metabolites More in Leaves Than in Fine Roots of Betula pendula. Front. Plant Sci. 2021, 12, 746165. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, Z.Y.; Zhang, X.H.; Shao, J.H.; Zhao, C.C. Recent Advance on Chemistry and Bioactivities of Secondary Metabolites from Viburnum Plants: An Update. Chem. Biodivers. 2021, 18, e2100404. [Google Scholar] [CrossRef]
- Shi, S.Y.; Yu, N.J.; Peng, D.Y.; Zhang, Y.L.; Zhu, Q.; GE, D.Z.; MA, L.; Schroder, S.; Han, R.C. Study on Different Parts from Root of Paeonia lactiflora Pall. by HPLC Characteristic Chromatogram. J. Yunnan Univ. Tradit. Chin. Med. 2020, 43, 12–18. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, J.; Du, Q.; Shi, S.; Ma, M.; Zhang, W.; Ge, D.; Xing, L.; Yu, N. Untargeted Metabolomics Based on UPLC-Q-Exactive-Orbitrap-MS/MS Revealed the Differences and Correlations between Different Parts of the Root of Paeonia lactiflora Pall. Molecules 2024, 29, 992. https://doi.org/10.3390/molecules29050992
Lv J, Du Q, Shi S, Ma M, Zhang W, Ge D, Xing L, Yu N. Untargeted Metabolomics Based on UPLC-Q-Exactive-Orbitrap-MS/MS Revealed the Differences and Correlations between Different Parts of the Root of Paeonia lactiflora Pall. Molecules. 2024; 29(5):992. https://doi.org/10.3390/molecules29050992
Chicago/Turabian StyleLv, Jiahui, Qianqian Du, Suying Shi, Mengzhen Ma, Wei Zhang, Dezhu Ge, Lihua Xing, and Nianjun Yu. 2024. "Untargeted Metabolomics Based on UPLC-Q-Exactive-Orbitrap-MS/MS Revealed the Differences and Correlations between Different Parts of the Root of Paeonia lactiflora Pall" Molecules 29, no. 5: 992. https://doi.org/10.3390/molecules29050992
APA StyleLv, J., Du, Q., Shi, S., Ma, M., Zhang, W., Ge, D., Xing, L., & Yu, N. (2024). Untargeted Metabolomics Based on UPLC-Q-Exactive-Orbitrap-MS/MS Revealed the Differences and Correlations between Different Parts of the Root of Paeonia lactiflora Pall. Molecules, 29(5), 992. https://doi.org/10.3390/molecules29050992