
MCC950 Ameliorates Diabetic Muscle Atrophy in Mice by Inhibition of Pyroptosis and Its Synergistic Effect with Aerobic Exercise
 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
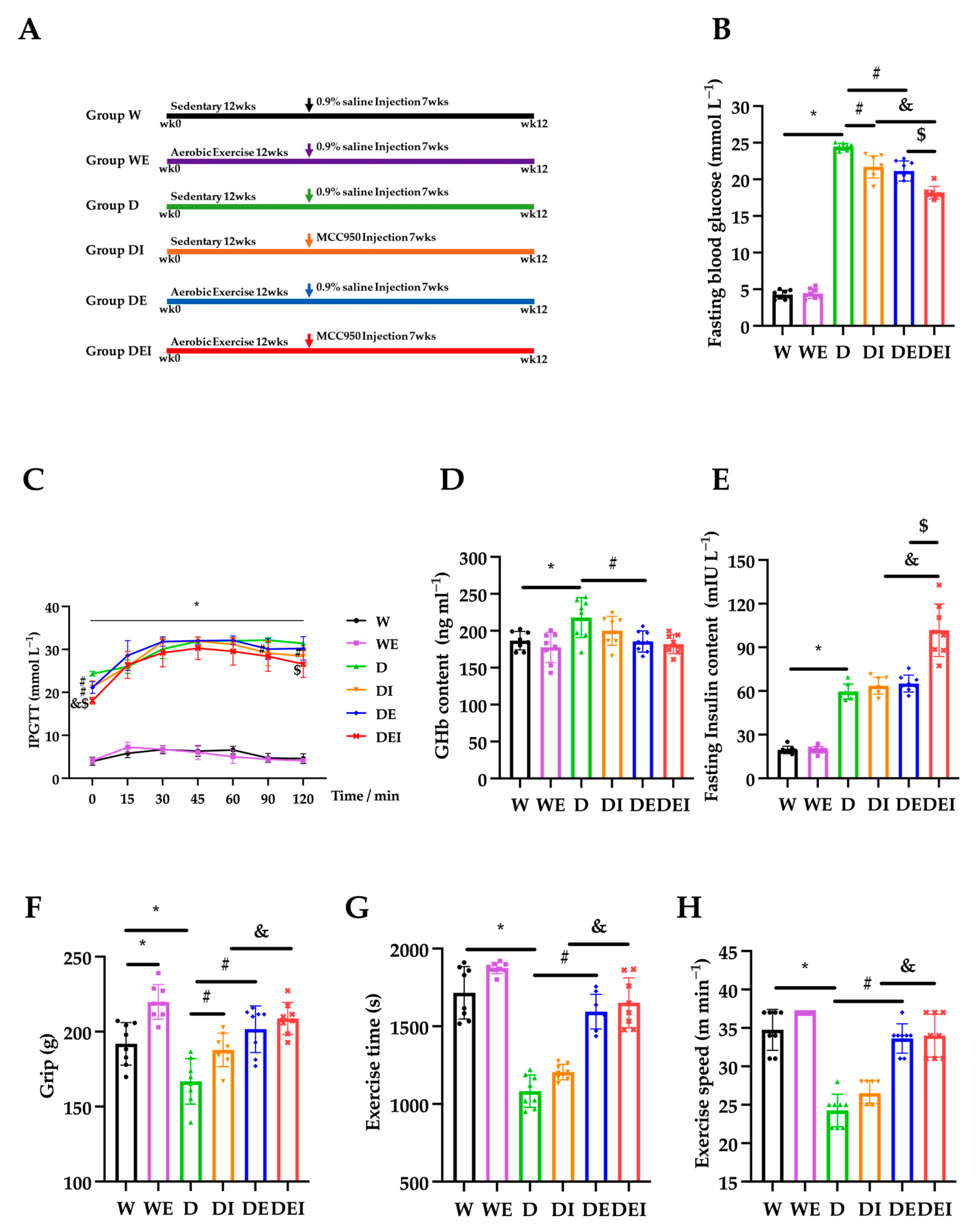
2.1. MCC950 Treatment and Aerobic Exercise Improves Diabetes Glucose Metabolism Levels and Muscle Function
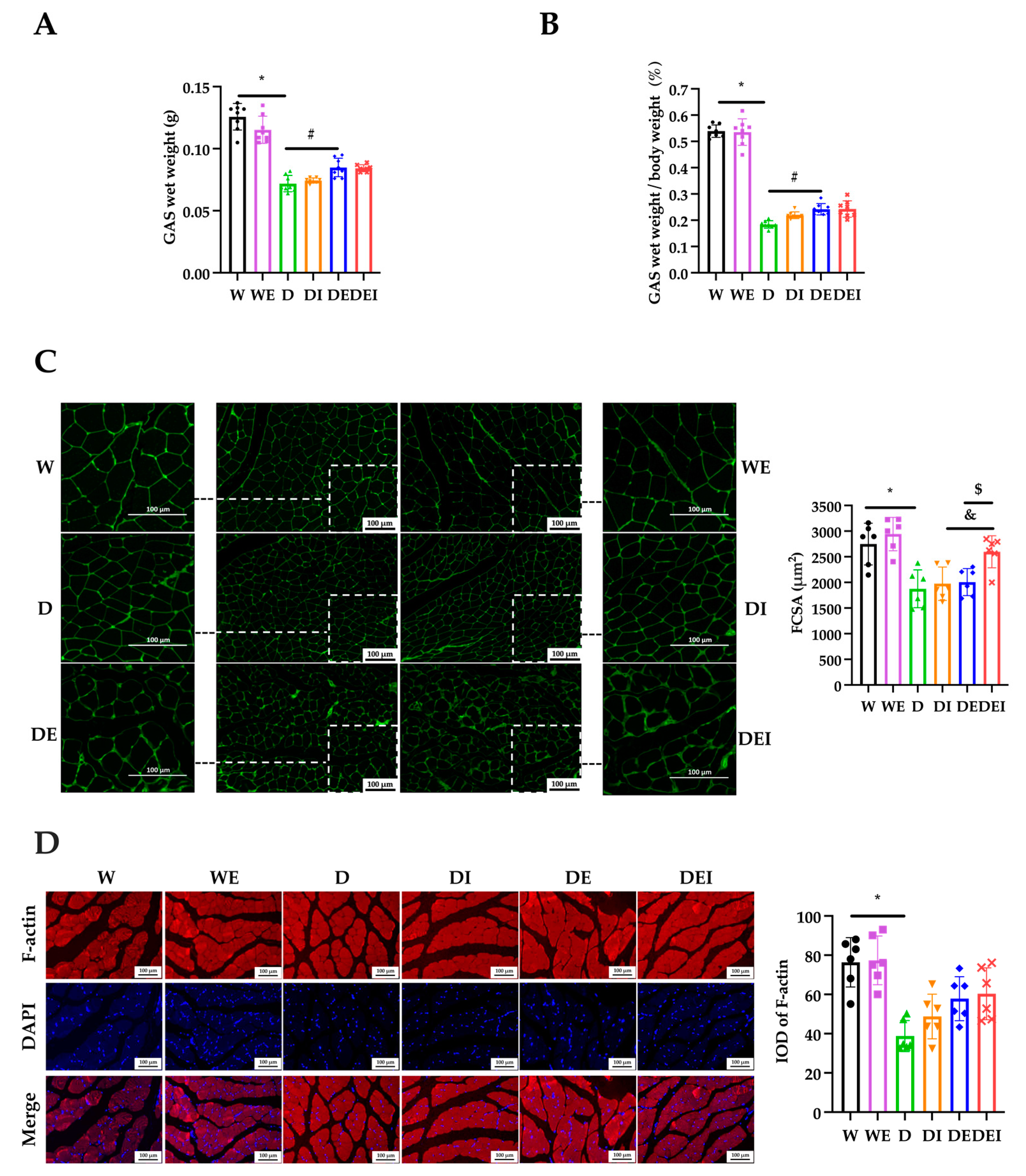
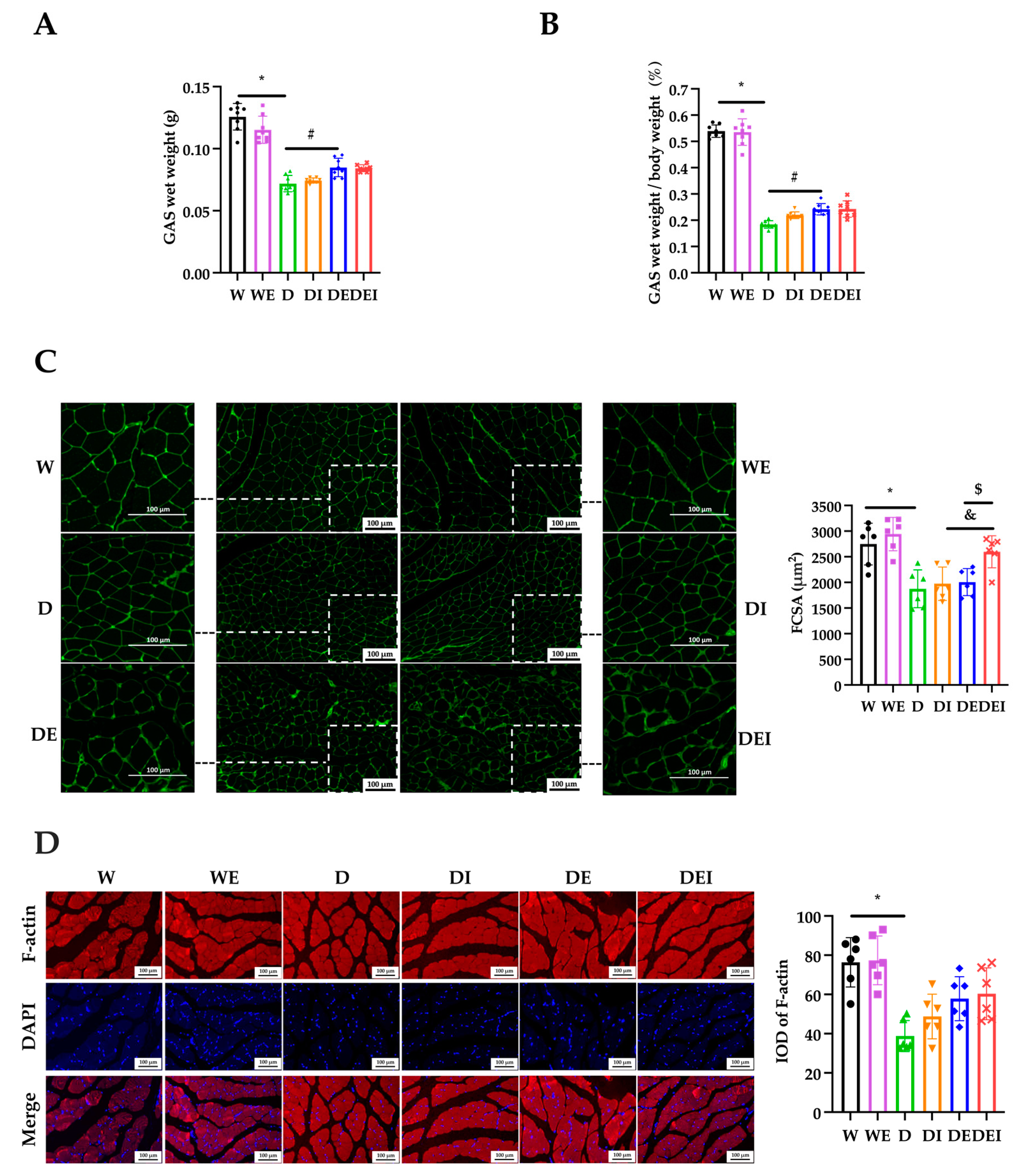
2.2. MCC950 Treatment and Aerobic Exercise Alleviates the Degree of Muscle Atrophy
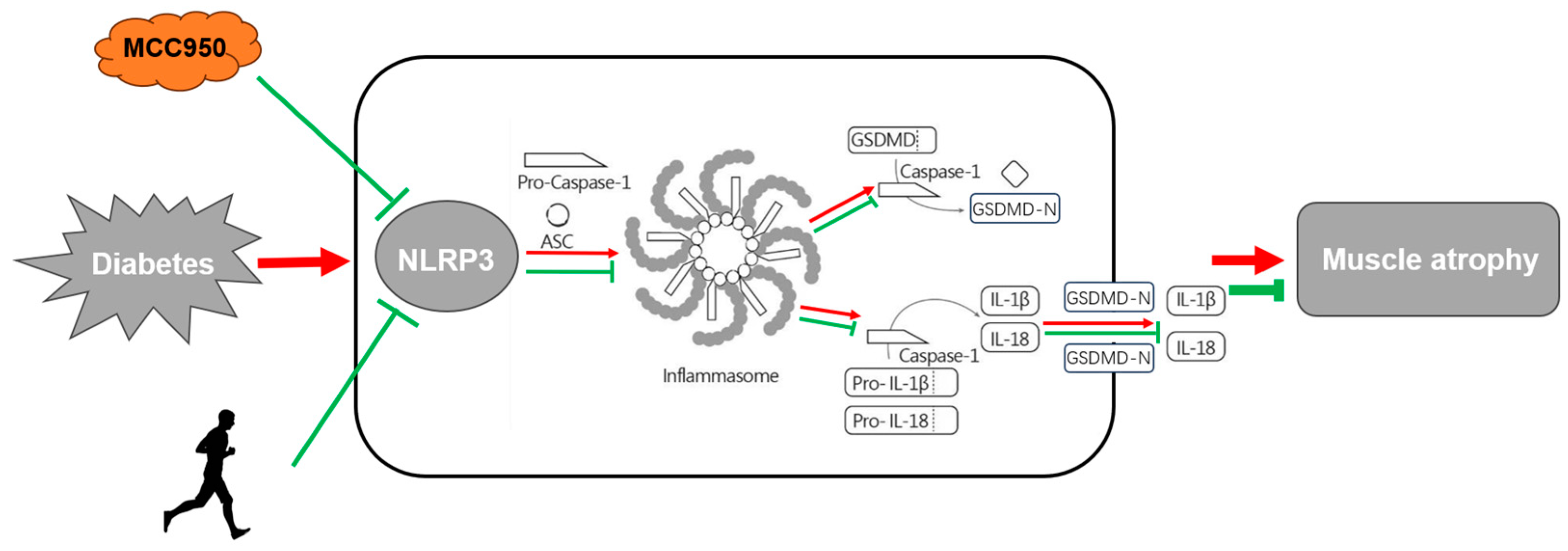
2.3. MCC950 Treatment and Aerobic Exercise Moderates the Levels of Apoptosis and Pyroptosis
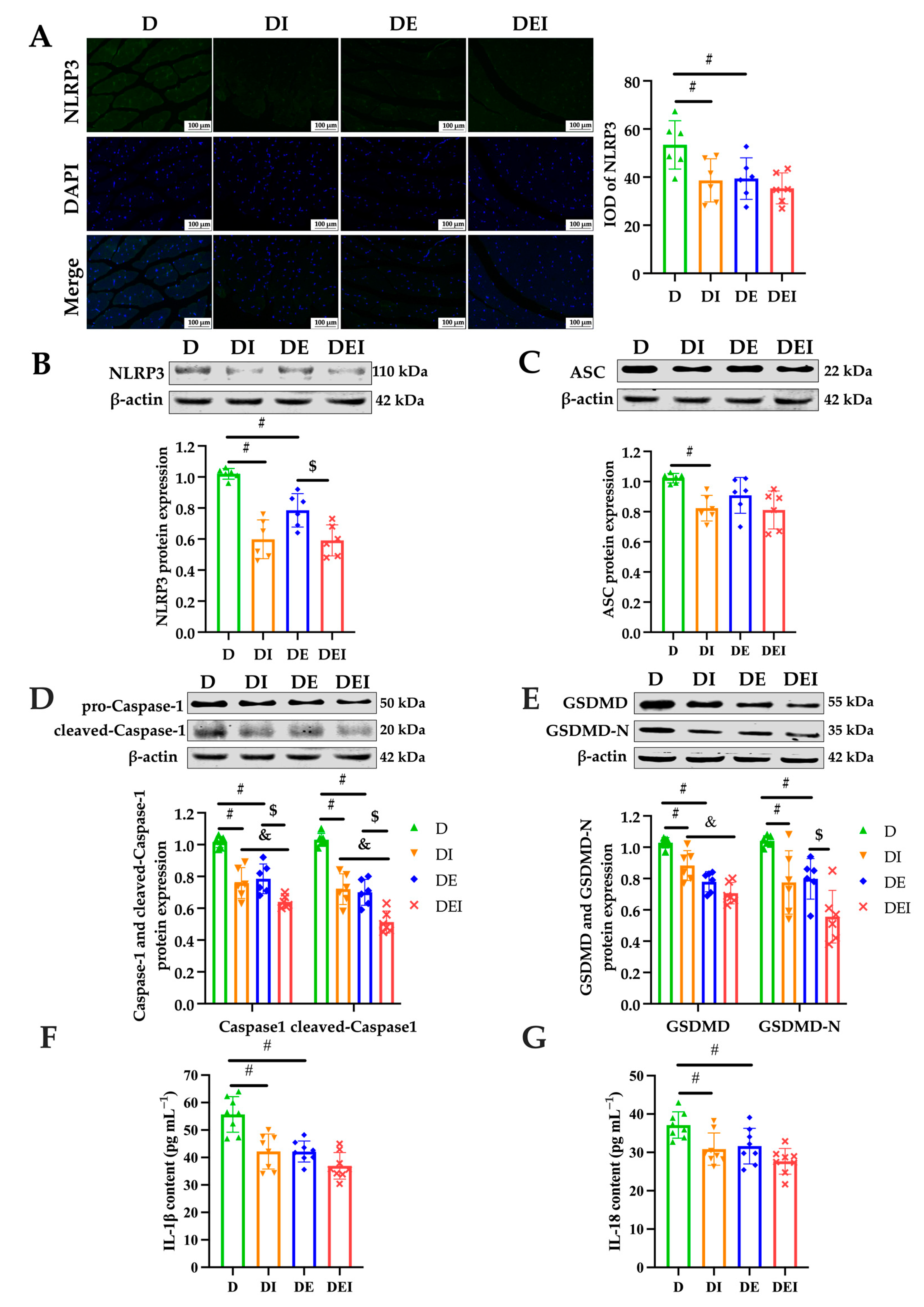
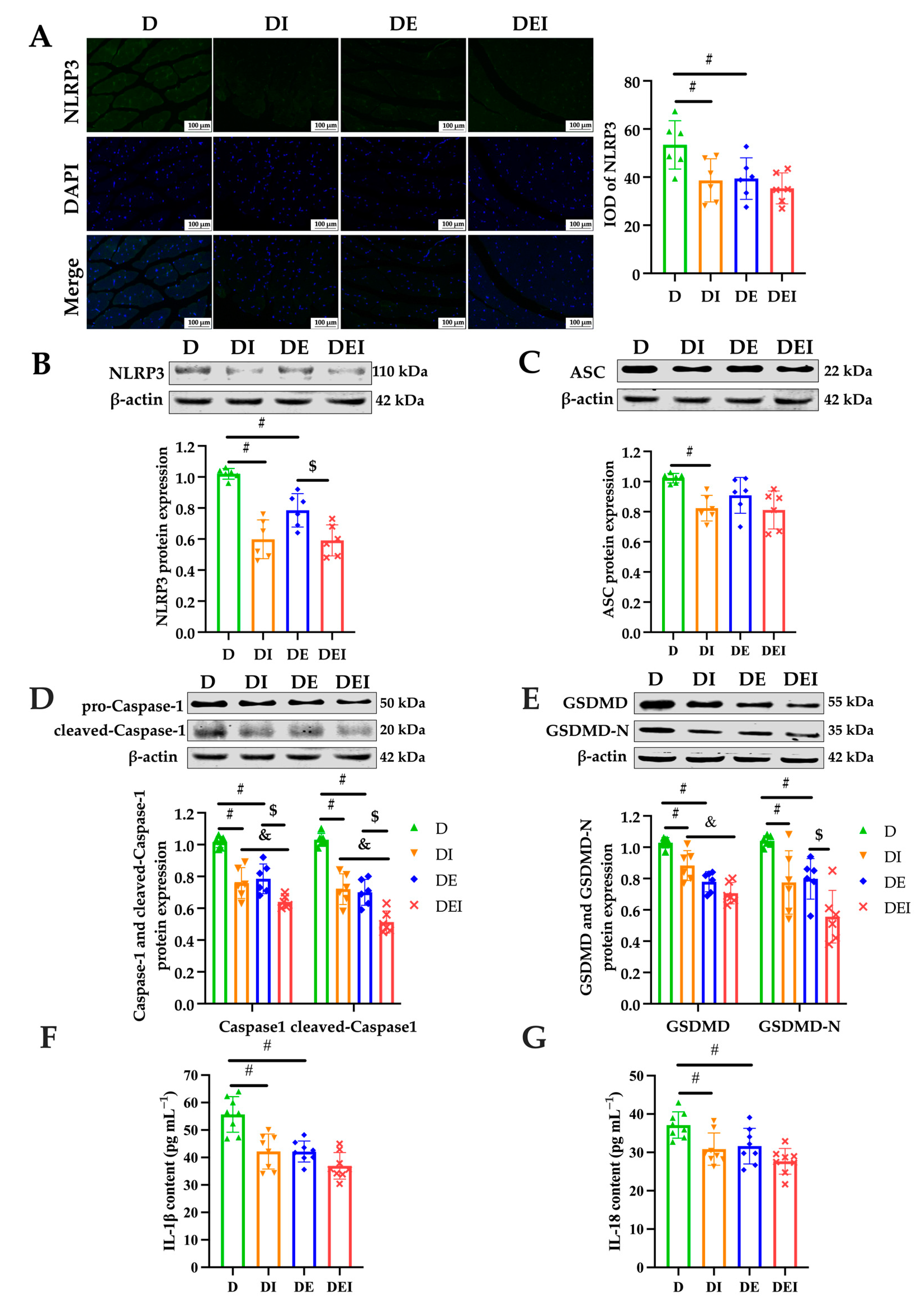
2.4. MCC950 Treatment and Aerobic Exercise Downregulates the Expression of Pyroptosis-Related Proteins
3. Materials and Methods
3.1. Animals
3.2. Mice Training Protocol
3.3. Intraperitoneal Glucose Tolerance Tests (IPGTT)
3.4. Muscle Sample Collection
3.5. Serum Glucose Profiles
3.6. Serum Glycosylated Hemoglobin (GHb), Insulin and Inflammatory Markers Content
3.7. Exercise Performance Test
3.8. Grip Strength Test
3.9. Immunofluorescence (IF) Staining
3.10. TdT-Mediated dUTP-Biotin Nick End Labeling (TUNEL) Staining
3.11. Transmission Electron Micrographs (TEM)
3.12. Western Blot
3.13. Statistical Analyses
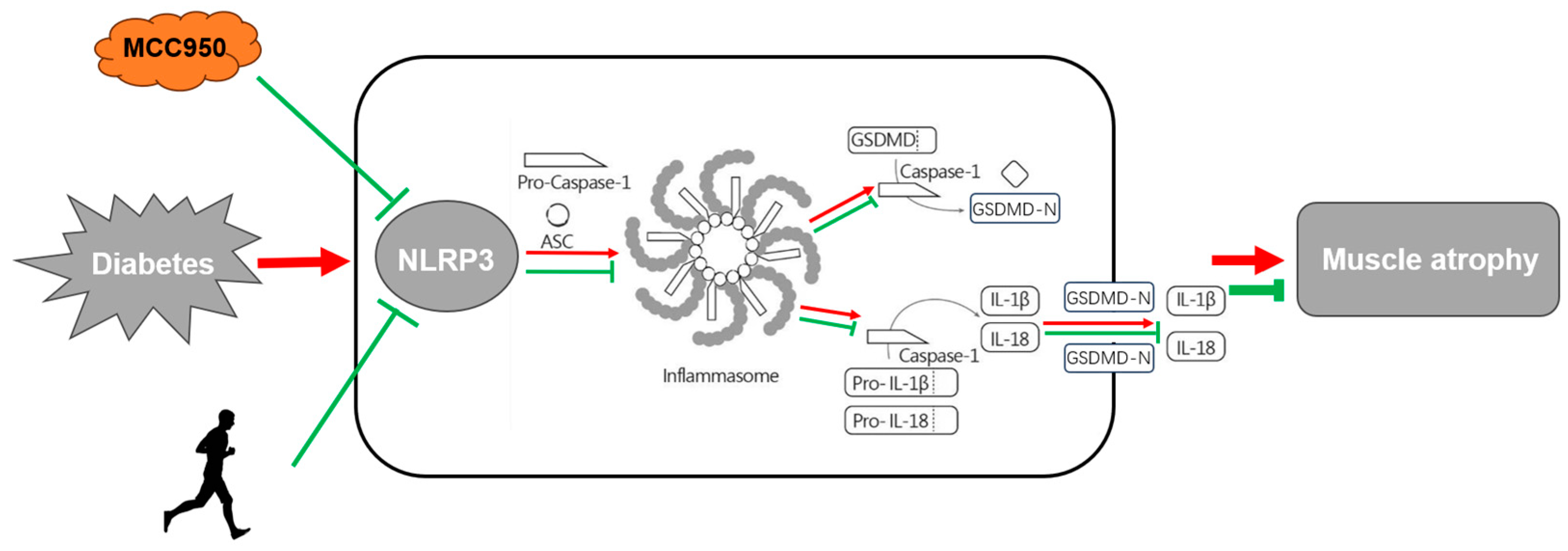
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, Y.; Li, M.; Wang, K.; Qi, G.; Liu, H.; Wang, W.; Ji, Y.; Chang, M.; Deng, C.; Xu, F.; et al. Diabetic muscular atrophy: Molecular mechanisms and promising therapies. Front. Endocrinol. 2022, 13, 917113. [Google Scholar] [CrossRef] [PubMed]
- Trierweiler, H.; Kisielewicz, G.; Hoffmann Jonasson, T.; Rasmussen Petterle, R.; Aguiar Moreira, C.; Zeghbi Cochenski Borba, V. Sarcopenia: A chronic complication of type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2018, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Nellaiappan, K.; Preeti, K.; Khatri, D.K.; Singh, S.B. Diabetic complications: An update on pathobiology and therapeutic strategies. Curr. Diabetes Rev. 2022, 18, e030821192146. [Google Scholar] [CrossRef] [PubMed]
- Kotake, H.; Ogura, Y.; Yamada, S.; Inoue, K.; Watanabe, S.; Ichikawa, D.; Sugaya, T.; Ohata, K.; Natsuki, Y.; Hoshino, S.; et al. Mechanism for exercise-mediated prevention against muscle wasting on extensor digitorum longus muscle in Spontaneously Diabetic Torii fatty rats. J. Physiol. Sci. JPS 2023, 73, 5. [Google Scholar] [CrossRef] [PubMed]
- Balducci, S.; Zanuso, S.; Nicolucci, A.; Fernando, F.; Cavallo, S.; Cardelli, P.; Fallucca, S.; Alessi, E.; Letizia, C.; Jimenez, A.; et al. Anti-inflammatory effect of exercise training in subjects with type 2 diabetes and the metabolic syndrome is dependent on exercise modalities and independent of weight loss. Nutr. Metab. Cardiovasc. Dis. NMCD 2010, 20, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Chang, S.J. Moderate exercise suppresses NF-κB signaling and activates the SIRT1-AMPK-PGC1α Axis to attenuate muscle loss in diabetic db/db mice. Front. Physiol. 2018, 9, 636. [Google Scholar] [CrossRef]
- Sriwijitkamol, A.; Christ-Roberts, C.; Berria, R.; Eagan, P.; Pratipanawatr, T.; DeFronzo, R.A.; Mandarino, L.J.; Musi, N. Reduced skeletal muscle inhibitor of kappaB beta content is associated with insulin resistance in subjects with type 2 diabetes: Reversal by exercise training. Diabetes 2006, 55, 760–767. [Google Scholar] [CrossRef]
- Gordon, P.L.; Vannier, E.; Hamada, K.; Layne, J.; Hurley, B.F.; Roubenoff, R.; Castaneda-Sceppa, C. Resistance training alters cytokine gene expression in skeletal muscle of adults with type 2 diabetes. Int. J. Immunopathol. Pharmacol. 2006, 19, 739–749. [Google Scholar] [CrossRef]
- Bai, Y.; Mu, Q.; Bao, X.; Zuo, J.; Fang, X.; Hua, J.; Zhang, D.; Jiang, G.; Li, P.; Gao, S.; et al. Targeting NLRP3 inflammasome in the treatment of diabetes and diabetic complications: Role of natural compounds from herbal medicine. Aging Dis. 2021, 12, 1587–1604. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Okon, I.; Liu, Z.; Bedarida, T.; Wang, Q.; Ramprasath, T.; Zhang, M.; Song, P.; Zou, M.H. Ablation of Neuropilin 1 in Myeloid Cells Exacerbates High-Fat Diet-Induced Insulin Resistance through Nlrp3 Inflammasome In Vivo. Diabetes 2017, 66, 2424–2435. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, D.; Li, T.; Yang, F.; Li, Z.; Bai, X.; Wang, Y. The role of NLRP3 inflammasome in inflammation-related skeletal muscle atrophy. Front. Immunol. 2022, 13, 1035709. [Google Scholar] [CrossRef] [PubMed]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Muñoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nat. Med. 2015, 21, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Yang, Z.; Zhang, C.; Shi, Y.; Han, W.; Song, S.; Mu, L.; Du, C.; Shi, Y. Inhibition of NLRP3 inflammasome ameliorates podocyte damage by suppressing lipid accumulation in diabetic nephropathy. Metab. Clin. Exp. 2021, 118, 154748. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Choi, J.S.Y.; Stefanovic, N.; Al-Sharea, A.; Simpson, D.S.; Mukhamedova, N.; Jandeleit-Dahm, K.; Murphy, A.J.; Sviridov, D.; Vince, J.E.; et al. Specific NLRP3 Inhibition Protects Against Diabetes-Associated Atherosclerosis. Diabetes 2021, 70, 772–787. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Meng, X.; Ye, T.; Xie, W.; Sun, G.; Sun, X. Inhibiting the NLRP3 Inflammasome Activation with MCC950 Ameliorates Diabetic Encephalopathy in db/db Mice. Molecules 2018, 23, 522. [Google Scholar] [CrossRef] [PubMed]
- Ge, K.; Wang, Y.; Li, P.; Li, M.; Zhang, W.; Dan, H.; Hu, X.; Zhou, J.; Yang, Q.; Wang, J.; et al. Down-expression of the NLRP3 inflammasome delays the progression of diabetic retinopathy. Microvasc. Res. 2022, 139, 104265. [Google Scholar] [CrossRef] [PubMed]
- Eggelbusch, M.; Shi, A.; Broeksma, B.C.; Vázquez-Cruz, M.; Soares, M.N.; de Wit, G.M.J.; Everts, B.; Jaspers, R.T.; Wüst, R.C.I. The NLRP3 inflammasome contributes to inflammation-induced morphological and metabolic alterations in skeletal muscle. J. Cachexia Sarcopenia Muscle 2022, 13, 3048–3061. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson, N.; Davis-López de Carrizosa, M.A.; Versele, R.; Selvais, C.M.; Noel, L.; Van den Bergh, P.Y.D.; Brichard, S.M.; Abou-Samra, M. Inhibiting the inflammasome with MCC950 counteracts muscle pyroptosis and improves Duchenne muscular dystrophy. Front. Immunol. 2022, 13, 1049076. [Google Scholar] [CrossRef]
- Li, H.; Guan, Y.; Liang, B.; Ding, P.; Hou, X.; Wei, W.; Ma, Y. Therapeutic potential of MCC950, a specific inhibitor of NLRP3 inflammasome. Eur. J. Pharmacol. 2022, 928, 175091. [Google Scholar] [CrossRef]
- Bakhshi, S.; Shamsi, S. MCC950 in the treatment of NLRP3-mediated inflammatory diseases: Latest evidence and therapeutic outcomes. Int. Immunopharmacol. 2022, 106, 108595. [Google Scholar] [CrossRef]
- Javaid, H.M.A.; Sahar, N.E.; ZhuGe, D.L.; Huh, J.Y. Exercise inhibits NLRP3 inflammasome activation in obese mice via the anti-inflammatory effect of meteorin-like. Cells 2021, 10, 3480. [Google Scholar] [CrossRef]
- Chen, H.; Charlat, O.; Tartaglia, L.A.; Woolf, E.A.; Weng, X.; Ellis, S.J.; Lakey, N.D.; Culpepper, J.; Moore, K.J.; Breitbart, R.E.; et al. Evidence that the diabetes gene encodes the leptin receptor: Identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495. [Google Scholar] [CrossRef]
- Yoon, J.W.; Leiter, E.H.; Coleman, D.L.; Kim, M.K.; Pak, C.Y.; McArthur, R.G.; Roncari, D.A. Genetic control of organ-reactive autoantibody production in mice by obesity (ob) diabetes (db) genes. Diabetes 1988, 37, 1287–1293. [Google Scholar] [CrossRef]
- Shao, J.; Yamashita, H.; Qiao, L.; Friedman, J.E. Decreased Akt kinase activity and insulin resistance in C57BL/KsJ-Leprdb/db mice. J. Endocrinol. 2000, 167, 107–115. [Google Scholar] [CrossRef]
- Su, W.; Guo, Z.; Randall, D.C.; Cassis, L.; Brown, D.R.; Gong, M.C. Hypertension and disrupted blood pressure circadian rhythm in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1634–H1641. [Google Scholar] [CrossRef]
- Xiang, M.; Yuan, X.; Zhang, N.; Zhang, L.; Liu, Y.; Liu, J.; Gao, Y.; Xu, Y.; Sun, W.; Tang, Q.; et al. Effects of exercise, metformin, and combination treatments on type 2 diabetic mellitus-induced muscle atrophy in db/db mice: Crosstalk between autophagy and the proteasome. J. Physiol. Biochem. 2023, 80, 235–247. [Google Scholar] [CrossRef]
- Lu, F.; Lu, B.; Zhang, L.; Wen, J.; Wang, M.; Zhang, S.; Li, Q.; Shu, F.; Sun, Y.; Liu, N.; et al. Hydrogen sulphide ameliorating skeletal muscle atrophy in db/db mice via Muscle RING finger 1 S-sulfhydration. J. Cell. Mol. Med. 2020, 24, 9362–9377. [Google Scholar] [CrossRef]
- Bamba, R.; Okamura, T.; Hashimoto, Y.; Majima, S.; Senmaru, T.; Ushigome, E.; Nakanishi, N.; Asano, M.; Yamazaki, M.; Takakuwa, H.; et al. Extracellular lipidome change by an SGLT2 inhibitor, luseogliflozin, contributes to prevent skeletal muscle atrophy in db/db mice. J. Cachexia Sarcopenia Muscle 2022, 13, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Moon, J.W.; Lee, J.O.; Kim, J.H.; Jung, E.J.; Kim, S.J.; Oh, J.Y.; Wu, S.W.; Lee, P.R.; Park, S.H.; et al. Metformin induces muscle atrophy by transcriptional regulation of myostatin via HDAC6 and FoxO3a. J. Cachexia Sarcopenia Muscle 2022, 13, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.L.; McMillin, S.L.; Weyrauch, L.A.; Witczak, C.A. Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training. Nutrients 2019, 11, 2432. [Google Scholar] [CrossRef] [PubMed]
- Alaca, N.; Uslu, S.; Gulec Suyen, G.; Ince, U.; Serteser, M.; Kurtel, H. Effects of different aerobic exercise frequencies on streptozotocin-nicotinamide-induced type 2 diabetic rats: Continuous versus short bouts and weekend warrior exercises. J. Diabetes 2018, 10, 73–84. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Takahashi, F.; Okamura, T.; Hamaguchi, M.; Fukui, M. Diet, exercise, and pharmacotherapy for sarcopenia in people with diabetes. Metab. Clin. Exp. 2023, 144, 155585. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.H.; Fan, C.; Zhang, Q.; Jiang, Y.H.; Li, Y.; Xue, Y.T. MCC950 attenuates doxorubicin-induced myocardial injury in vivo and in vitro by inhibiting NLRP3-mediated pyroptosis. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 143, 112133. [Google Scholar] [CrossRef]
- Américo-Da-Silva, L.; Aguilera, J.; Quinteros-Waltemath, O.; Sánchez-Aguilera, P.; Russell, J.; Cadagan, C.; Meneses-Valdés, R.; Sánchez, G.; Estrada, M.; Jorquera, G.; et al. Activation of the NLRP3 inflammasome increases the IL-1β Level and decreases GLUT4 translocation in skeletal muscle during insulin resistance. Int. J. Mol. Sci. 2021, 22, 10212. [Google Scholar] [CrossRef] [PubMed]
- Nalbandian, A.; Khan, A.A.; Srivastava, R.; Llewellyn, K.J.; Tan, B.; Shukr, N.; Fazli, Y.; Kimonis, V.E.; BenMohamed, L. Activation of the NLRP3 inflammasome is associated with valosin-containing protein myopathy. Inflammation 2017, 40, 21–41. [Google Scholar] [CrossRef]
- Hull, C.; Dekeryte, R.; Buchanan, H.; Kamli-Salino, S.; Robertson, A.; Delibegovic, M.; Platt, B. NLRP3 inflammasome inhibition with MCC950 improves insulin sensitivity and inflammation in a mouse model of frontotemporal dementia. Neuropharmacology 2020, 180, 108305. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Ma, Y.; Wang, X.; Zhang, Y.; Zhu, L.; Shi, S.; Pan, S.; Liu, Z. Advanced glycation end products induce skeletal muscle atrophy and insulin resistance via activating ROS-mediated ER stress PERK/FOXO1 signaling. Am. J. Physiol. Endocrinol. Metab. 2023, 324, E279–E287. [Google Scholar] [CrossRef]
- Yang, X.; Wang, L.; Zhang, L.; Zhai, X.; Sheng, X.; Quan, H.; Lin, H. Exercise mitigates Dapagliflozin-induced skeletal muscle atrophy in STZ-induced diabetic rats. Diabetol. Metab. Syndr. 2023, 15, 154. [Google Scholar] [CrossRef]
- Duan, S.; Jiang, X.; Li, J.; Fu, M.; Li, Z.; Cheng, Y.; Zhuang, Y.; Yang, M.; Xiao, W.; Ping, H.; et al. The RXFP2-PLC/PKC signaling pathway mediates INSL3-induced regulation of the proliferation, migration and apoptosis of mouse gubernacular cells. Cell. Mol. Biol. Lett. 2023, 28, 16. [Google Scholar] [CrossRef]
- Costamagna, D.; Costelli, P.; Sampaolesi, M.; Penna, F. Role of inflammation in muscle homeostasis and myogenesis. Mediat. Inflamm. 2015, 2015, 805172. [Google Scholar] [CrossRef] [PubMed]
- Perry, B.D.; Caldow, M.K.; Brennan-Speranza, T.C.; Sbaraglia, M.; Jerums, G.; Garnham, A.; Wong, C.; Levinger, P.; Asrar Ul Haq, M.; Hare, D.L.; et al. Muscle atrophy in patients with Type 2 Diabetes Mellitus: Roles of inflammatory pathways, physical activity and exercise. Exerc. Immunol. Rev. 2016, 22, 94–109. [Google Scholar]
- Sayed, R.K.; Fernández-Ortiz, M.; Fernández-Martínez, J.; Aranda Martínez, P.; Guerra-Librero, A.; Rodríguez-Santana, C.; de Haro, T.; Escames, G.; Acuña-Castroviejo, D.; Rusanova, I. The impact of melatonin and NLRP3 inflammasome on the expression of microRNAs in aged muscle. Antioxidants 2021, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Chen, W.; Hua, Y.; Jia, H.; Song, Y.; Wang, Y. Aerobic exercise ameliorates cardiac hypertrophy by regulating mitochondrial quality control and endoplasmic reticulum stress through M(2) AChR. J. Cell. Physiol. 2021, 236, 6581–6596. [Google Scholar] [CrossRef]
- Slavin, M.B.; Khemraj, P.; Hood, D.A. Exercise, mitochondrial dysfunction and inflammasomes in skeletal muscle. Biomed. J. 2023, 47, 100636. [Google Scholar] [CrossRef]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Tian, J.; Fan, J.; Liu, X.; Wang, R. Exercise training-attenuated insulin resistance and liver injury in elderly pre-diabetic patients correlates with NLRP3 inflammasome. Front. Immunol. 2023, 14, 1082050. [Google Scholar] [CrossRef] [PubMed]
- Bortolon, J.R.; Murata, G.M.; Borges, L.; Weimann, E.; Silva, M.B.B.; Dermargos, A.; Hatanaka, E. Recovery of diabetic rats after physical exhaustion: Kinetic alterations in muscle inflammation and muscle-signaling proteins to atrophy and hypertrophy. Front. Physiol. 2020, 11, 573416. [Google Scholar] [CrossRef]
- De Paepe, B. Progressive skeletal muscle atrophy in muscular dystrophies: A role for toll-like receptor-signaling in disease pathogenesis. Int. J. Mol. Sci. 2020, 21, 4440. [Google Scholar] [CrossRef]
- Burdette, B.E.; Esparza, A.N.; Zhu, H.; Wang, S. Gasdermin D in pyroptosis. Acta Pharm. Sinica. B 2021, 11, 2768–2782. [Google Scholar] [CrossRef]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef]
- Nabata, A.; Kuroki, M.; Ueba, H.; Hashimoto, S.; Umemoto, T.; Wada, H.; Yasu, T.; Saito, M.; Momomura, S.I.; Kawakami, M. C-reactive protein induces endothelial cell apoptosis and matrix metalloproteinase-9 production in human mononuclear cells: Implications for the destabilization of atherosclerotic plaque. Atherosclerosis 2008, 196, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sun, X.; Wang, C.; He, H. Effects of exercise on inflammatory cytokines in patients with type 2 diabetes: A meta-analysis of randomized controlled trials. Oxidative Med. Cell. Longev. 2020, 2020, 6660557. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Mammucari, C. Regulation of skeletal muscle growth by the IGF1-Akt/PKB pathway: Insights from genetic models. Skelet. Muscle 2011, 1, 4. [Google Scholar] [CrossRef]
- Zhou, H.; Li, X.M.; Meinkoth, J.; Pittman, R.N. Akt regulates cell survival and apoptosis at a postmitochondrial level. J. Cell Biol. 2000, 151, 483–494. [Google Scholar] [CrossRef]
- Feng, L.; Li, B.; Xi, Y.; Cai, M.; Tian, Z. Aerobic exercise and resistance exercise alleviate skeletal muscle atrophy through IGF-1/IGF-1R-PI3K/Akt pathway in mice with myocardial infarction. Am. J. Physiol. Cell Physiol. 2022, 322, C164–C176. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.; Zhang, J.; Du, J.; Zhang, Q.; Ge, Z.; Ding, S. Chrono-Aerobic Exercise Optimizes Metabolic State in DB/DB Mice through CLOCK-Mitophagy-Apoptosis. Int. J. Mol. Sci. 2022, 23, 9308. [Google Scholar] [CrossRef]
- Li, N.; Zhou, H.; Wu, H.; Wu, Q.; Duan, M.; Deng, W.; Tang, Q. STING-IRF3 contributes to lipopolysaccharide-induced cardiac dysfunction, inflammation, apoptosis and pyroptosis by activating NLRP3. Redox Biol. 2019, 24, 101215. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Huang, X.; Xiang, Y.; Dai, J.; Xu, L.; Jiang, J.; Xu, J. Ablation of NLRP3 inflammasome attenuates muscle atrophy via inhibiting pyroptosis, proteolysis and apoptosis following denervation. Theranostics 2023, 13, 374–390. [Google Scholar] [CrossRef]
- Kong, H.; Zhao, H.; Chen, T.; Song, Y.; Cui, Y. Targeted P2X7/NLRP3 signaling pathway against inflammation, apoptosis, and pyroptosis of retinal endothelial cells in diabetic retinopathy. Cell Death Dis. 2022, 13, 336. [Google Scholar] [CrossRef]
- Lin, Q.; Li, S.; Jiang, N.; Jin, H.; Shao, X.; Zhu, X.; Wu, J.; Zhang, M.; Zhang, Z.; Shen, J.; et al. Inhibiting NLRP3 inflammasome attenuates apoptosis in contrast-induced acute kidney injury through the upregulation of HIF1A and BNIP3-mediated mitophagy. Autophagy 2021, 17, 2975–2990. [Google Scholar] [CrossRef]
- Dempsey, C.; Rubio Araiz, A.; Bryson, K.J.; Finucane, O.; Larkin, C.; Mills, E.L.; Robertson, A.A.B.; Cooper, M.A.; O’Neill, L.A.J.; Lynch, M.A. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-β and cognitive function in APP/PS1 mice. Brain Behav. Immun. 2017, 61, 306–316. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, Y.; Ouyang, F.; Su, M.; Li, W.; Chen, J.; Xiao, H.; Zhou, X.; Liu, B. Extracellular vesicles derived from mesenchymal stem cells alleviate neuroinflammation and mechanical allodynia in interstitial cystitis rats by inhibiting NLRP3 inflammasome activation. J. Neuroinflammation 2022, 19, 80. [Google Scholar] [CrossRef]
- Higashikuni, Y.; Liu, W.; Numata, G.; Tanaka, K.; Fukuda, D.; Tanaka, Y.; Hirata, Y.; Imamura, T.; Takimoto, E.; Komuro, I.; et al. NLRP3 inflammasome activation through heart-brain interaction initiates cardiac inflammation and hypertrophy during pressure overload. Circulation 2023, 147, 338–355. [Google Scholar] [CrossRef]
- Gao, R.F.; Li, X.; Xiang, H.Y.; Yang, H.; Lv, C.Y.; Sun, X.L.; Chen, H.Z.; Gao, Y.; Yang, J.S.; Luo, W.; et al. The covalent NLRP3-inflammasome inhibitor Oridonin relieves myocardial infarction induced myocardial fibrosis and cardiac remodeling in mice. Int. Immunopharmacol. 2021, 90, 107133. [Google Scholar] [CrossRef]
- Zeng, W.; Wu, D.; Sun, Y.; Suo, Y.; Yu, Q.; Zeng, M.; Gao, Q.; Yu, B.; Jiang, X.; Wang, Y. The selective NLRP3 inhibitor MCC950 hinders atherosclerosis development by attenuating inflammation and pyroptosis in macrophages. Sci. Rep. 2021, 11, 19305. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, Y.; Dellsperger, K.C.; Zhang, C. Exercise training improves endothelial function via adiponectin-dependent and independent pathways in type 2 diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H306–H314. [Google Scholar] [CrossRef]
- Zhang, B.; Lu, J.; Jiang, Y.; Feng, Y. Asprosin contributes to nonalcoholic fatty liver disease through regulating lipid accumulation and inflammatory response via AMPK signaling. Immun. Inflamm. Dis. 2023, 11, e947. [Google Scholar] [CrossRef] [PubMed]
- Ustunel, I.; Demir, R. A histochemical, morphometric and ultrastructural study of gastrocnemius and soleus muscle fiber type composition in male and female rats. Acta Anat. 1997, 158, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Jiao, Y.; Fan, Q.; Qi, A.; Xiao, L.; Li, J. Homocysteine, interleukin-1β, and fasting blood glucose levels as prognostic markers for diabetes mellitus complicated with cerebral infarction and correlated with carotid intima-media thickness. Exp. Ther. Med. 2020, 19, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Wu, C.H.; Tsai, M.L.; Huang, Y.J.; Chen, S.H. Detection of total and A1c-glycosylated hemoglobin in human whole blood using sandwich immunoassays on polydimethylsiloxane-based antibody microarrays. Anal. Chem. 2012, 84, 8635–8641. [Google Scholar] [CrossRef] [PubMed]
- Alimu, A.; Abudureman, H.; Wang, Y.Z.; Li, M.Y.; Wang, J.S.; Liu, Z.L. Decabromodiphenyl ether causes insulin resistance and glucose and lipid metabolism disorders in mice. World J. Diabetes 2021, 12, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Song, A.Q.; Gao, B.; Fan, J.J.; Zhu, Y.J.; Zhou, J.; Wang, Y.L.; Xu, L.Z.; Wu, W.N. NLRP1 inflammasome contributes to chronic stress-induced depressive-like behaviors in mice. J. Neuroinflammation 2020, 17, 178. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, A.V.; Brícola, R.S.; Braga, R.R.; Lenhare, L.; Silva, V.R.R.; Anaruma, C.P.; Katashima, C.K.; Crisol, B.M.; Simabuco, F.M.; Silva, A.S.R.; et al. Aerobic exercise training induces the mitonuclear imbalance and UPRmt in the skeletal muscle of aged mice. J. Gerontol.. Ser. A Biol. Sci. Med. Sci. 2020, 75, 2258–2261. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chan, M.C.; Yu, Y.; Bei, Y.; Chen, P.; Zhou, Q.; Cheng, L.; Chen, L.; Ziegler, O.; Rowe, G.C.; et al. miR-29b contributes to multiple types of muscle atrophy. Nat. Commun. 2017, 8, 15201. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Z.; Li, X.; Chen, J.; Yu, Z.; Li, X.; Sun, C.; Hu, L.; Wu, M.; Liu, L. Polydatin protects against atherosclerosis by activating autophagy and inhibiting pyroptosis mediated by the NLRP3 inflammasome. J. Ethnopharmacol. 2023, 309, 116304. [Google Scholar] [CrossRef]
- Peng, Y.; Li, J.; Luo, D.; Zhang, S.; Li, S.; Wang, D.; Wang, X.; Zhang, Z.; Wang, X.; Sun, C.; et al. Muscle atrophy induced by overexpression of ALAS2 is related to muscle mitochondrial dysfunction. Skelet. Muscle 2021, 11, 9. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Fu, P.; Zhang, Y.; Ling, D.; Reynolds, L.; Hua, W.; Wang, Z.; Ma, F.; Li, B.; Yu, J.; et al. MCC950 Ameliorates Diabetic Muscle Atrophy in Mice by Inhibition of Pyroptosis and Its Synergistic Effect with Aerobic Exercise. Molecules 2024, 29, 712. https://doi.org/10.3390/molecules29030712
Yan X, Fu P, Zhang Y, Ling D, Reynolds L, Hua W, Wang Z, Ma F, Li B, Yu J, et al. MCC950 Ameliorates Diabetic Muscle Atrophy in Mice by Inhibition of Pyroptosis and Its Synergistic Effect with Aerobic Exercise. Molecules. 2024; 29(3):712. https://doi.org/10.3390/molecules29030712
Chicago/Turabian StyleYan, Xiaoyu, Pengyu Fu, Yimin Zhang, Dongmei Ling, Lewis Reynolds, Weicheng Hua, Zhiyuan Wang, Fangyuan Ma, Boxuan Li, Jingjing Yu, and et al. 2024. "MCC950 Ameliorates Diabetic Muscle Atrophy in Mice by Inhibition of Pyroptosis and Its Synergistic Effect with Aerobic Exercise" Molecules 29, no. 3: 712. https://doi.org/10.3390/molecules29030712
APA StyleYan, X., Fu, P., Zhang, Y., Ling, D., Reynolds, L., Hua, W., Wang, Z., Ma, F., Li, B., Yu, J., Liu, Y., Gong, L., & Zhang, E. (2024). MCC950 Ameliorates Diabetic Muscle Atrophy in Mice by Inhibition of Pyroptosis and Its Synergistic Effect with Aerobic Exercise. Molecules, 29(3), 712. https://doi.org/10.3390/molecules29030712