
Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Conventional Treatment
2.1. 5-Fluorouracil
2.2. Photodynamic Therapy
2.3. Laser Therapy
2.4. Cryosurgery
2.5. Hedgehog Inhibitors
2.6. Immunotherapy
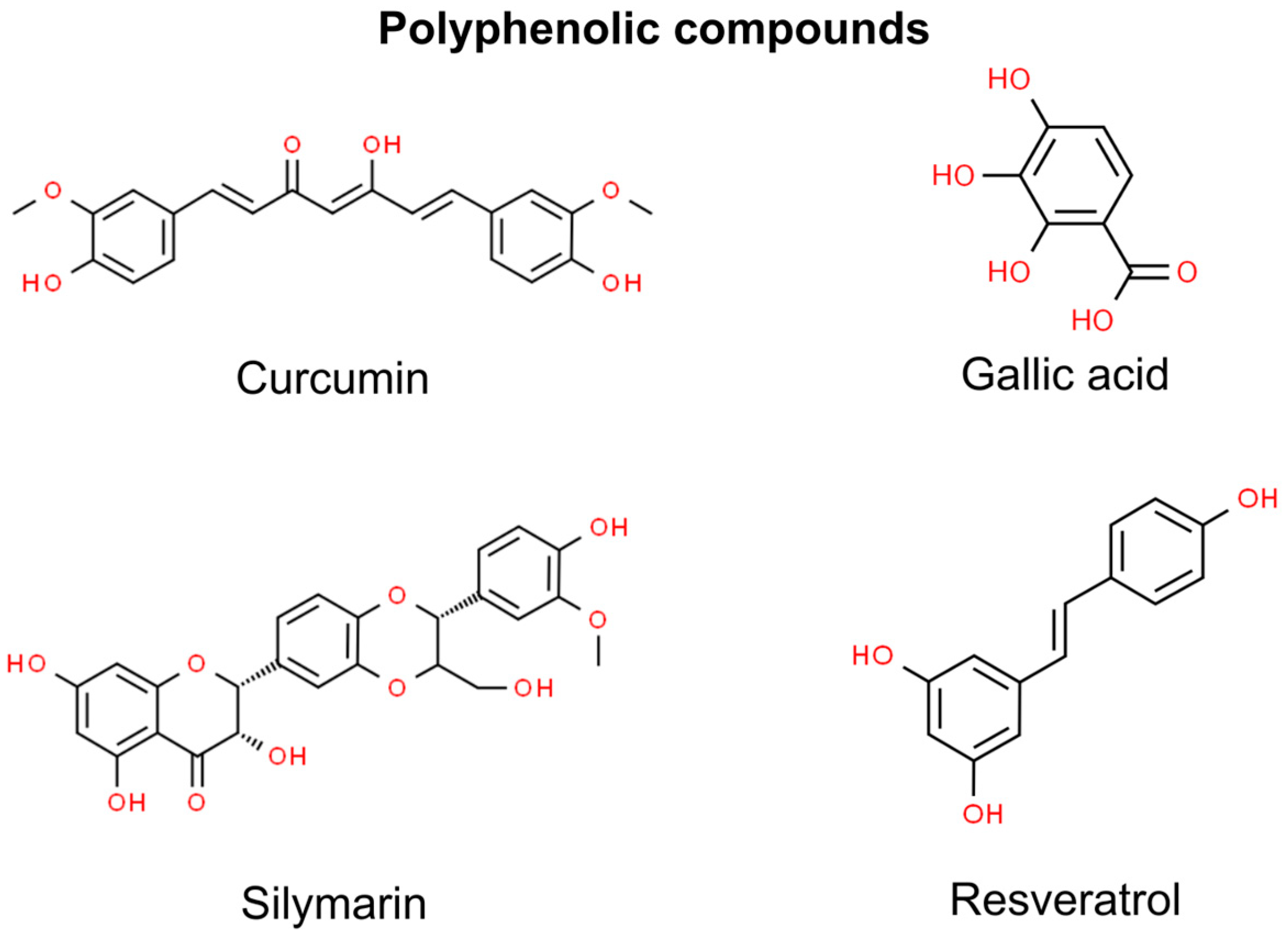
3. Polyphenolic Compounds
3.1. Curcumin
3.2. Gallic Acid
3.3. Silymarin
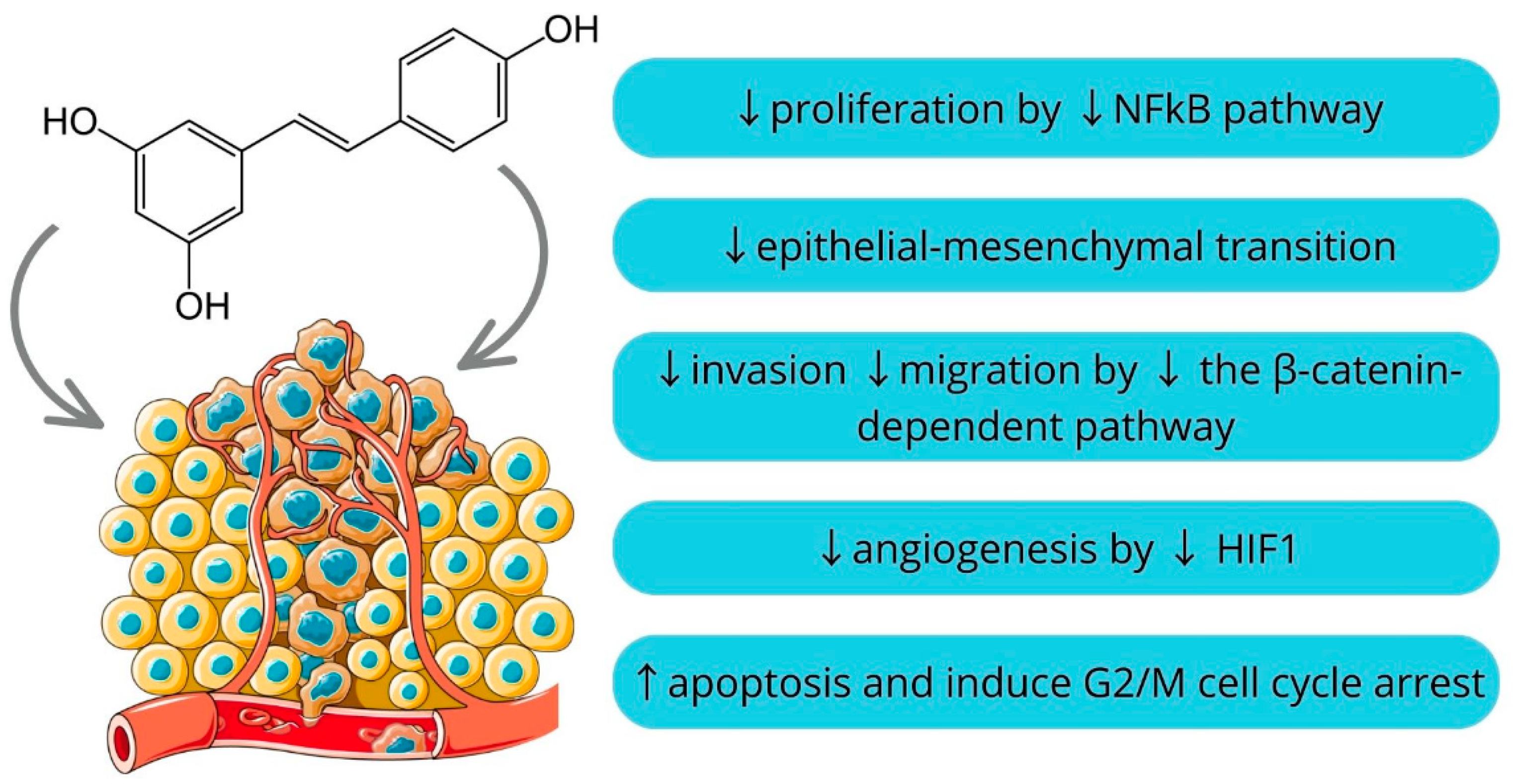
3.4. Resveratrol
Resveratrol and Ursolic Acid
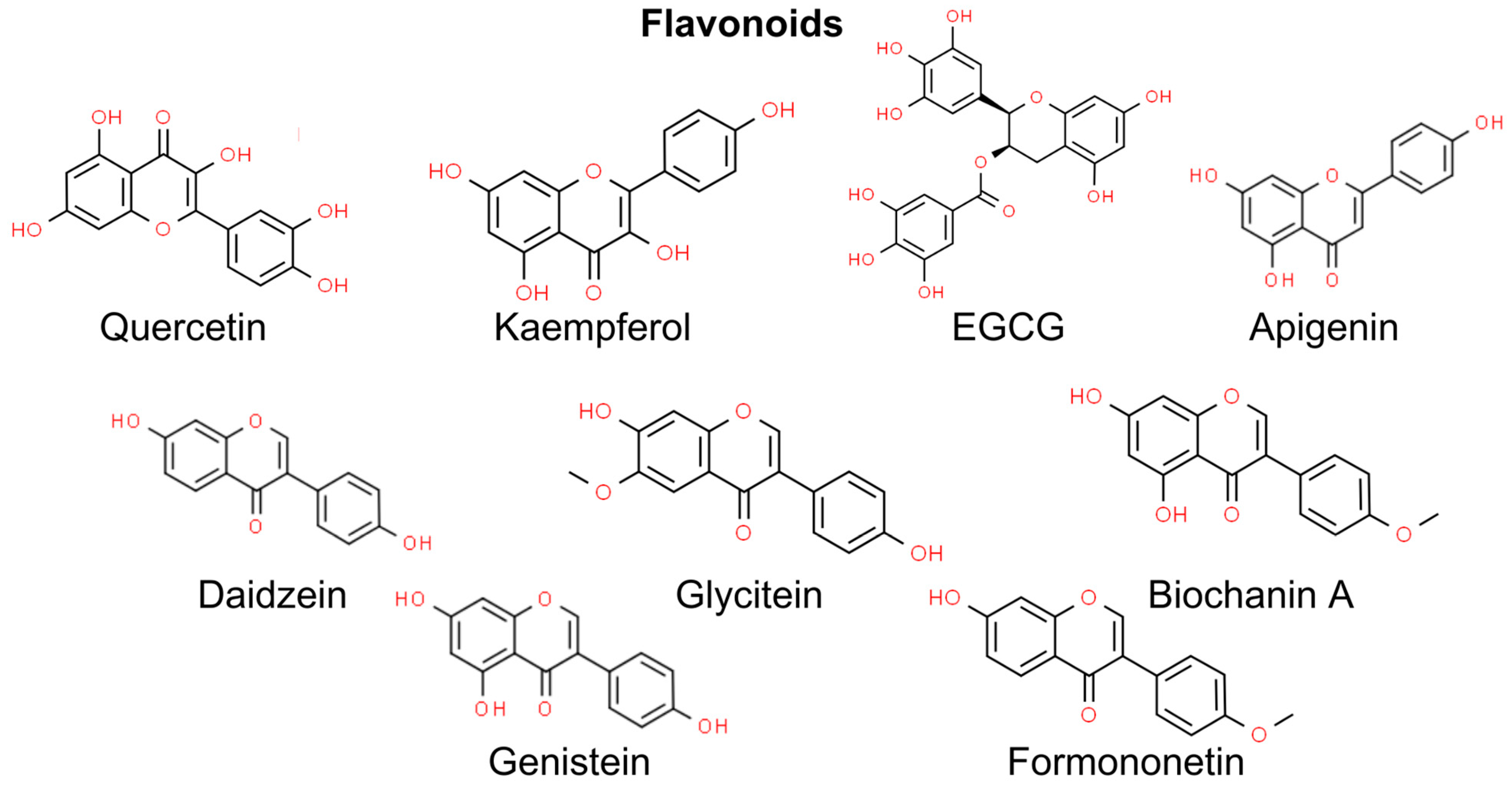
4. Flavonoids
4.1. Quercetin
4.2. Kaempferol
4.3. Epigallocatechin Gallate and Gallocatechin Gallate
4.4. Apigenin
4.5. Isoflavonoids
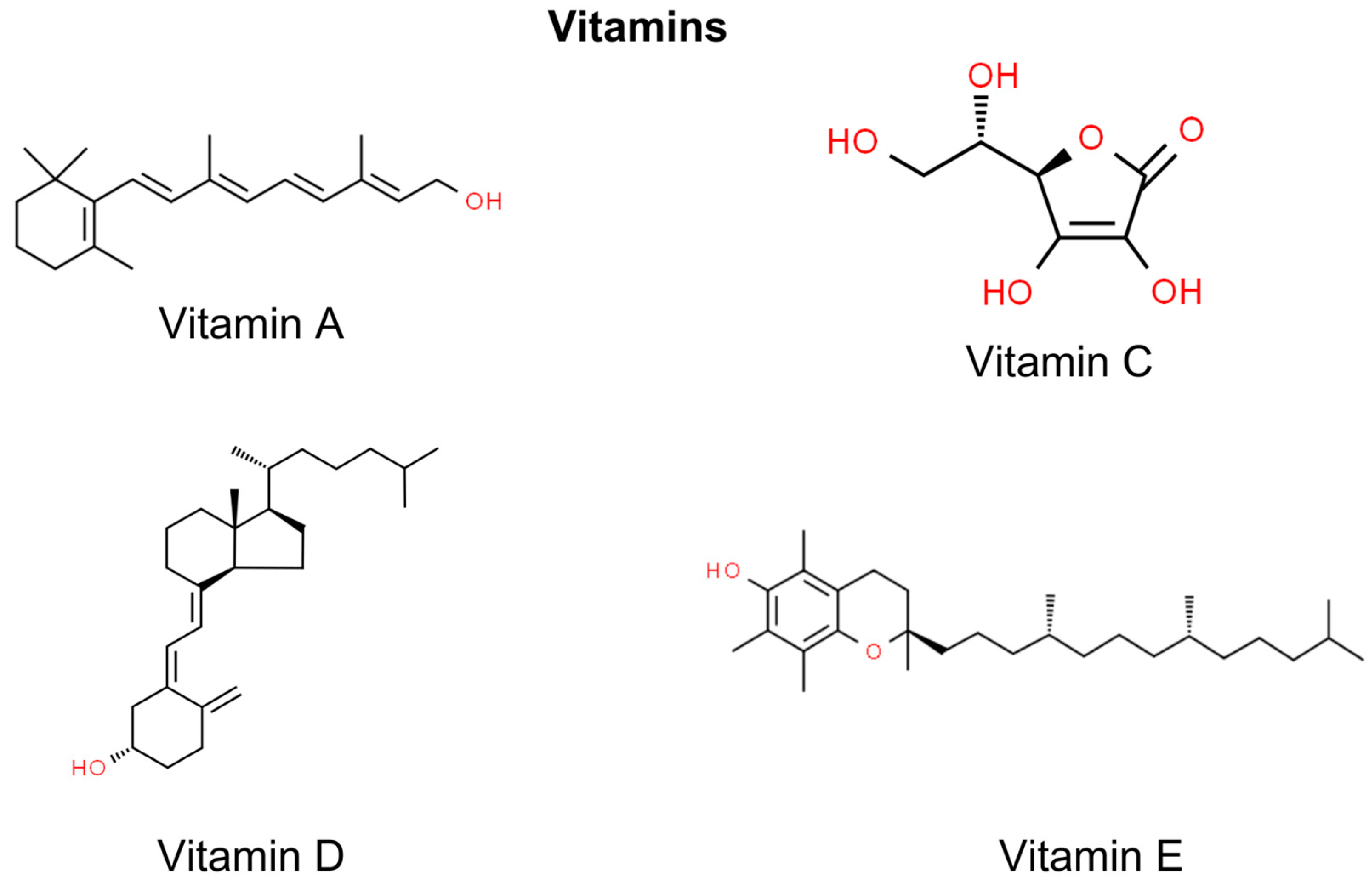
5. Vitamins
5.1. Vitamin A
5.2. Vitamin C
5.3. Vitamin E
5.4. Vitamin D
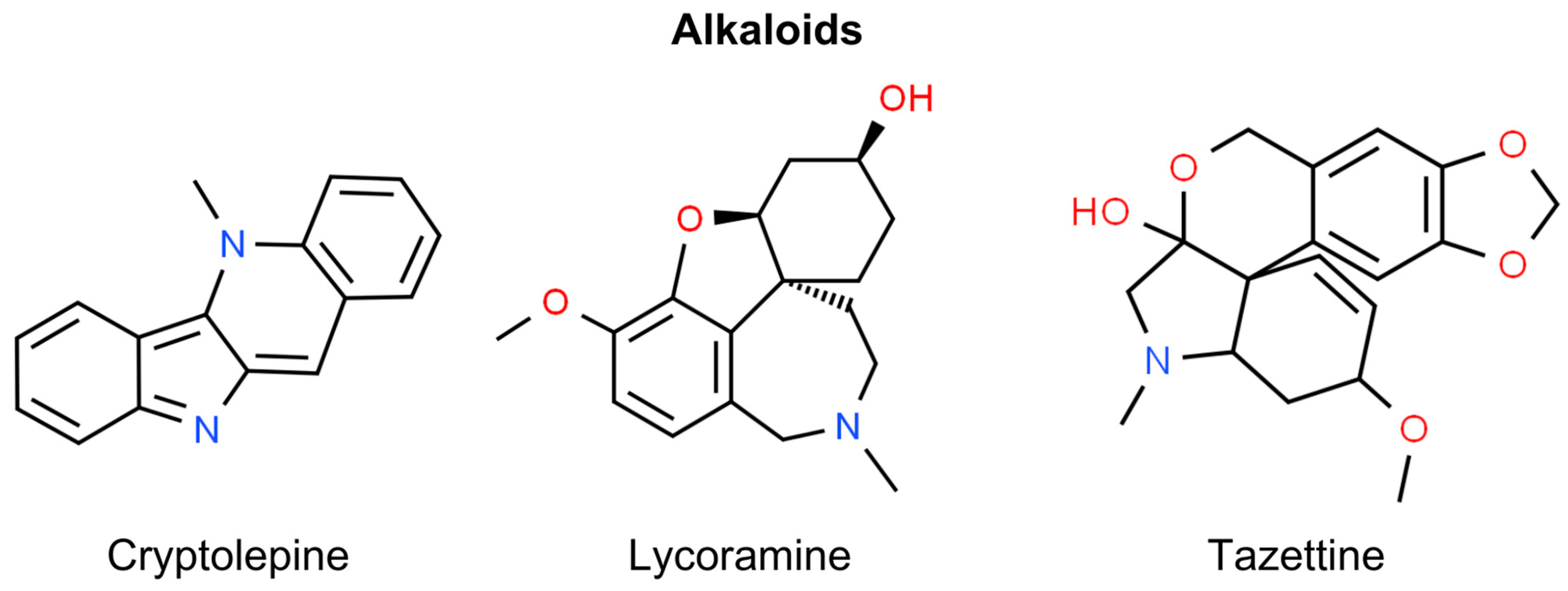
6. Alkaloids
Cryptolepine
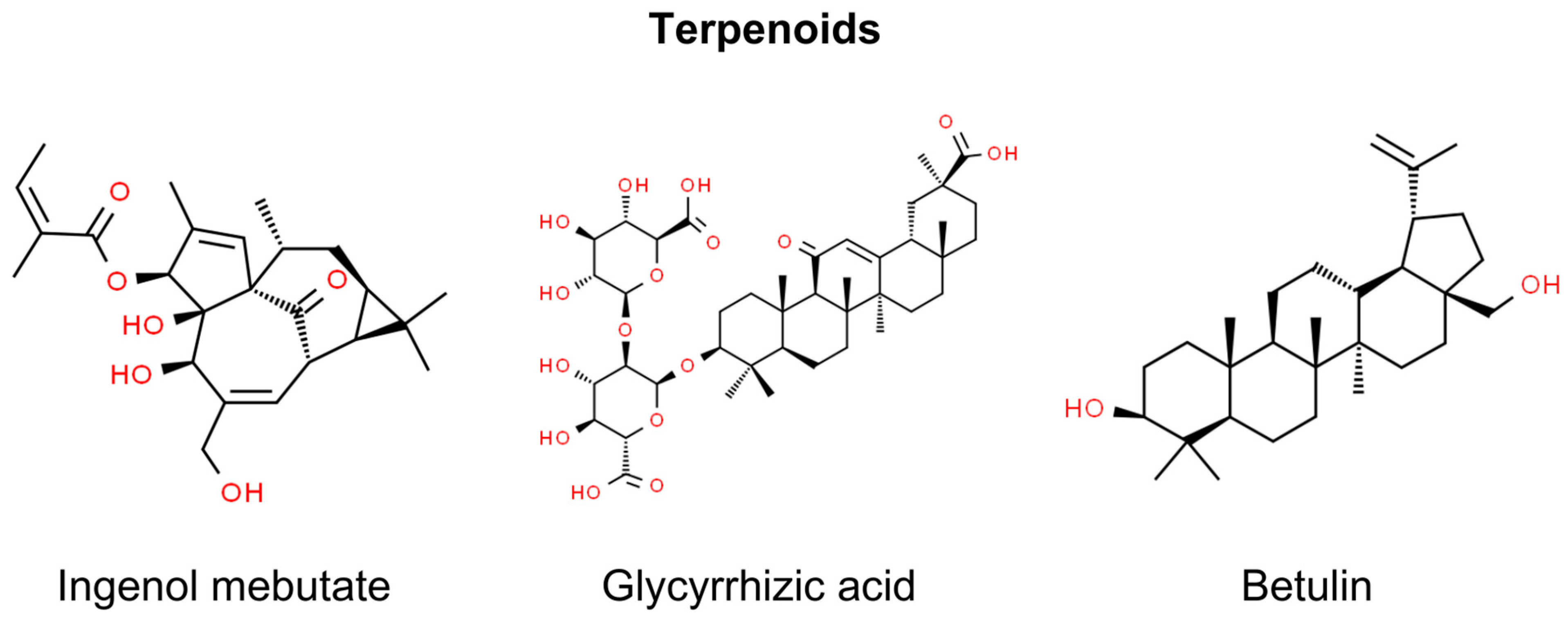
7. Terpenoids
7.1. Ingenol Mebutate
7.2. Glycyrrhizic Acid
7.3. Betulin and Betulinic Acid
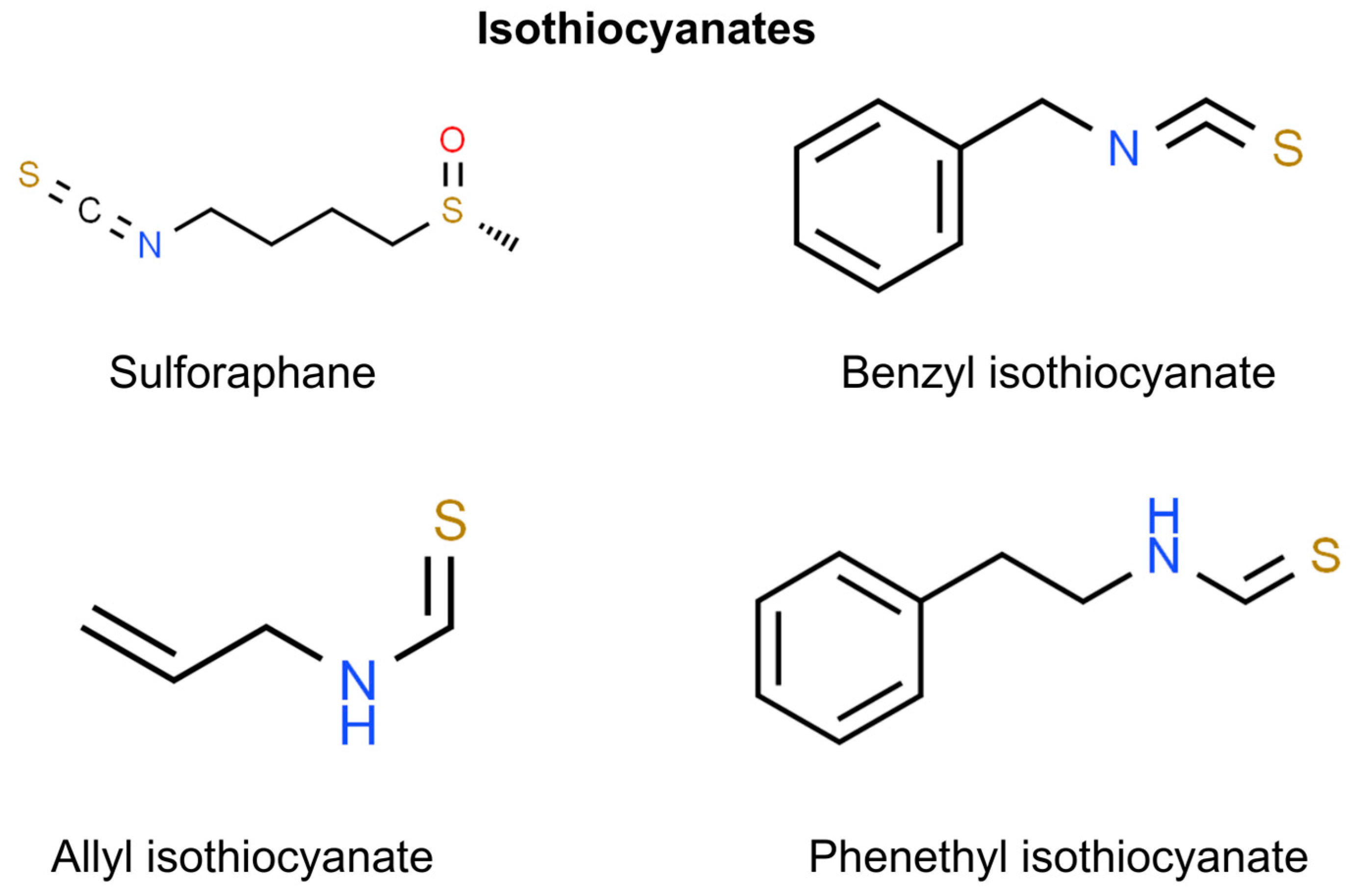
8. Isothiocyanates
Sulforaphane
9. Cannabinoids
10. Carotenoids
10.1. Lycopene
10.2. Astaxanthin and Fucoxanthin
11. Ceramides
12. Wound Healing and Skin Regeneration
13. Reactive Oxygen Species and NMSC
14. Methodology
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bosch, R.; Philips, N.; Suárez-Pérez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxidants 2015, 4, 248. [Google Scholar] [CrossRef]
- Madan, V.; Lear, J.T.; Szeimies, R.M. Non-Melanoma Skin Cancer. Lancet 2010, 375, 673–685. [Google Scholar] [CrossRef]
- Collins, L.; Quinn, A.; Stasko, T. Skin Cancer and Immunosuppression. Dermatol. Clin. 2019, 37, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Koh, D.; Wang, H.; Lee, J.; Chia, K.S.; Lee, H.P.; Goh, C.L. Basal Cell Carcinoma, Squamous Cell Carcinoma and Melanoma of the Skin: Analysis of the Singapore Cancer Registry Data 1968–1997. Br. J. Dermatol. 2003, 148, 1161–1166. [Google Scholar] [CrossRef]
- Bhandari, P.R.; Pai, V.V. Novel Medical Strategies Combating Nonmelanoma Skin Cancer. Indian. J. Dermatol. 2014, 59, 531. [Google Scholar] [CrossRef]
- Joseph, K. Non-Melanoma Skin Cancer—Overview. Curr. Cancer Ther. Rev. 2016, 12, 142–151. [Google Scholar] [CrossRef]
- Sellheyer, K. Basal Cell Carcinoma: Cell of Origin, Cancer Stem Cell Hypothesis and Stem Cell Markers. Br. J. Dermatol. 2011, 164, 696–711. [Google Scholar] [CrossRef]
- Gailani, M.R.; Stahle-Backdahl, M.; Leffell, D.J.; Glynn, M.; Zaphiropoulos, P.G.; Pressman, C.; Unden, A.B.; Dean, M.; Brash, D.E.; Bale, A.E.; et al. The Role of the Human Homologue of Drosophila Patched in Sporadic Basal Cell Carcinomas. Nat. Genet. 1996, 14, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Athar, M.; Li, C.; Kim, A.L.; Spiegelman, V.S.; Bickers, D.R. Sonic Hedgehog Signaling in Basal Cell Nevus Syndrome. Cancer Res. 2014, 74, 4967–4975. [Google Scholar] [CrossRef]
- Figueras Nart, I.; Cerio, R.; Dirschka, T.; Dréno, B.; Lear, J.T.; Pellacani, G.; Peris, K.; Ruiz de Casas, A. Defining the Actinic Keratosis Field: A Literature Review and Discussion. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 544–563. [Google Scholar] [CrossRef] [PubMed]
- Criscione, V.D.; Weinstock, M.A.; Naylor, M.F.; Luque, C.; Eide, M.J.; Bingham, S.F. Actinic Keratoses: Natural History and Risk of Malignant Transformation in the Veterans Affairs Topical Tretinoin Chemoprevention Trial. Cancer 2009, 115, 2523–2530. [Google Scholar] [CrossRef]
- Martincorena, I.; Roshan, A.; Gerstung, M.; Ellis, P.; Van Loo, P.; McLaren, S.; Wedge, D.C.; Fullam, A.; Alexandrov, L.B.; Tubio, J.M.; et al. High Burden and Pervasive Positive Selection of Somatic Mutations in Normal Human Skin. Science 2015, 348, 880. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.P.; Henning, S.; Faust, A.; Boukamp, P.; Volkmer, B.; Greinert, R. UVA-Induced Epigenetic Regulation of P16(INK4a) in Human Epidermal Keratinocytes and Skin Tumor Derived Cells. Photochem. Photobiol. Sci. 2012, 11, 180–190. [Google Scholar] [CrossRef] [PubMed]
- de Lima, P.O.; Joseph, S.; Panizza, B.; Simpson, F. Epidermal Growth Factor Receptor’s Function in Cutaneous Squamous Cell Carcinoma and Its Role as a Therapeutic Target in the Age of Immunotherapies. Curr. Treat. Options Oncol. 2020, 21, 9. [Google Scholar] [CrossRef] [PubMed]
- Tyler, L.N.; Ai, L.; Zuo, C.; Fan, C.Y.; Smoller, B.R. Analysis of Promoter Hypermethylation of Death-Associated Protein Kinase and P16 Tumor Suppressor Genes in Actinic Keratoses and Squamous Cell Carcinomas of the Skin. Mod. Pathol. 2003, 16, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Chitsazzadeh, V.; Coarfa, C.; Drummond, J.A.; Nguyen, T.; Joseph, A.; Chilukuri, S.; Charpiot, E.; Adelmann, C.H.; Ching, G.; Nguyen, T.N.; et al. Cross-Species Identification of Genomic Drivers of Squamous Cell Carcinoma Development across Preneoplastic Intermediates. Nat. Commun. 2016, 7, 12601. [Google Scholar] [CrossRef]
- Gatti, V.; Fierro, C.; Annicchiarico-Petruzzelli, M.; Melino, G.; Peschiaroli, A. ΔNp63 in Squamous Cell Carcinoma: Defining the Oncogenic Routes Affecting Epigenetic Landscape and Tumour Microenvironment. Mol. Oncol. 2019, 13, 981. [Google Scholar] [CrossRef]
- Darido, C.; Georgy, S.R.; Wilanowski, T.; Dworkin, S.; Auden, A.; Zhao, Q.; Rank, G.; Srivastava, S.; Finlay, M.J.; Papenfuss, A.T.; et al. Targeting of the Tumor Suppressor GRHL3 by a MiR-21-Dependent Proto-Oncogenic Network Results in PTEN Loss and Tumorigenesis. Cancer Cell 2011, 20, 635–648. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, B.; Wen, X.; Hao, D.; Du, D.; He, G.; Jiang, X. The Roles of LncRNA in Cutaneous Squamous Cell Carcinoma. Front. Oncol. 2020, 10, 158. [Google Scholar] [CrossRef]
- Darr, O.A.; Colacino, J.A.; Tang, A.L.; McHugh, J.B.; Bellile, E.L.; Bradford, C.R.; Prince, M.P.; Chepeha, D.B.; Rozek, L.S.; Moyer, J.S. Epigenetic Alterations in Metastatic Cutaneous Carcinoma. Head Neck 2015, 37, 994. [Google Scholar] [CrossRef]
- Dickinson, S.E.; Rusche, J.J.; Bec, S.L.; Horn, D.J.; Janda, J.; Rim, S.H.; Smith, C.L.; Bowden, G.T. The Effect of Sulforaphane on Histone Deacetylase Activity in Keratinocytes: Differences between in Vitro and in Vivo Analyses. Mol. Carcinog. 2015, 54, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Paquet, M.; Villanueva, E.; Brintnell, W. Interventions for Actinic Keratoses. Cochrane Database Syst. Rev. 2012, 12, CD004415. [Google Scholar] [CrossRef]
- Chetty, P.; Choi, F.; Mitchell, T. Primary Care Review of Actinic Keratosis and Its Therapeutic Options: A Global Perspective. Dermatol. Ther. 2015, 5, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.A.; Korgavkar, K.; Weinstock, M.A. Current Perspective on Actinic Keratosis: A Review. Br. J. Dermatol. 2017, 177, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Berman, B.; Cockerell, C.J.; Zografos, P. Pathobiology of Actinic Keratosis: Ultraviolet-Dependent Keratinocyte Proliferation. J. Am. Acad. Dermatol. 2013, 68, S10–S19. [Google Scholar] [CrossRef] [PubMed]
- Padilla, R.S.; Sebastian, S.; Jiang, Z.; Nindl, I.; Larson, R. Gene Expression Patterns of Normal Human Skin, Actinic Keratosis, and Squamous Cell Carcinoma: A Spectrum of Disease Progression. Arch. Dermatol. 2010, 146, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.; Schmieder, G.J.; Werschler, W.P.; Tschen, E.H.; Ling, M.R.; Stough, D.B.; Katsamas, J. Randomized, Double-Blind, Double-Dummy, Vehicle-Controlled Study of Ingenol Mebutate Gel 0.025% and 0.05% for Actinic Keratosis. J. Am. Acad. Dermatol. 2009, 60, 934–943. [Google Scholar] [CrossRef]
- Ratushny, V.; Gober, M.D.; Hick, R.; Ridky, T.W.; Seykora, J.T. From Keratinocyte to Cancer: The Pathogenesis and Modeling of Cutaneous Squamous Cell Carcinoma. J. Clin. Investig. 2012, 122, 464. [Google Scholar] [CrossRef]
- Dubois-Pot-Schneider, H.; Khairallah, G.; Brzenczek, C.; Plénat, F.; Marchal, F.; Amouroux, M. Transcriptomic Study on Human Skin Samples: Identification of Two Subclasses of Actinic Keratoses. Int. J. Mol. Sci. 2023, 24, 5937. [Google Scholar] [CrossRef]
- Firnhaber, J.M. Basal Cell and Cutaneous Squamous Cell Carcinomas: Diagnosis and Treatment. Am. Fam. Physician 2020, 102, 339–346. [Google Scholar]
- Reinehr, C.P.H.; Bakos, R.M. Actinic Keratoses: Review of Clinical, Dermoscopic, and Therapeutic Aspects. An. Bras. Dermatol. 2019, 94, 637–657. [Google Scholar] [CrossRef]
- Rosen, T.; Lebwohl, M.G.; Zografos, P. Prevalence and Awareness of Actinic Keratosis: Barriers and Opportunities. J. Am. Acad. Dermatol. 2013, 68, S2. [Google Scholar] [CrossRef]
- Canadian Cancer Society’s Advisory Committee on Cancer Statistics. Canadian Cancer Statistics 2015 Special Topic: Predictions of the Future Burden of Cancer in Canada; Canadian Cancer Society’s Advisory Committee on Cancer Statistics: Toronto, ON, Canada, 2015. [Google Scholar]
- Steeb, T.; Petzold, A.; Hornung, A.; Wessely, A.; Berking, C.; Heppt, M.V. Spontaneous Regression Rates of Actinic Keratosis: A Systematic Review and Pooled Analysis of Randomized Controlled Trials. Sci. Rep. 2022, 12, 5884. [Google Scholar] [CrossRef] [PubMed]
- King, S.C.; Chen, S. Analyzing the Cost of Preventing Nonmelanoma Skin Cancer. J. Investig. Dermatol. 2009, 129, 2745–2746. [Google Scholar] [CrossRef] [PubMed]
- Ramchatesingh, B.; Martínez Villarreal, A.; Arcuri, D.; Lagacé, F.; Setah, S.A.; Touma, F.; Al-Badarin, F.; Litvinov, I.V. The Use of Retinoids for the Prevention and Treatment of Skin Cancers: An Updated Review. Int. J. Mol. Sci. 2022, 23, 12622. [Google Scholar] [CrossRef] [PubMed]
- Shalhout, S.Z.; Emerick, K.S.; Kaufman, H.L.; Miller, D.M. Immunotherapy for Non-Melanoma Skin Cancer. Curr. Oncol. Rep. 2021, 23, 125. [Google Scholar] [CrossRef]
- Sharon, E.; Snast, I.; Lapidoth, M.; Kaftory, R.; Mimouni, D.; Hodak, E.; Levi, A. Laser Treatment for Non-Melanoma Skin Cancer: A Systematic Review and Meta-Analysis. Am. J. Clin. Dermatol. 2021, 22, 25–38. [Google Scholar] [CrossRef]
- Szepietowski, J.; Baran, W. Terapia w Dermatologii, 1st ed.; PZWL: Warszawa, Poland, 2019; Volume 1. [Google Scholar]
- Nahm, W.J.; Shen, J.; Zito, P.M.; Gonzalez, A.M.; Nagrani, N.; Moore, K.; Badiavas, E.V.; Kirsner, R.S.; Nichols, A.J. A Non-Surgical and Cost-Effective Treatment Approach Employing Topical Imiquimod, 5-Fluorouracil, and Tretinoin for Primary Non-Melanoma Skin Cancers. J. Drugs Dermatol. 2021, 20, 260–267. [Google Scholar] [CrossRef]
- Gracia-Cazaña, T.; González, S.; Gilaberte, Y. Resistance of Nonmelanoma Skin Cancer to Nonsurgical Treatments. Part I: Topical Treatments. Actas Dermo-Sifiliogr. (Engl. Ed.) 2016, 107, 730–739. [Google Scholar] [CrossRef]
- Souto, E.B.; da Ana, R.; Vieira, V.; Fangueiro, J.F.; Dias-Ferreira, J.; Cano, A.; Zielińska, A.; Silva, A.M.; Staszewski, R.; Karczewski, J. Non-Melanoma Skin Cancers: Physio-Pathology and Role of Lipid Delivery Systems in New Chemotherapeutic Treatments. Neoplasia 2022, 30, 100810. [Google Scholar] [CrossRef]
- Wang, Q.; Atluri, K.; Tiwari, A.K.; Babu, R.J. Exploring the Application of Micellar Drug Delivery Systems in Cancer Nanomedicine. Pharmaceuticals 2023, 16, 433. [Google Scholar] [CrossRef] [PubMed]
- Prince, G.T.; Cameron, M.C.; Fathi, R.; Alkousakis, T. Topical 5-Fluorouracil in Dermatologic Disease. Int. J. Dermatol. 2018, 57, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Searle, T.; Al-Niaimi, F.; Ali, F.R. 5-Fluorouracil in Dermatology: The Diverse Uses beyond Malignant and Premalignant Skin Disease. Dermatol. Surg. 2021, 47, E66–E70. [Google Scholar] [CrossRef]
- Ceilley, R.I. Mechanisms of Action of Topical 5-Fluorouracil: Review and Implications for the Treatment of Dermatological Disorders. J. Dermatol. Treat. 2012, 23, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sethy, C.; Kundu, C.N. 5-Fluorouracil (5-FU) Resistance and the New Strategy to Enhance the Sensitivity against Cancer: Implication of DNA Repair Inhibition. Biomed. Pharmacother. 2021, 137, 111285. [Google Scholar] [CrossRef]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic Therapy—Mechanisms, Photosensitizers and Combinations. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef]
- Solban, N.; Ortel, B.; Pogue, B.; Hasan, T. Targeted Optical Imaging and Photodynamic Therapy. In Molecular Imaging; Bogdanov, A.A., Licha, K., Eds.; Ernst Schering Research Foundation Workshop; Springer: Berlin/Heidelberg, Germany, 2005; Volume 49, pp. 229–258. [Google Scholar] [CrossRef]
- Juarranz, Á.; Jaén, P.; Sanz-Rodríguez, F.; Cuevas, J.; González, S. Photodynamic Therapy of Cancer. Basic Principles and Applications. Clin. Transl. Oncol. 2008, 10, 148–154. [Google Scholar] [CrossRef]
- Miles, J.; Luis-Gronau, C.; Cruzval-O’Reilly, E.; Lugo-Somolinos, A.; Jolly, P.S. Same Day Biopsy and Treatment of Non-Melanoma Skin Cancer in Patients with Field Cancerization: A Retrospective Chart Study. J. Skin. Cancer 2023, 2023, 9990046. [Google Scholar] [CrossRef]
- Gallego-Rentero, M.; Gutiérrez-Pérez, M.; Fernández-Guarino, M.; Mascaraque, M.; Portillo-Esnaola, M.; Gilaberte, Y.; Carrasco, E.; Juarranz, Á. Tgfβ1 Secreted by Cancer-Associated Fibroblasts as an Inductor of Resistance to Photodynamic Therapy in Squamous Cell Carcinoma Cells. Cancers 2021, 13, 5613. [Google Scholar] [CrossRef]
- Anfray, C.; Ummarino, A.; Andón, F.T.; Allavena, P. Current Strategies to Target Tumor-Associated-Macrophages to Improve Anti-Tumor Immune Responses. Cells 2020, 9, 46. [Google Scholar] [CrossRef]
- Fahradyan, A.; Howell, A.C.; Wolfswinkel, E.M.; Tsuha, M.; Sheth, P.; Wong, A.K. Updates on the Management of Non-Melanoma Skin Cancer (NMSC). Healthcare 2017, 5, 82. [Google Scholar] [CrossRef]
- Soleymani, T.; Abrouk, M.; Kelly, K.M. An Analysis of Laser Therapy for the Treatment of Nonmelanoma Skin Cancer. Dermatol. Surg. 2017, 43, 615–624. [Google Scholar] [CrossRef]
- Samarasinghe, V.; Madan, V. Nonmelanoma Skin Cancer. J. Cutan. Aesthet. Surg. 2012, 5, 3. [Google Scholar] [CrossRef]
- Samain, A.; Boullié, M.C.; Duval-Modeste, A.B.; Joly, P. Cryosurgery and Curettage-Cryosurgery for Basal Cell Carcinomas of the Mid-Face. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1291–1296. [Google Scholar] [CrossRef]
- Peikert, J.M. Prospective Trial of Curettage and Cryosurgery in the Management of Non-Facial, Superficial, and Minimally Invasive Basal and Squamous Cell Carcinoma. Int. J. Dermatol. 2011, 50, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Kuflik, E.G. Cryosurgery for Skin Cancer: 30-Year Experience and Cure Rates. Dermatol. Surg. 2004, 30, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Ferry, A.M.; Sarrami, S.M.; Hollier, P.C.; Gerich, C.F.; Thornton, J.F. Treatment of Non-Melanoma Skin Cancers in the Absence of Mohs Micrographic Surgery. Plast. Reconstr. Surg. Glob. Open 2020, 8, E3300. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.S.; Bar, A. Basal Cell Carcinoma. Dermatol. Clin. 2023, 41, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Schadendorf, D. Update in the Treatment of Non-Melanoma Skin Cancers: The Use of PD-1 Inhibitors in Basal Cell Carcinoma and Cutaneous Squamous-Cell Carcinoma. J. Immunother. Cancer 2022, 10, e005082. [Google Scholar] [CrossRef] [PubMed]
- Calienni, M.N.; Febres-Molina, C.; Llovera, R.E.; Zevallos-Delgado, C.; Tuttolomondo, M.E.; Paolino, D.; Fresta, M.; Barazorda-Ccahuana, H.L.; Gómez, B.; Alonso, S.d.V.; et al. Nanoformulation for Potential Topical Delivery of Vismodegib in Skin Cancer Treatment. Int. J. Pharm. 2019, 565, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Strategie Molekularne w Leczeniu Raków Skóry, Adam Włodarkiewicz—Przegląd Dermatologiczny 2/2012. Available online: https://www.termedia.pl/Strategie-molekularne-w-leczeniu-rakow-skory,56,18519,0,0.html (accessed on 26 January 2024).
- Shalhout, S.Z.; Kaufman, H.L.; Emerick, K.S.; Miller, D.M. Immunotherapy for Nonmelanoma Skin Cancer: Facts and Hopes. Clin. Cancer Res. 2022, 28, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Fettouh, N.; Kubicki, S.L.; Chen, L.; Silapunt, S.; Migden, M.R. Targeted Therapy and Immunotherapy in Nonmelanoma Skin Cancer. Dermatol. Clin. 2023, 41, 23–37. [Google Scholar] [CrossRef]
- Wilde, D.C.; Glaun, M.E.; Wong, M.K.; Gross, N.D. Neoadjuvant Approaches to Non-Melanoma Skin Cancer. Cancers 2023, 15, 5494. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of Dietary Polyphenols: The Role of Metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Cherian, E.; Goyal, M.; Mittal, N.; Yesodharan, V.; Ramadoss, R.; Thomas, C. Assessment of Therapeutic Bio-Activity of Cinnamoyl Sulfonamide Hydroxamate in Squamous Cell Carcinoma. Cureus 2023, 15, e43949. [Google Scholar] [CrossRef]
- Lee, C.H.; Tsao, Y.H.; Weng, Y.P.; Wang, I.C.; Chen, Y.P.; Hung, P.F. Therapeutic Effects of Perilla Phenols in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2023, 24, 14931. [Google Scholar] [CrossRef] [PubMed]
- Waer, C.N.; Kaur, P.; Tumur, Z.; Dee Hui, D.; Le, B.; Guerra, C.; Henson, B.; Seleem, D.; Lewis, J. Rosmarinic Acid/ Blue Light Combination Treatment Inhibits Head and Neck Squamous Cell Carcinoma In Vitro. Anticancer Res. 2020, 40, 751–758. [Google Scholar] [CrossRef]
- Almosa, H.; Alqriqri, M.; Denetiu, I.; Baghdadi, M.A.; Alkhaled, M.; Alhosin, M.; Aldajani, W.A.; Zamzami, M.; Ucisik, M.H.; Damiati, S. Cytotoxicity of Standardized Curcuminoids Mixture against Epithelial Ovarian Cancer Cell Line SKOV-3. Sci. Pharm. 2020, 88, 11. [Google Scholar] [CrossRef]
- Shakeri, A.; Ward, N.; Panahi, Y.; Sahebkar, A. Anti-Angiogenic Activity of Curcumin in Cancer Therapy: A Narrative Review. Curr. Vasc. Pharmacol. 2018, 17, 262–269. [Google Scholar] [CrossRef]
- Anand, P.; Thomas, S.G.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Sung, B.; Tharakan, S.T.; Misra, K.; Priyadarsini, I.K.; Rajasekharan, K.N.; et al. Biological Activities of Curcumin and Its Analogues (Congeners) Made by Man and Mother Nature. Biochem. Pharmacol. 2008, 76, 1590–1611. [Google Scholar] [CrossRef]
- Li, M.; Guo, T.; Lin, J.; Huang, X.; Ke, Q.; Wu, Y.; Fang, C.; Hu, C. Curcumin Inhibits the Invasion and Metastasis of Triple Negative Breast Cancer via Hedgehog/Gli1 Signaling Pathway. J. Ethnopharmacol. 2022, 283, 114689. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-d.; Pang, Y.-x.; Zhao, X.-r.; Li, R.; Jin, C.-j.; Xue, J.; Dong, R.-y.; Liu, P.-s. Curcumin Induces Apoptotic Cell Death and Protective Autophagy by Inhibiting AKT/MTOR/P70S6K Pathway in Human Ovarian Cancer Cells. Arch. Gynecol. Obstet. 2019, 299, 1627–1639. [Google Scholar] [CrossRef]
- Zhu, M.; Zheng, Z.; Huang, J.; Ma, X.; Huang, C.; Wu, R.; Li, X.; Liang, Z.; Deng, F.; Wu, J.; et al. Modulation of MiR-34a in Curcumin-Induced Antiproliferation of Prostate Cancer Cells. J. Cell. Biochem. 2019, 120, 15616–15624. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of Curcumin, a Component of Golden Spice, and Its Miraculous Biological Activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Sundaram, C.; Jhurani, S.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin and Cancer: An “Old-Age” Disease with an “Age-Old” Solution. Cancer Lett. 2008, 267, 133–164. [Google Scholar] [CrossRef]
- Xu, X.Y.; Meng, X.; Li, S.; Gan, R.Y.; Li, Y.; Li, H. Bin Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients 2018, 10, 1553. [Google Scholar] [CrossRef]
- Li, H.; Gao, A.; Jiang, N.; Liu, Q.; Liang, B.; Li, R.; Zhang, E.; Li, Z.; Zhu, H. Protective Effect of Curcumin Against Acute Ultraviolet B Irradiation-Induced Photo-Damage. Photochem. Photobiol. 2016, 92, 808–815. [Google Scholar] [CrossRef]
- Phillips, J.; Moore-Medlin, T.; Sonavane, K.; Ekshyyan, O.; McLarty, J.; Nathan, C.A.O. Curcumin Inhibits UV Radiation-Induced Skin Cancer in SKH-1 Mice. Otolaryngol. Head Neck Surg. 2013, 148, 797–803. [Google Scholar] [CrossRef]
- Khandelwal, A.R.; Rong, X.; Moore-Medlin, T.; Ekshyyan, O.; Abreo, F.; Gu, X.; Nathan, C.A.O. Photopreventive Effect and Mechanism of AZD4547 and Curcumin C3 Complex on UVB-Induced Epidermal Hyperplasia. Cancer Prev. Res. 2016, 9, 296–304. [Google Scholar] [CrossRef]
- Study Details|Curcumin Biomarker Trial in Head and Neck Cancer|ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/study/NCT01160302 (accessed on 2 December 2023).
- Latimer, B.; Ekshyyan, O.; Nathan, N.; Moore-Medlin, T.; Rong, X.; Ma, X.; Khandelwal, A.; Christy, H.T.; Abreo, F.; McClure, G.; et al. Enhanced Systemic Bioavailability of Curcumin through Transmucosal Administration of a Novel Microgranular Formulation. Anticancer Res. 2015, 35, 6411–6418. [Google Scholar]
- Naksuriya, O.; Okonogi, S.; Schiffelers, R.M.; Hennink, W.E. Curcumin Nanoformulations: A Review of Pharmaceutical Properties and Preclinical Studies and Clinical Data Related to Cancer Treatment. Biomaterials 2014, 35, 3365–3383. [Google Scholar] [CrossRef] [PubMed]
- Jose, A.; Labala, S.; Venuganti, V.V.K. Co-Delivery of Curcumin and STAT3 SiRNA Using Deformable Cationic Liposomes to Treat Skin Cancer. J. Drug Target. 2017, 25, 330–341. [Google Scholar] [CrossRef]
- Malathi, S.; Pavithra, P.S.; Sridevi, S.; Verma, R.S. Fabrication of Nanopatterned PLGA Films of Curcumin and TPGS for Skin Cancer. Int. J. Pharm. 2020, 578, 119100. [Google Scholar] [CrossRef]
- Chaudhari, S.P.; Tam, A.Y.; Barr, J.A. Curcumin: A Contact Allergen. J. Clin. Aesthet. Dermatol. 2015, 8, 43–48. [Google Scholar] [PubMed]
- Yarla, N.S.; Bishayee, A.; Sethi, G.; Reddanna, P.; Kalle, A.M.; Dhananjaya, B.L.; Dowluru, K.S.V.G.K.; Chintala, R.; Duddukuri, G.R. Targeting Arachidonic Acid Pathway by Natural Products for Cancer Prevention and Therapy. Semin. Cancer Biol. 2016, 40–41, 48–81. [Google Scholar] [CrossRef]
- Jiang, Y.; Pei, J.; Zheng, Y.; Miao, Y.-J.; Duan, B.-Z.; Huang, L.-F. Gallic Acid: A Potential Anti-Cancer Agent. Chin. J. Integr. Med. 2022, 28, 661–671. [Google Scholar] [CrossRef] [PubMed]
- de Jesus, S.F.; de Souza, M.G.; Queiroz, L.d.R.P.; de Paula, D.P.S.; Tabosa, A.T.L.; Alves, W.S.M.; da Silveira, L.H.; Ferreira, A.T.d.S.; Martuscelli, O.J.D.; Farias, L.C.; et al. Gallic Acid Has an Inhibitory Effect on Skin Squamous Cell Carcinoma and Acts on the Heat Shock Protein HSP90AB1. Gene 2023, 851, 147041. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, L.; Saldías-Fuentes, C.; Carrasco, K.; Halpern, A.C.; Mao, J.J.; Navarrete-Dechent, C. Complementary and Alternative Therapies in Skin Cancer: A Literature Review of Biologically Active Compounds. Dermatol. Ther. 2022, 35, e15842. [Google Scholar] [CrossRef]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Gillessen, A.; Schmidt, H.H.-J. Silymarin as Supportive Treatment in Liver Diseases: A Narrative Review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.R.; Paudel, S.; Raina, K.; Agarwal, R. Silibinin and Non-Melanoma Skin Cancers. J. Tradit. Complement. Med. 2020, 10, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Kwah, M.X.Y.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.L.; Wang, L.; Ong, P.S.; et al. Resveratrol for Cancer Therapy: Challenges and Future Perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef]
- Hecker, A.; Schellnegger, M.; Hofmann, E.; Luze, H.; Nischwitz, S.P.; Kamolz, L.P.; Kotzbeck, P. The Impact of Resveratrol on Skin Wound Healing, Scarring, and Aging. Int. Wound J. 2022, 19, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxid. Med. Cell. Longev. 2015, 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Srivani, G.; Behera, S.K.; Dariya, B.; Aliya, S.; Alam, A.; Nagaraju, G.P. Resveratrol Binds and Inhibits Transcription Factor HIF-1α in Pancreatic Cancer. Exp. Cell Res. 2020, 394, 112126. [Google Scholar] [CrossRef]
- Ji, Q.; Liu, X.; Fu, X.; Zhang, L.; Sui, H.; Zhou, L.; Sun, J.; Cai, J.; Qin, J.; Ren, J.; et al. Resveratrol Inhibits Invasion and Metastasis of Colorectal Cancer Cells via MALAT1 Mediated Wnt/β-Catenin Signal Pathway. PLoS ONE 2013, 8, e78700. [Google Scholar] [CrossRef]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Evidence That TNF-β Induces Proliferation in Colorectal Cancer Cells and Resveratrol Can down-Modulate It. Exp. Biol. Med. 2019, 244, 1–12. [Google Scholar] [CrossRef]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol Suppresses Epithelial-to-Mesenchymal Transition in Colorectal Cancer through TGF-Β1/Smads Signaling Pathway Mediated Snail/E-Cadherin Expression. BMC Cancer 2015, 15, 97. [Google Scholar] [CrossRef]
- Yu, X.D.; Yang, J.-l.; Zhang, W.L.; Liu, D.X. Resveratrol Inhibits Oral Squamous Cell Carcinoma through Induction of Apoptosis and G2/M Phase Cell Cycle Arrest. Tumour Biol. 2016, 37, 2871–2877. [Google Scholar] [CrossRef]
- Ponzo, V.; Soldati, L.; Bo, S. Resveratrol: A Supplementation for Men or for Mice? J. Transl. Med. 2014, 12, 158. [Google Scholar] [CrossRef]
- Seo, D.Y.; Lee, S.R.; Heo, J.W.; No, M.H.; Rhee, B.D.; Ko, K.S.; Kwak, H.B.; Han, J. Ursolic Acid in Health and Disease. Korean J. Physiol. Pharmacol. 2018, 22, 235. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Tuli, H.S.; Sharma, A.K. Ursolic Acid (UA): A Metabolite with Promising Therapeutic Potential. Life Sci. 2016, 146, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Rho, O.; Junco, J.; Carbajal, S.; Siegel, D.; Slaga, T.J.; Di Giovanni, J. Effect of Combined Treatment with Ursolic Acid and Resveratrol on Skin Tumor Promotion by 12-O-Tetradecanoylphorbol-13-Acetate. Cancer Prev. Res. 2015, 8, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Junco, J.J.; Mancha, A.; Malik, G.; Wei, S.J.; Kim, D.J.; Liang, H.; Slaga, T.J. Resveratrol and P-Glycoprotein Inhibitors Enhance the Anti-Skin Cancer Effects of Ursolic Acid. Mol. Cancer Res. 2013, 11, 1521–1529. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Cheng, Y.; Gao, H.; Chen, X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules 2023, 28, 4982. [Google Scholar] [CrossRef]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic Potential of Flavonoids in Cancer: ROS-Mediated Mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Klllç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849. [Google Scholar] [CrossRef]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84. [Google Scholar] [CrossRef]
- Stevanato, R.; Bertelle, M.; Fabris, S. Photoprotective Characteristics of Natural Antioxidant Polyphenols. Regul. Toxicol. Pharmacol. 2014, 69, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Nan, W.; Ding, L.; Chen, H.; Khan, F.U.; Yu, L.; Sui, X.; Shi, X. Topical Use of Quercetin-Loaded Chitosan Nanoparticles against Ultraviolet B Radiation. Front. Pharmacol. 2018, 9, 826. [Google Scholar] [CrossRef]
- Chitkara, A.; Mangla, B.; Kumar, P.; Javed, S.; Ahsan, W.; Popli, H. Design-of-Experiments (DoE)-Assisted Fabrication of Quercetin-Loaded Nanoemulgel and Its Evaluation against Human Skin Cancer Cell Lines. Pharmaceutics 2022, 14, 2517. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Montaño, J.M.; Burgos-Moron, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Yao, K.; Chen, H.; Liu, K.; Langfald, A.; Yang, G.; Zhang, Y.; Yu, D.H.; Kim, M.O.; Lee, M.H.; Li, H.; et al. Kaempferol Targets RSK2 and MSK1 to Suppress UV Radiation-Induced Skin Cancer. Cancer Prev. Res. 2014, 7, 958–967. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Pezzani, R.; Redaelli, M.; Zorzan, M.; Imran, M.; Khalil, A.A.; Salehi, B.; Sharopov, F.; Cho, W.C.; Sharifi-Rad, J. Preclinical Pharmacological Activities of Epigallocatechin-3-Gallate in Signaling Pathways: An Update on Cancer. Molecules 2020, 25, 467. [Google Scholar] [CrossRef]
- Singh, T.; Katiyar, S.K. Green Tea Polyphenol, (-)-Epigallocatechin-3-Gallate, Induces Toxicity in Human Skin Cancer Cells by Targeting β-Catenin Signaling. Toxicol. Appl. Pharmacol. 2013, 273, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Mao, Q.; Yang, J.; Du, N.; Zhu, Y.; Min, W. (–)-Epigallocatechin-3-Gallate Protects Human Skin Fibroblasts from Ultraviolet a Induced Photoaging. Clin. Cosmet. Investig. Dermatol. 2023, 16, 149. [Google Scholar] [CrossRef]
- Sheng, Y.Y.; Xiang, J.; Lu, J.L.; Ye, J.H.; Chen, Z.J.; Zhao, J.W.; Liang, Y.R.; Zheng, X.Q. Protective Effects of Gallocatechin Gallate against Ultraviolet B Induced Skin Damages in Hairless Mice. Sci. Rep. 2022, 12, 1310. [Google Scholar] [CrossRef]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.; Zhang, Z.; Yin, M.; Chen, X.; Zhao, S.; Wu, L. Apigenin Induced Apoptosis by Downregulating Sulfiredoxin Expression in Cutaneous Squamous Cell Carcinoma. Oxid. Med. Cell. Longev. 2022, 2022, 8172866. [Google Scholar] [CrossRef]
- Li, L.; Li, M.; Xu, S.; Chen, H.; Chen, X.; Gu, H. Apigenin Restores Impairment of Autophagy and Downregulation of Unfolded Protein Response Regulatory Proteins in Keratinocytes Exposed to Ultraviolet B Radiation. J. Photochem. Photobiol. B 2019, 194, 84–95. [Google Scholar] [CrossRef]
- García-garcía, V.A.; Alameda, J.P.; Page, A.; Mérida-garcía, A.; Navarro, M.; Tejero, A.; Paramio, J.M.; García-fernández, R.A.; Casanova, M.L. IKKα Induces Epithelial-Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin. Int. J. Mol. Sci. 2022, 23, 1375. [Google Scholar] [CrossRef]
- Kiraly, A.J.; Soliman, E.; Jenkins, A.; Van Dross, R.T. Apigenin Inhibits COX-2, PGE2, and EP1 and Also Initiates Terminal Differentiation in the Epidermis of Tumor Bearing Mice. Prostaglandins Leukot. Essent. Fatty Acids 2016, 104, 44–53. [Google Scholar] [CrossRef]
- Desmawati, D.; Sulastri, D. Phytoestrogens and Their Health Effect. Open Access Maced. J. Med. Sci. 2019, 7, 495. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [PubMed]
- AV, S.; AH, H. Phytoestrogens and Their Effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef]
- Huang, P.H.; Hu, S.C.S.; Lee, C.W.; Yeh, A.C.; Tseng, C.H.; Yen, F.L. Design of Acid-Responsive Polymeric Nanoparticles for 7,3′,4′-Trihydroxyisoflavone Topical Administration. Int. J. Nanomed. 2016, 11, 1615–1627. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, C.H.; Lee, J.Y.; Yang, H.; Kim, J.H.; Park, J.H.Y.; Kim, J.E.; Lee, K.W. Topical Application of 7,3′,4′-Trihydroxyisoflavone Alleviates Atopic Dermatitis-Like Symptoms in NC/Nga Mice. Planta Med. 2020, 86, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.E.; Kim, T.; Yeom, M.H.; Park, J.S.; di Luccio, E.; Chen, H.; Dong, Z.; Lee, K.W.; Kang, N.J. 7,3′,4′-Trihydroxyisoflavone, a Metabolite of the Soy Isoflavone Daidzein, Suppresses α-Melanocyte-Stimulating Hormone-Induced Melanogenesis by Targeting Melanocortin 1 Receptor. Front. Mol. Biosci. 2020, 7, 577284. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.H.; Tseng, C.H.; Lin, C.Y.; Lee, C.W.; Yen, F.L. Preparation, Characterizations and Anti-Pollutant Activity of 7,3′,4′-Trihydroxyisoflavone Nanoparticles in Particulate Matter-Induced HaCaT Keratinocytes. Int. J. Nanomed. 2018, 13, 3279–3293. [Google Scholar] [CrossRef]
- Tang, S.C.; Hsiao, Y.P.; Ko, J.L. Genistein Protects against Ultraviolet B–Induced Wrinkling and Photoinflammation in in Vitro and in Vivo Models. Genes. Nutr. 2022, 17, 4. [Google Scholar] [CrossRef]
- Terra, V.A.; Souza-Neto, F.P.; Frade, M.A.C.; Ramalho, L.N.Z.; Andrade, T.A.M.; Pasta, A.A.C.; Conchon, A.C.; Guedes, F.A.; Luiz, R.C.; Cecchini, R.; et al. Genistein Prevents Ultraviolet B Radiation-Induced Nitrosative Skin Injury and Promotes Cell Proliferation. J. Photochem. Photobiol. B 2015, 144, 20–27. [Google Scholar] [CrossRef]
- Ramachandran, V.; V, I.K.; Hr, K.K.; Tiwari, R.; Tiwari, G. Biochanin-A: A Bioactive Natural Product with Versatile Therapeutic Perspectives. Curr. Drug Res. Rev. 2022, 14, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.G.; Kim, J.E.; Jung, S.K.; Li, Y.; Bode, A.M.; Park, J.S.; Yeom, M.H.; Dong, Z.; Lee, K.W. MLK3 Is a Direct Target of Biochanin A, Which Plays a Role in Solar UV-Induced COX-2 Expression in Human Keratinocytes. Biochem. Pharmacol. 2013, 86, 896. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, T.H.; Chen, C.P.; Xiang, J.J.; Zhao, X.B.; Gui, R.Y.; Liao, X.H. Targeting COX-2 Potently Inhibits Proliferation of Cancer Cells in Vivo but Not in Vitro in Cutaneous Squamous Cell Carcinoma. Transl. Cancer Res. 2021, 10, 2219. [Google Scholar] [CrossRef] [PubMed]
- McEldrew, E.P.; Lopez, M.J.; Milstein, H. Vitamin A; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Kennel, K.A.; Drake, M.T. Vitamin D in the Cancer Patient. Curr. Opin. Support. Palliat. Care 2013, 7, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Pullar, J.M.; Carr, A.C.; Vissers, M.C.M. The Roles of Vitamin C in Skin Health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Keen, M.A.; Hassan, I. Vitamin E in Dermatology. Indian. Dermatol. Online J. 2016, 7, 311. [Google Scholar] [CrossRef] [PubMed]
- Chinembiri, T.N.; du Plessis, L.H.; Gerber, M.; Hamman, J.H.; du Plessis, J. Review of Natural Compounds for Potential Skin Cancer Treatment. Molecules 2014, 19, 11679–11721. [Google Scholar] [CrossRef]
- Katta, R.; Brown, D.N. Diet and Skin Cancer: The Potential Role of Dietary Antioxidants in Nonmelanoma Skin Cancer Prevention. J. Skin. Cancer 2015, 2015, 893149. [Google Scholar] [CrossRef] [PubMed]
- Maalouf, S.; El-Sabban, M.; Darwiche, N.; Gali-Muhtasib, H. Protective Effect of Vitamin E on Ultraviolet B Light–Induced Damage in Keratinocytes. Mol. Carcinog. 2002, 34, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Offord, E.A.; Gautier, J.-C.; Avanti, O.; Scaletta, C.; Runge, F.; Krämer, K.; Applegate, L.A. Photoprotective Potential of Lycopene, Beta-Carotene, Vitamin E, Vitamin C and Carnosic Acid in UVA-Irradiated Human Skin Fibroblasts. Free Radic. Biol. Med. 2002, 32, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Ghaedi, E.; Rahrovani, F.; Javanbakht, M.H.; Ehsani, A.-H.; Esrafili, A.; Mohammadi, H.; Zarei, M.; Djalali, M. Retinol and α-Tocopherol Levels in the Serum and Subcutaneous Adipose Tissue of Newly Diagnosed Basal Cell Carcinoma Patients. Iran. J. Public. Health 2019, 48, 1838–1846. [Google Scholar] [CrossRef] [PubMed]
- Argos, M.; Dignam, J.J.; Parvez, F.; Rahman, M.; Hore, S.K.; Shahriar, M.H.; Sarwar, G.; Islam, T.; Slavkovich, V.; Graziano, J.; et al. A Randomized Trial of Selenium and Vitamin E for Primary Prevention of Non-Melanoma Skin Cancer: Trial Results and Experience from a Low-Resource Setting. J. Clin. Oncol. 2017, 35, 1511. [Google Scholar] [CrossRef]
- Karampinis, E.; Aloizou, A.-M.; Zafiriou, E.; Bargiota, A.; Skaperda, Z.; Kouretas, D.; Roussaki-Schulze, A.-V. Non-Melanoma Skin Cancer and Vitamin D: The “Lost Sunlight” Paradox and the Oxidative Stress Explanation. Antioxidants 2023, 12, 1107. [Google Scholar] [CrossRef] [PubMed]
- Nemazannikova, N.; Antonas, K.; Dass, C.R. Role of Vitamin D Metabolism in Cutaneous Tumour Formation and Progression. J. Pharm. Pharmacol. 2013, 65, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Mah, J.; Amirkia, V. Alkaloids Used as Medicines: Structural Phytochemistry Meets Biodiversity—An Update and Forward Look. Molecules 2021, 26, 1836. [Google Scholar] [CrossRef]
- Castaneda, C.; Bravo, K.; Cortes, N.; Bedoya, J.; de Borges, W.S.; Bastida, J.; Osorio, E. Amaryllidaceae Alkaloids in Skin Cancer Management: Photoprotective Effect on Human Keratinocytes and Anti-Proliferative Activity in Melanoma Cells. J. Appl. Biomed. 2023, 21, 36–47. [Google Scholar] [CrossRef]
- Pal, H.C.; Katiyar, S.K. Cryptolepine, a Plant Alkaloid, Inhibits the Growth of Non-Melanoma Skin Cancer Cells through Inhibition of Topoisomerase and Induction of DNA Damage. Molecules 2016, 21, 1758. [Google Scholar] [CrossRef]
- Heinrich, M.; Barnes, J.; Prieto-Garcia, J.; Gibbons, S.; Williamson, E.M. Fundamentals of Pharmacognosy and Phytotherapy, 3rd ed.; Churchill Livingstone: Edinburgh, UK, 2018; Volume 1. [Google Scholar]
- Min, F.; Liu, X.; Li, Y.; Dong, M.; Qu, Y.; Liu, W. Carnosic Acid Suppresses the Development of Oral Squamous Cell Carcinoma via Mitochondrial-Mediated Apoptosis. Front. Oncol. 2021, 11, 760861. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hamid, I.S.; Mouselhy, Y.Y.; El-Shafei, M.M.; Sayed, M.M.; Mohamed, A.F.; Farag, D.B. COX-2 Docking Structural Analysis with Phytochemical Extracts of Rosemary: A Possible Cytotoxicity on Head and Neck Squamous Cell Carcinoma Cell Line (HEp-2). Anticancer Agents Med. Chem. 2019, 19, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Li, L.; Wu, G. Induction of Ferroptosis by Carnosic Acid-Mediated Inactivation of Nrf2/HO-1 Potentiates Cisplatin Responsiveness in OSCC Cells. Mol. Cell. Probes 2022, 64, 101821. [Google Scholar] [CrossRef] [PubMed]
- Rosen, R.H.; Gupta, A.K.; Tyring, S.K. Dual Mechanism of Action of Ingenol Mebutate Gel for Topical Treatment of Actinic Keratoses: Rapid Lesion Necrosis Followed by Lesion-Specific Immune Response. J. Am. Acad. Dermatol. 2012, 66, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Ogbourne, S.M.; Suhrbier, A.; Jones, B.; Cozzi, S.J.; Boyle, G.M.; Morris, M.; McAlpine, D.; Johns, J.; Scott, T.M.; Sutherland, K.P.; et al. Antitumor Activity of 3-Ingenyl Angelate: Plasma Membrane and Mitochondrial Disruption and Necrotic Cell Death. Cancer Res. 2004, 64, 2833–2839. [Google Scholar] [CrossRef]
- Challacombe, J.M.; Suhrbier, A.; Parsons, P.G.; Jones, B.; Hampson, P.; Kavanagh, D.; Rainger, G.E.; Morris, M.; Lord, J.M.; Le, T.T.T.; et al. Neutrophils Are a Key Component of the Antitumor Efficacy of Topical Chemotherapy with Ingenol-3-Angelate. J. Immunol. 2006, 177, 8123–8132. [Google Scholar] [CrossRef]
- Picato|European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/picato (accessed on 4 December 2023).
- Picato|European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/referrals/picato (accessed on 4 December 2023).
- Drug Safety Update Latest Advice for Medicines Users the Monthly Newsletter from the MHRA and Its Independent Advisor the Commission on Human Medicines. August 2020, Volume 14. Available online: https://assets.publishing.service.gov.uk/media/5f465ff3d3bf7f3d651d8b89/Aug-2020-DSU-PDF.pdf (accessed on 4 December 2023).
- Drug Safety Update Latest Advice for Medicines Users. October 2019, Volume 13. Available online: https://assets.publishing.service.gov.uk/media/5da9bde040f0b6598ec848fd/Oct-2019-PDF.pdf (accessed on 4 December 2023).
- Picato® (Ingenol Mebutate) Gel Product Withdrawal in Canada Due to Potential Increased Risk of Non-Melanoma Skin Cancer|LEO Pharma. Available online: https://www.leo-pharma.ca/Media-Centre/News/News-2019/Picato%C2%AE-(ingenol-mebutate)-gel-Product-Withdrawal-in-Canada-Due-to-Potential-Increased-Risk-of-Non-Melanoma-Skin-Cancer.aspx (accessed on 4 December 2023).
- Juin, S.K.; Ghosh, S.; Majumdar, S. Glycyrrhizic Acid Facilitates Anti-Tumor Immunity by Attenuating Tregs and MDSCs: An Immunotherapeutic Approach. Int. Immunopharmacol. 2020, 88, 106932. [Google Scholar] [CrossRef]
- Umar, S.A.; Tanveer, M.A.; Nazir, L.A.; Divya, G.; Vishwakarma, R.A.; Tasduq, S.A. Glycyrrhizic Acid Prevents Oxidative Stress Mediated DNA Damage Response through Modulation of Autophagy in Ultraviolet-B-Irradiated Human Primary Dermal Fibroblasts. Cell. Physiol. Biochem. 2019, 53, 242–257. [Google Scholar] [CrossRef]
- Adepoju, F.O.; Duru, K.C.; Li, E.; Kovaleva, E.G.; Tsurkan, M.V. Pharmacological Potential of Betulin as a Multitarget Compound. Biomolecules 2023, 13, 1105. [Google Scholar] [CrossRef]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A.; Kurzepa, J. Betulin and Betulinic Acid: Triterpenoids Derivatives with a Powerful Biological Potential. Phytochem. Rev. 2019, 18, 929–951. [Google Scholar] [CrossRef]
- Micali, G.; Lacarrubba, F.; Nasca, M.R.; Schwartz, R.A. Topical Pharmacotherapy for Skin Cancer: Part I. Pharmacology. J. Am. Acad. Dermatol. 2014, 70, 965.e1–965.e12. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, K.; Hu, F.; Chen, L.; Zhang, X.; Wang, F.; Yan, B. Protective Effects of Natural Compounds against Oxidative Stress in Ischemic Diseases and Cancers via Activating the Nrf2 Signaling Pathway: A Mini Review. J. Biochem. Mol. Toxicol. 2021, 35, 22658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.M.; Xu, H.G.; Wang, L.; Li, Y.J.; Sun, P.H.; Wu, X.M.; Wang, G.J.; Chen, W.M.; Ye, W.C. Betulinic Acid and Its Derivatives as Potential Antitumor Agents. Med. Res. Rev. 2015, 35, 1127–1155. [Google Scholar] [CrossRef] [PubMed]
- Drag-Zalesińska, M.; Wysocka, T.; Borska, S.; Drag, M.; Poreba, M.; Choromańska, A.; Kulbacka, J.; Saczko, J. The New Esters Derivatives of Betulin and Betulinic Acid in Epidermoid Squamous Carcinoma Treatment—In Vitro Studies. Biomed. Pharmacother. 2015, 72, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Dehelean, C.A.; Feflea, S.; Gheorgheosu, D.; Ganta, S.; Cimpean, A.M.; Muntean, D.; Amiji, M.M. Anti-Angiogenic and Anti-Cancer Evaluation of Betulin Nanoemulsion in Chicken Chorioallantoic Membrane and Skin Carcinoma in Balb/c Mice. J. Biomed. Nanotechnol. 2013, 9, 577–589. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Yuan, J.M.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, 1700965. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Preetha, R.; Haque, S.; Akhter, N.; Khan, S.; Ahmad, S.; Hussain, A. Dietary Isothiocyanates Inhibit Cancer Progression by Modulation of Epigenome. Semin. Cancer Biol. 2022, 83, 353–376. [Google Scholar] [CrossRef]
- Varadarajan, S.; Madapusi, B.T.; Narasimhan, M.; Pandian, C.D.; Dhanapal, S. Anticancer Effects of Carica Papaya L. and Benzyl Isothiocyanate on an Oral Squamous Cell Carcinoma Cell Line: An In Vitro Study. J. Contemp. Dent. Pract. 2022, 23, 839–844. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Yang, C.Y.; Peng, B.; Ho, S.L.; Tsao, C.H.; Lin, C.K.; Lin, C.S.; Lin, G.J.; Lin, H.Y.; Huang, H.C.; et al. Allyl Isothiocyanate Suppresses the Proliferation in Oral Squamous Cell Carcinoma via Mediating the KDM8/CCNA1 Axis. Biomedicines 2023, 11, 2669. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Yang, Y.; Lu, Y.; Chen, Y.; Luo, X.; Xin, D.; Meng, X.; Shan, Z.; Jiang, G.; Wang, F. Reactivation of Mutant P53 in Esophageal Squamous Cell Carcinoma by Isothiocyanate Inhibits Tumor Growth. Front. Pharmacol. 2023, 14, 1141420. [Google Scholar] [CrossRef]
- Vermeulen, M.; Boerboom, A.M.M.J.F.; Blankvoort, B.M.G.; Aarts, J.M.M.J.G.; Rietjens, I.M.C.M.; van Bladeren, P.J.; Vaes, W.H.J. Potency of Isothiocyanates to Induce Luciferase Reporter Gene Expression via the Electrophile-Responsive Element from Murine Glutathione S-Transferase Ya. Toxicol. In Vitro 2009, 23, 617–621. [Google Scholar] [CrossRef]
- Shirasugi, I.; Kamada, M.; Matsui, T.; Sakakibara, Y.; Liu, M.C.; Suiko, M. Sulforaphane Inhibited Melanin Synthesis by Regulating Tyrosinase Gene Expression in B16 Mouse Melanoma Cells. Biosci. Biotechnol. Biochem. 2010, 74, 579–582. [Google Scholar] [CrossRef]
- Pawlik, A.; Wiczk, A.; Kaczyńska, A.; Antosiewicz, J.; Herman-Antosiewicz, A. Sulforaphane Inhibits Growth of Phenotypically Different Breast Cancer Cells. Eur. J. Nutr. 2013, 52, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Sung, B.; Kang, Y.J.; Hwang, S.Y.; Kim, M.J.; Yoon, J.H.; Im, E.; Kim, N.D. Sulforaphane Inhibits Hypoxia-Induced HIF-1α and VEGF Expression and Migration of Human Colon Cancer Cells. Int. J. Oncol. 2015, 47, 2226–2232. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.X.; Zou, Y.J.; Zhuang, X.B.; Chen, S.X.; Lin, Y.; Li, W.L.; Lin, J.J.; Lin, Z.Q. Sulforaphane Suppresses EMT and Metastasis in Human Lung Cancer through MiR-616-5p-Mediated GSK3β/β-Catenin Signaling Pathways. Acta Pharmacol. Sin. 2017, 38, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Yuanfeng, W.; Gongnian, X.; Jianwei, M.; Shiwang, L.; Jun, H.; Lehe, M. Dietary Sulforaphane Inhibits Histone Deacetylase Activity in B16 Melanoma Cells. J. Funct. Foods 2015, 18, 182–189. [Google Scholar] [CrossRef]
- Yuqing Yang, A.; Kim, H.; Li, W.; Tony Kong, A.-N. Natural Compound-Derived Epigenetic Regulators Targeting Epigenetic Readers, Writers and Erasers. Curr. Top. Med. Chem. 2016, 16, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Shukla, S.; Khan, S.; Tollefsbol, T.O.; Meeran, S.M. Epigenetic Reactivation of P21CIP1/WAF1 and KLOTHO by a Combination of Bioactive Dietary Supplements Is Partially ERα-Dependent in ERα-Negative Human Breast Cancer Cells. Mol. Cell. Endocrinol. 2015, 406, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Saha, K.; Hornyak, T.J.; Eckert, R.L. Epigenetic Cancer Prevention Mechanisms in Skin Cancer. AAPS J. 2013, 15, 1064–1071. [Google Scholar] [CrossRef]
- Su, Z.Y.; Zhang, C.; Lee, J.H.; Shu, L.; Wu, T.Y.; Khor, T.O.; Conney, A.H.; Lu, Y.P.; Kong, A.N.T. Requirement and Epigenetics Reprogramming of Nrf2 in Suppression of Tumor Promoter TPA-Induced Mouse Skin Cell Transformation by Sulforaphane. Cancer Prev. Res. 2014, 7, 319–329. [Google Scholar] [CrossRef]
- Abel, E.L.; Boulware, S.; Fields, T.; McIvor, E.; Powell, K.L.; DiGiovanni, J.; Vasquez, K.M.; MacLeod, M.C. Sulforaphane Induces Phase II Detoxication Enzymes in Mouse Skin and Prevents Mutagenesis Induced by a Mustard Gas Analog. Toxicol. Appl. Pharmacol. 2013, 266, 439. [Google Scholar] [CrossRef]
- Saha, K.; Fisher, M.L.; Adhikary, G.; Grun, D.; Eckert, R.L. Sulforaphane Suppresses PRMT5/MEP50 Function in Epidermal Squamous Cell Carcinoma Leading to Reduced Tumor Formation. Carcinogenesis 2017, 38, 827–836. [Google Scholar] [CrossRef]
- Rorke, E.A.; Adhikary, G.; Szmacinski, H.; Lakowicz, J.R.; Weber, D.J.; Godoy-Ruiz, R.; Puranik, P.; Keillor, J.W.; Gates, E.W.J.; Eckert, R.L. Sulforaphane Covalently Interacts with the Transglutaminase 2 Cancer Maintenance Protein to Alter Its Structure and Suppress Its Activity. Mol. Carcinog. 2022, 61, 19–32. [Google Scholar] [CrossRef]
- Kerr, C.; Adhikary, G.; Grun, D.; George, N.; Eckert, R.L. Combination Cisplatin and Sulforaphane Treatment Reduces Proliferation, Invasion and Tumor Formation in Epidermal Squamous Cell Carcinoma. Mol. Carcinog. 2018, 57, 3. [Google Scholar] [CrossRef]
- McAllister, S.D.; Abood, M.E.; Califano, J.; Guzmán, M. Cannabinoid Cancer Biology and Prevention. J. Natl. Cancer Inst. Monogr. 2021, 2021, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Ramer, R.; Wendt, F.; Wittig, F.; Schäfer, M.; Boeckmann, L.; Emmert, S.; Hinz, B. Impact of Cannabinoid Compounds on Skin Cancer. Cancers 2022, 14, 1769. [Google Scholar] [CrossRef] [PubMed]
- Capodice, J.L.; Kaplan, S.A. The Endocannabinoid System, Cannabis, and Cannabidiol: Implications in Urology and Men’s Health. Curr. Urol. 2021, 15, 95. [Google Scholar] [CrossRef]
- Zheng, D.; Bode, A.M.; Zhao, Q.; Cho, Y.Y.; Zhu, F.; Ma, W.Y.; Dong, Z. The Cannabinoid Receptors Are Required for UV-Induced Inflammation and Skin Cancer Development. Cancer Res. 2008, 68, 3992. [Google Scholar] [CrossRef]
- Gegotek, A.; Biernacki, M.; Ambrozewicz, E.; Surazyński, A.; Wroński, A.; Skrzydlewska, E. The Cross-Talk between Electrophiles, Antioxidant Defence and the Endocannabinoid System in Fibroblasts and Keratinocytes after UVA and UVB Irradiation. J. Dermatol. Sci. 2016, 81, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Soliman, E.; Henderson, K.L.; Danell, A.S.; Van Dross, R. Arachidonoyl-Ethanolamide Activates Endoplasmic Reticulum Stress-Apoptosis in Tumorigenic Keratinocytes: Role of Cyclooxygenase-2 and Novel J-Series Prostamides. Mol. Carcinog. 2016, 55, 117–130. [Google Scholar] [CrossRef]
- Soliman, E.; Van Dross, R. Anandamide-Induced Endoplasmic Reticulum Stress and Apoptosis Are Mediated by Oxidative Stress in Non-Melanoma Skin Cancer: Receptor-Independent Endocannabinoid Signaling. Mol. Carcinog. 2016, 55, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Hyeraci, M.; Papanikolau, E.S.; Grimaldi, M.; Ricci, F.; Pallotta, S.; Monetta, R.; Minafò, Y.A.; Di Lella, G.; Galdo, G.; Abeni, D.; et al. Systemic Photoprotection in Melanoma and Non-Melanoma Skin Cancer. Biomolecules 2023, 13, 1067. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Lokesh, V.; Shang, X.; Shin, J.; Keum, Y.S.; Lee, J.H. Carotenoids: Dietary Sources, Extraction, Encapsulation, Bioavailability, and Health Benefits—A Review of Recent Advancements. Antioxidants 2022, 11, 795. [Google Scholar] [CrossRef]
- Khalaf, R.A.; Awad, M. Lycopene as a Potential Bioactive Compound: Chemistry, Extraction, and Anticancer Prospective. Curr. Cancer Drug Targets 2023, 23, 634–642. [Google Scholar] [CrossRef]
- Amorim, A.D.G.N.; Vasconcelos, A.G.; Souza, J.; Oliveira, A.; Gullón, B.; de Souza de Almeida Leite, J.R.; Pintado, M. Bio-Availability, Anticancer Potential, and Chemical Data of Lycopene: An Overview and Technological Prospecting. Antioxidants 2022, 11, 360. [Google Scholar] [CrossRef]
- Balić, A.; Mokos, M. Do We Utilize Our Knowledge of the Skin Protective Effects of Carotenoids Enough? Antioxidants 2019, 8, 259. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, Y.Y.; Wang, X.; Shen, P.; Jia, Q.; Yu, S.; Wang, Y.; Li, X.; Chen, W.; Wang, A.; et al. Lycopene Prevents Carcinogen-Induced Cutaneous Tumor by Enhancing Activation of the Nrf2 Pathway through P62-Triggered Autophagic Keap1 Degradation. Aging 2020, 12, 8167. [Google Scholar] [CrossRef]
- Brotosudarmo, T.H.P.; Limantara, L.; Setiyono, E. Heriyanto Structures of Astaxanthin and Their Consequences for Therapeutic Application. Int. J. Food Sci. 2020, 2020, 2156582. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, Y.; Zhang, Y.; Zhang, S.; Qu, J.; Wang, X.; Kong, R.; Han, C.; Liu, Z. Fucoxanthin: A Promising Medicinal and Nutritional Ingredient. Evid. Based Complement. Alternat. Med. 2015, 2015, 723515. [Google Scholar] [CrossRef]
- Catanzaro, E.; Bishayee, A.; Fimognari, C. On a Beam of Light: Photoprotective Activities of the Marine Carotenoids Astaxanthin and Fucoxanthin in Suppression of Inflammation and Cancer. Mar. Drugs 2020, 18, 544. [Google Scholar] [CrossRef]
- Matsui, M.; Tanaka, K.; Higashiguchi, N.; Okawa, H.; Yamada, Y.; Tanaka, K.; Taira, S.; Aoyama, T.; Takanishi, M.; Natsume, C.; et al. Protective and Therapeutic Effects of Fucoxanthin against Sunburn Caused by UV Irradiation. J. Pharmacol. Sci. 2016, 132, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, I.; Cao, M.; Su, Z.-y.; Wu, R.; Guo, Y.; Fang, M.; Kong, A.N. Fucoxanthin Elicits Epigenetic Modifications, Nrf2 Activation and Blocking Transformation in Mouse Skin JB6 P+ Cells. AAPS J. 2018, 20, 32. [Google Scholar] [CrossRef]
- Zheng, J.; Piao, M.J.; Kim, K.C.; Yao, C.W.; Cha, J.W.; Hyun, J.W. Fucoxanthin Enhances the Level of Reduced Glutathione via the Nrf2-Mediated Pathway in Human Keratinocytes. Mar. Drugs 2014, 12, 4214. [Google Scholar] [CrossRef]
- Gde Putra Wiraguna, A.A.; Pangkahila, W.; Mantik Astawa, I.N. Antioxidant Properties of Topical Caulerpa Sp. Extract on UVB-Induced Photoaging in Mice. Dermatol. Rep. 2018, 10, 20–25. [Google Scholar] [CrossRef]
- Yoshihisa, Y.; ur Rehman, M.; Shimizu, T. Astaxanthin, a Xanthophyll Carotenoid, Inhibits Ultraviolet-Induced Apoptosis in Keratinocytes. Exp. Dermatol. 2014, 23, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.R.; Sindhuja, H.N.; Dharmesh, S.M.; Sankar, K.U.; Sarada, R.; Ravishankar, G.A. Effective Inhibition of Skin Cancer, Tyrosinase, and Antioxidative Properties by Astaxanthin and Astaxanthin Esters from the Green Alga Haematococcus Pluvialis. J. Agric. Food Chem. 2013, 61, 3842–3851. [Google Scholar] [CrossRef]
- Coderch, L.; López, O.; De La Maza, A.; Parra, J.L. Ceramides and Skin Function. Am. J. Clin. Dermatol. 2012, 4, 107–129. [Google Scholar] [CrossRef]
- Morad, S.A.F.; Cabot, M.C. Ceramide-Orchestrated Signalling in Cancer Cells. Nat. Rev. Cancer 2012, 13, 51–65. [Google Scholar] [CrossRef]
- Alessenko, A.V.; Albi, E. Exploring Sphingolipid Implications in Neurodegeneration. Front. Neurol. 2020, 11, 513554. [Google Scholar] [CrossRef]
- Reyland, M.E. Protein Kinase C Isoforms: Multi-Functional Regulators of Cell Life and Death. Front. Biosci. 2009, 14, 2386–2399. [Google Scholar] [CrossRef]
- Ghosh, S.; Juin, S.K.; Nandi, P.; Majumdar, S.B.; Bose, A.; Baral, R.; Sil, P.C.; Majumdar, S. PKCζ Mediated Anti-Proliferative Effect of C2 Ceramide on Neutralization of the Tumor Microenvironment and Melanoma Regression. Cancer Immunol. Immunother. 2020, 69, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Sugiki, H.; Hozumi, Y.; Maeshima, H.; Katagata, Y.; Mitsuhashi, Y.; Kondo, S. C2-ceramide Induces Apoptosis in a Human Squamous Cell Carcinoma Cell Line. Br. J. Dermatol. 2000, 143, 1154–1163. [Google Scholar] [CrossRef]
- Ebeling, S.; Naumann, K.; Pollok, S.; Wardecki, T.; Vidal-y-Sy, S.; Nascimento, J.M.; Boerries, M.; Schmidt, G.; Brandner, J.M.; Merfort, I. From a Traditional Medicinal Plant to a Rational Drug: Understanding the Clinically Proven Wound Healing Efficacy of Birch Bark Extract. PLoS ONE 2014, 9, e86147. [Google Scholar] [CrossRef] [PubMed]
- Pârvănescu, R.D.; Watz, C.G.; Moacă, E.A.; Vlaia, L.; Marcovici, I.; Macașoi, I.G.; Borcan, F.; Olariu, I.; Coneac, G.; Drăghici, G.A.; et al. Oleogel Formulations for the Topical Delivery of Betulin and Lupeol in Skin Injuries—Preparation, Physicochemical Characterization, and Pharmaco-Toxicological Evaluation. Molecules 2021, 26, 4174. [Google Scholar] [CrossRef] [PubMed]
- Berce, C.; Muresan, M.S.; Soritau, O.; Petrushev, B.; Tefas, L.; Rigo, I.; Ungureanu, G.; Catoi, C.; Irimie, A.; Tomuleasa, C. Cutaneous Wound Healing Using Polymeric Surgical Dressings Based on Chitosan, Sodium Hyaluronate and Resveratrol. A Preclinical Experimental Study. Colloids Surf. B Biointerfaces 2018, 163, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, R.; Campbell, J.; Ukani, G.; O’Reilly Beringhs, A.; Selvaraju, V.; Thirunavukkarasu, M.; Lu, X.; Palesty, J.A.; Maulik, N. Evaluation of Dermal Tissue Regeneration Using Resveratrol Loaded Fibrous Matrix in a Preclinical Mouse Model of Full-Thickness Ischemic Wound. Int. J. Pharm. 2019, 558, 177–186. [Google Scholar] [CrossRef]
- (5) (PDF) Evaluation of Efficacy and Tolerance of a Nighttime Topical Antioxidant Containing Resveratrol, Baicalin, and Vitamin E for Treatment of Mild to Moderately Photodamaged Skin. Available online: https://www.researchgate.net/publication/271538214_Evaluation_of_Efficacy_and_Tolerance_of_a_Nighttime_Topical_Antioxidant_Containing_Resveratrol_Baicalin_and_Vitamin_E_for_Treatment_of_Mild_to_Moderately_Photodamaged_Skin (accessed on 6 November 2023).
- Zulkefli, N.; Che Zahari, C.N.M.; Sayuti, N.H.; Kamarudin, A.A.; Saad, N.; Hamezah, H.S.; Bunawan, H.; Baharum, S.N.; Mediani, A.; Ahmed, Q.U.; et al. Flavonoids as Potential Wound-Healing Molecules: Emphasis on Pathways Perspective. Int. J. Mol. Sci. 2023, 24, 4607. [Google Scholar] [CrossRef]
- Polcz, M.E.; Barbul, A. The Role of Vitamin A in Wound Healing. Nutr. Clin. Pract. 2019, 34, 695–700. [Google Scholar] [CrossRef]
- Hobson, R. Vitamin E and Wound Healing: An Evidence-based Review. Int. Wound J. 2016, 13, 331. [Google Scholar] [CrossRef]
- Kumari, A.; Raina, N.; Wahi, A.; Goh, K.W.; Sharma, P.; Nagpal, R.; Jain, A.; Ming, L.C.; Gupta, M. Wound-Healing Effects of Curcumin and Its Nanoformulations: A Comprehensive Review. Pharmaceutics 2022, 14, 2288. [Google Scholar] [CrossRef] [PubMed]
- Jeya Rajkumar, R.S.; Nadar MSA, M.; Mosae Selvakumar, P. Phytochemicals as a Potential Source for Anti-Microbial, Anti-Oxidant and Wound Healing—A Review. MOJ Bioorg. Org. Chem. 2018, 2, 61–70. [Google Scholar] [CrossRef]
- Hemati, H.; Haghiralsadat, F.; Hemati, M.; Sargazi, G.; Razi, N. Design and Evaluation of Liposomal Sulforaphane-Loaded Polyvinyl Alcohol/Polyethylene Glycol (PVA/PEG) Hydrogels as a Novel Drug Delivery System for Wound Healing. Gels 2023, 9, 748. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Cell Carcinoma (BCC) | Squamous Cell Carcinoma (SCC) | Actinic Keratosis (AK) | |
|---|---|---|---|
| Appearance | Various clinical types, with the nodular type as the most frequently observed variant, usually shiny, pearly papule with a smooth surface, rolled borders, telangiectatic surface vessels [6,30] | Firm, smooth, or hyperkeratotic papule or plaque, possible central ulceration [30] | Macules, papules, or hyperkeratotic plaques with an erythematous background [31] |
| Metastatic potential | Less than 0.1% [5] | From 0.3% to 3.7% [5] | Pre-malignant lesion [32] |
| Site of development | Sun-exposed areas of the body, including the scalp, face, and forehead [6] | Sun-exposed areas of the body, de novo or from a predisposing lesion, including AK [11] | Sun-exposed areas of the body, including the face, neck, dorsum of the hands, forearms, and lower legs [32] |
| Survival rate | About 100% [33] | 95% [33] | About 100% [34] |
| Mutation statues | Hh pathway genes, particularly PTCH and SMO [8] | TP53, p16INK4a genes, PTEN [12,13,18] | TP53, p16INK4a genes, MYC, EGFR genes [26,27,28] |
| Compound | Origin | Study | Conditions | Prevention /Treatment |
|---|---|---|---|---|
| Curcumin | Curcuma longa | Curcumin treatment in HaCaT cells significantly attenuated acute UVB-induced damage by reducing lactate dehydrogenase release, intracellular ROS, and DNA damage while upregulating phase II detoxifying enzymes and promoting DNA repair activity. Topical curcumin application inhibited UVB-induced inflammation, collagen disruption, and lipid peroxidation while promoting Nrf2 nuclear accumulation in hairless mice skin [81]. | In vitro (HaCaT cells) and in vivo (uncovered hairless mice) | Prevention |
| Curcumin-loaded nanopatterned films | Curcuma longa | The curcumin-loaded nanopatterned films presented good cytotoxicity against the SCC cell model [88]. | In vitro (A431 cell line) | Treatment |
| Curcuminoid complex | Curcuma longa | Pre-treatment application of the curcuminoid complex hindered UVB-induced FGF-2 expression, FGF-2-mediated cell proliferation, progression, and colony formation, along with suppressing mTORC1 and mTORC2 activation, as well as FGFR2 phosphorylation, in JB6 epithelial cells. Oral application of the curcuminoid complex on mice inhibited UVB-induced epidermal hyperplasia and hyperproliferation [83]. | In vitro (murine epidermal JB6 P+ cells) and in vivo (SKH-1 mice) | Prevention |
| Curcumin-loaded liposome-siRNA | Curcuma longa | The topical application of a curcumin-loaded liposome-siRNA complex induced a more pronounced inhibition of SCC cell model growth and apoptosis events compared with the control group [87]. | In vitro (A431 cells) | Treatment |
| Gallic acid | Gallnuts, sumac, witch hazel, tea leaves, oak bark | In BCC, gallic acid reduces cell migration and proliferation. In SCC, gallic acid reduces cell migration and proliferation, promotes cell death and significantly lowers HSP90AB1 levels in cancer cells [92]. | In vitro (frozen samples of BCC, SCC from the Human Biological Bank) | Treatment |
| Silymarin | Silybum marianum | Silymarin induces an anti-inflammatory response, protects against thymine dimer formation induced by UVB radiation, triggers apoptosis in damaged cells, encourages the repairment of DNA, and targets aberrant signaling pathways [96]. | In vitro (various types of cell lines, including A431 and HaCaT cells) | prevention |
| Resveratrol + Ursolic Acid | Plant Resveratrol: grapes Ursolic Acid: Arctostaphylos uva ursi | The administration of ursolic acid + resveratrol during skin tumor promotion caused by TPA showed greater tumor development inhibition than using each agent alone [109]. | In vivo (Hsd: ICR (CD-1) mice) | Prevention |
| Quercetin | Onions, grapes, berries, cherries, broccoli, and citrus fruits | Quercetin reduced UVB-induced skin edema and blocked the NF-Κβ/COX-2 signaling pathway [116]. | In vitro (HaCaT cells) | Prevention |
| Kaempferol | Broccoli, cabbage, kale, beans, endives, leek, tomatoes, strawberries, and grapes | Kaempferol delayed tumor growth by targeting RSK2 and MSK1 [119]. | Animal model (SKH-1 hairless mice) | Prevention |
| Epigallocatechin gallate (EGCG) | Green tea | EGCG exhibits anti-proliferation potential by inactivating β-catenin signaling and reduces targets of β-catenin signaling, including MMPs, c-Myc, and VEGF. It decreases the levels of COX-2 and PGE2 as well [121]. | In vitro (A431 and SCC13 cells) | Prevention |
| Gallocatechin gallate (GCG) | Green tea (epimer of EGCG) | GCG protects skin from UVB-induced photodamage, improves skin elasticity, and increases the number of collagen fibers, as well as inhibits aberrations in mitochondria and the formation of melanosomes [123]. | Animal model (BALB/c hairless mice) | Prevention |
| Apigenin | Parsley, chamomile, celery, vine spinach, artichokes, and oregano | Apigenin suppresses the expression of IKKα epithelial-mesenchymal transition (EMT) was diminished, indicating potential anti-metastatic activity [127]. | In vitro (PDVC57 and PB cells) | Prevention |
| Apigenin may induce apoptosis by downregulating sulfiredoxin expression and activating the MAPK signaling pathway 2022 [125]. | In vitro (JB6 cells) | Prevention | ||
| Daidzein | Soybeans | Secondary daidzein metabolite reduced the overexpression of COX-2 and MMP-9 by downregulating MAPK pathway signaling in particulate matter-exposed HaCaT keratinocytes [135]. | In vitro (HaCaT cells) | Prevention |
| Genistein | Soybeans | Genistein suppresses UVB-induced inflammatory cytokines CXCL1, IL-1, MIF, and PLANH1 in vivo. Topical-administrated genistein decreased the number of skin folds and wrinkles induced by UVB in animal models. Diet reach in genistein in human participants significantly reduced the severity of UVB-induced wrinkling [136]. | In vitro (HaCaT cells), in vivo (Sprague–Dawley rats, humans) | Prevention |
| Genistein has an anti-nitrosative effect, preventing UVB-induced cell damage [137]. | In vivo (hairless HRS/J mice) | Prevention | ||
| Biochanin A | Red clover, chickpeas, soybeans | Biochanin A inhibited the expression of UV-induced COX-2 [139]. | In vitro (HaCaT cells, JB6 P+ mouse skin) | Prevention |
| Vitamin A | Eggs, dairy products, meat organs, fish | Vitamin A represses overexpressed activator protein-1, inhibits tumor angiogenesis, and diminishes activation of STAT3 [36]. | In vitro (human BCC cell line—BCC-1/KMC) and in vivo (Ptch1+/− mice; humans) | Prevention, treatment |
| Vitamin C | Brussels sprouts, bell peppers, and berries, such as strawberries and blackcurrants | Vitamin C has antioxidant properties, influences DNA repair and replication, and produces hydrogen peroxide in extracellular fluid [146]. | In vitro (Colo-16 cells) | Prevention |
| Vitamin E | Plant nuts, plant-based oils, soybeans, wheat germ | Vitamin E prevents the peroxidation of membrane lipids and reduces UVB-induced damage [144]. | In vivo (C3H/HeN mice; humans) | Prevention |
| Vitamin D | Fish, mushrooms | Vitamin D regulates cancer cell proliferation, apoptosis, differentiation, and angiogenesis [142]. | In vivo (humans) | prevention |
| Cryptolepine | Cryptolepis sanguinolenta | Cryptolepine expands phosphorilation of ATM/ATR, BRCA1, Chk1/Chk2, and γH2AX, activates the p53 signaling pathway, and promotes apoptosis. It also has an anti-proliferative effect and downregulates cyclin-dependent kinases, cyclin A, cyclin E, cyclin A, cyclin E kinases, and cyclin D1 [155]. | In vitro (SCC-13, A431, HaCaT, NHEK cells) | Treatment |
| Lycoramine Tazettine | Amaryllidaceae | Lycoramine and tazettine, which protected human keratinocytes from UVB-induced production of ROS and IL-6 [154]. | In vitro (HaCaT cells) | Prevention |
| Glycyrrhizic acid | Glycyrrhiza glabra | Glycyrrhizic acid provides protection to skin cells against UVB radiation, a key contributor to NMSC. It reduces cell death and DNA damage, exhibits antioxidant properties, affects autophagy processes, downregulates DNA damage marker proteins, and stabilizes the AKT/PTEN axis disrupted by UVB radiation. Its protective effects are influenced by autophagy regulators [169]. | In vitro (human dermal fibroblasts) | Prevention |
| Betulin, Betulinic Acid, Betulin esters | Betula pendula | Newly modified betulin-originated esters have significant therapeutic potential for actinic keratosis [175]. | In vitro (HaCaT cells) | Treatment |
| Betulin reduces skin lesions and irritation, notably decreasing erythema, and inhibits the initiation and promotion of skin tumors [176]. | In vivo (Balb/c mice) | Prevention | ||
| Sulforaphane | Broccoli, broccoli sprouts | SFN reactivates Nrf2, a transcription factor for antioxidant enzymes, by downregulating DNA methyltransferases (DNMTs) and HDACs in JB6 mouse skin epidermal cells exposed to TPA, thus suppressing TPA-induced malignant transformation [191]. | In vitro (JB6 P+) | Prevention |
| Through the involvement of the Nrf2-dependent mechanism, topical application of SFN on mouse skin results in increased glutathione (GSH) and glutathione S-transferase 4 (GST4) synthesis, which inhibits skin mutagenesis [192]. | In vivo (C57BL/6 and Big Blue mice) | Prevention | ||
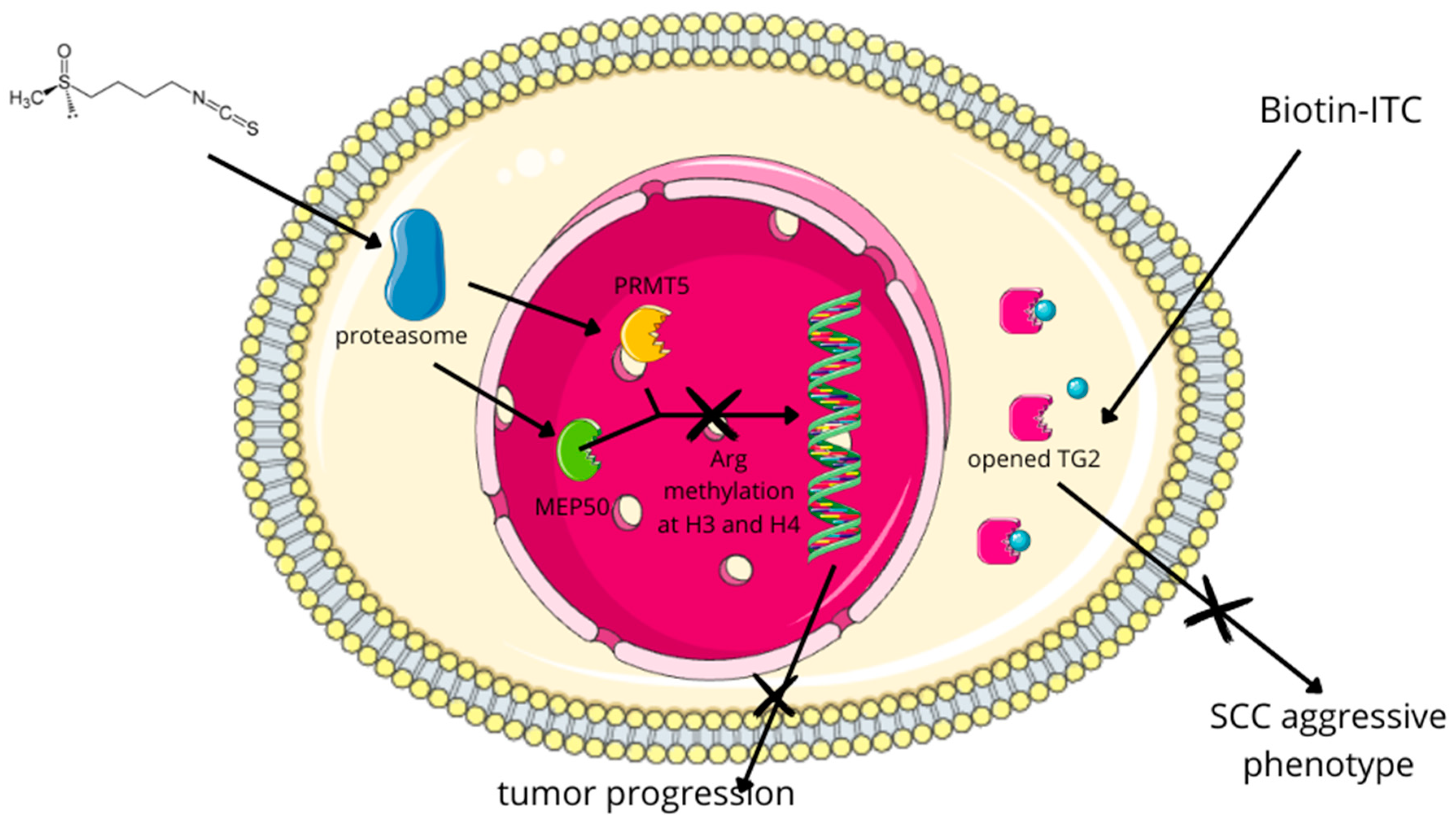
| The administration of SFN diminishes the levels of PRMT5 and MEP50, thereby causing the formation of H4R3me2s. This phenomenon correlates with decreased cellular proliferation, invasion, and migration of SCC [193]. | In vitro (SCC-13, A431, HaCaT cells) | Treatment | ||
| Biotin-tagged SFN analog (Biotin-ITC) | Broccoli, broccoli sprouts | The application of a biotin-tagged SFN analog (Biotin-ITC) showed that SFN, through the inhibition of TG2, partially inhibits its binding to GTP, which is crucial for maintaining the aggressive SCC phenotype [194]. | In vitro (SCC-13, HaCaT cells) | Treatment |
| Sulforaphane and cisplatin | Broccoli, broccoli sprouts | Combined therapy of SFN and cisplatin for SCC occurred to suppress tumor formation and reduce the population of cancer stem cells within the tumor [195]. | In vitro (SCC-13 cells) | Treatment |
| AEA | Endogenous | AEA induces ER stress-induced apoptosis in NMSC cells through a receptor-independent mechanism mediated by oxidative stress [202]. | In vitro (murine squamous carcinoma cell line JWF2) | Treatment |
| AEA is selectively toxic in tumor cells that overexpress COX-2 [201] | In vitro (murine squamous carcinoma cell line JWF2) | Treatment | ||
| Lycopene | Tomatoes | Lycopene reduces both the incidence and multiplicity of cutaneous tumors, as well as inhibiting the tumorigenesis of normal cutaneous cells during the promotion phase [208]. | In vitro (JB6 P+ cells) | Prevention |
| Astaxanthin (ASX) Fucoxanthin (FX) | ASX: Haematococcus pluvialis FX: Phaeophyceae | FX and ASX inhibit the TPA-induced transformation of mouse skin JB6 P+ cells [213]. | In vitro (murine skin JB6 P+ cells) | Prevention |
| Astaxanthin (ASX) | Haematococcus pluvialis | Prior exposure of ASX in human keratinocytes before UVB exposure can inhibit DNA damage [216]. | In vitro (HaCaT cells) | Prevention |
| The topical application of ASX in mice can protect against UVB-induced DNA damage [215]. | In vivo (Wistar mice) | Prevention | ||
| Astaxanthin Monesters (AXME) and Diesters (AXDE) | Haematococcus pluvialis | AXME and AXDE exhibit a more significant reduction in DMBA-induced tumor incidences compared with ASX alone [217] | In vivo (albino Wistar rats model) | Prevention |
| C2 Ceramide | Endogenous | C2 ceramide induces apoptosis in human SCC cells (HSC-I)—it is confirmed by dose-dependent toxicity and typical morphological changes in intrinsic apoptosis, such as chromatin condensation, internucleosomal DNA fragmentation, and nuclear fragmentation [223]. | In vitro (HSC-I) | Treatment |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalski, S.; Karska, J.; Tota, M.; Skinderowicz, K.; Kulbacka, J.; Drąg-Zalesińska, M. Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment. Molecules 2024, 29, 728. https://doi.org/10.3390/molecules29030728
Kowalski S, Karska J, Tota M, Skinderowicz K, Kulbacka J, Drąg-Zalesińska M. Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment. Molecules. 2024; 29(3):728. https://doi.org/10.3390/molecules29030728
Chicago/Turabian StyleKowalski, Szymon, Julia Karska, Maciej Tota, Katarzyna Skinderowicz, Julita Kulbacka, and Małgorzata Drąg-Zalesińska. 2024. "Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment" Molecules 29, no. 3: 728. https://doi.org/10.3390/molecules29030728
APA StyleKowalski, S., Karska, J., Tota, M., Skinderowicz, K., Kulbacka, J., & Drąg-Zalesińska, M. (2024). Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment. Molecules, 29(3), 728. https://doi.org/10.3390/molecules29030728