Abstract
Safflower (Carthamus tinctorius L.) has been recognized for its medicinal value, but there have been limited studies on the glycosyltransferases involved in the biosynthesis of flavonoid glycosides from safflower. In this research, we identified two highly efficient flavonoid O-glycosyltransferases, CtOGT1 and CtOGT2, from safflower performing local BLAST alignment. By constructing a prokaryotic expression vector, we conducted in vitro enzymatic reactions and discovered that these enzymes were capable of catalyzing two-step O-glycosylation using substrates such as kaempferol, quercetin, and eriodictyol. Moreover, they exhibited efficient catalytic activity towards various compounds, including flavones (apigenin, scutellarein), dihydrochalcone (phloretin), isoflavones (genistein, daidzein), flavanones (naringenin, glycyrrhizin), and flavanonols (dihydrokaempferol), leading to the formation of O-glycosides. The broad substrate specificity of these enzymes is noteworthy. This study provides valuable insights into the biosynthetic pathways of flavonoid glycosides in safflower. The discovery of CtOGT1 and CtOGT2 enhances our understanding of the enzymatic processes involved in synthesizing flavonoid glycosides in safflower, contributing to the overall comprehension of secondary metabolite biosynthesis in this plant species.
1. Introduction
Carthamus tinctorius L., commonly referred to as safflower, is an herbaceous plant belonging to the Carthamus genus of the Asteraceae family. It has been cultivated for approximately 4500 years and originated from the “New Crescent” region along the eastern Mediterranean coast [1]. Safflower has a rich history in traditional medicine and is known for its therapeutic properties in treating various conditions such as joint injuries and pain. In modern pharmacology, safflower has shown significant effects in improving myocardial ischemia, exerting anti-inflammatory responses, and preventing thrombotic events [2]. Flavonoids are valuable compounds found in safflower, which can be categorized into two major groups. The first group includes special compounds like hydroxysafflor yellow A (HSYA), currently undergoing phase III clinical trials for the treatment of acute ischemic stroke [3]. Safflower yellow injection, containing HSYA as the main component, has obtained approval from the Chinese National Medical Products Administration (NMPA) for treating angina and coronary heart diseases [4]. The second group comprises common compounds such as flavonoids, flavonols, and dihydroflavonoids. Examples of these compounds include quercetin-3-O-glucoside (known for its antioxidant, anti-inflammatory, and anticancer properties) [5,6], rutin (possessing anticancer, anti-inflammatory, antibacterial, and antidiabetic effects) [7,8,9], naringenin-7-O-glucoside (vasorelaxant, antioxidant, and antidiabetic) [10,11,12], isorhamnetin-3-O-glucoside (antiadipogenic, antioxidant, and anti-inflammatory) [13,14,15], and genistin (anticancer, antioxidant, cardioprotective, anti-apoptotic, neuroprotective, hepatoprotective, and antimicrobial) [16]. Xian et al. summarized a total of 24 special category compounds and 43 common compounds that have been identified in safflower (Table 1) [17,18].

Table 1.
The flavonoid compounds in safflower.
Glycosylation is a common post-translational modification in plant biosynthetic pathways that greatly contributes to the structural diversity of plant secondary metabolites [19,20]. Glycosides, categorized as O-, C-, S-, and N-glycosides based on the type of glycosidic bond, are formed through this process [21]. In the biosynthesis of glycosylated flavonoids, glycosyltransferases play a vital role. These enzymes utilize nucleotide diphosphate-activated sugar moieties such as UDP-glucose, UDP-xylose, UDP-galactose, UDP-arabinose, UDP-glucuronic acid, UDP-N-acetylglucosamine, and others as sugar donors. They facilitate the formation of region- and stereo-specific glycosidic bonds and belong to the UDP-dependent glycosyltransferase (UGT) family [22,23]. Despite the wide range of flavonoid glycosides present in safflower, only a few UGT genes involved in their biosynthesis have been functionally characterized thus far. The complete elucidation of the biosynthetic pathway for major active compounds like HSYA remains incomplete. Ren et al. demonstrated the glycosylation function of candidate UGTs using safflower corolla protoplasts in vivo for the first time. Overexpression of CtUGT3 resulted in a significant increase in the content of kaempferol-3-O-glucoside, indicating its glycosylation activity towards flavonoids with 3-OH and 7-OH moieties [24]. Qi et al. provided preliminary evidence for the function of CtUGT4 as a flavonoid-O-glycosyltransferase in safflower by demonstrating that its overexpression led to a noteworthy increase in the content of quercetin-3-O-rutinoside and a trend of increased quercetin-3-O-glycoside content [25]. Although several O-glycosyltransferases have been identified in safflower, the aforementioned enzymes did not exhibit significant di-O-glycosylation activity.
The application and development of synthetic biology has made significant breakthroughs in achieving the efficient synthesis of active compounds in safflower, which is of great significance for the sustainable utilization of valuable traditional Chinese medicine resources. Here, we reported two highly efficient di-O-glycosyltransferases with broad substrate specificity from the safflower, providing references for the elucidation of the biosynthetic pathways of flavonoids within it.
2. Results
2.1. Screening of Safflower Flavonoid O-Glycosyltransferases
To screen candidate genes encoding O-glycosyltransferases in the safflower transcriptome, considering the promiscuity of glycosyltransferase substrates, tBLASTn analysis was performed using the amino acid sequences of Vitis vinifera’s glycosyltransferase VvGT1 and Arabidopsis thaliana’s glycosyltransferase UGT89C1 as reference sequences (E-value < e−40) [26,27]. Three candidate genes with complete ORFs were selected from the safflower transcriptome screening and named CtOGT1 and CtOGT2. The ExPASy online server was used to predict the number of amino acids, relative molecular weight, theoretical isoelectric point, molecular formula, and instability coefficient encoded by the candidate genes. The results are shown in Table S1. The three-dimensional protein structure prediction of CtOGT1 and CtOGT2 is shown in Figure S1.
2.2. Phylogenetic and Multiple Sequence Alignment Analysis of CtOGTs
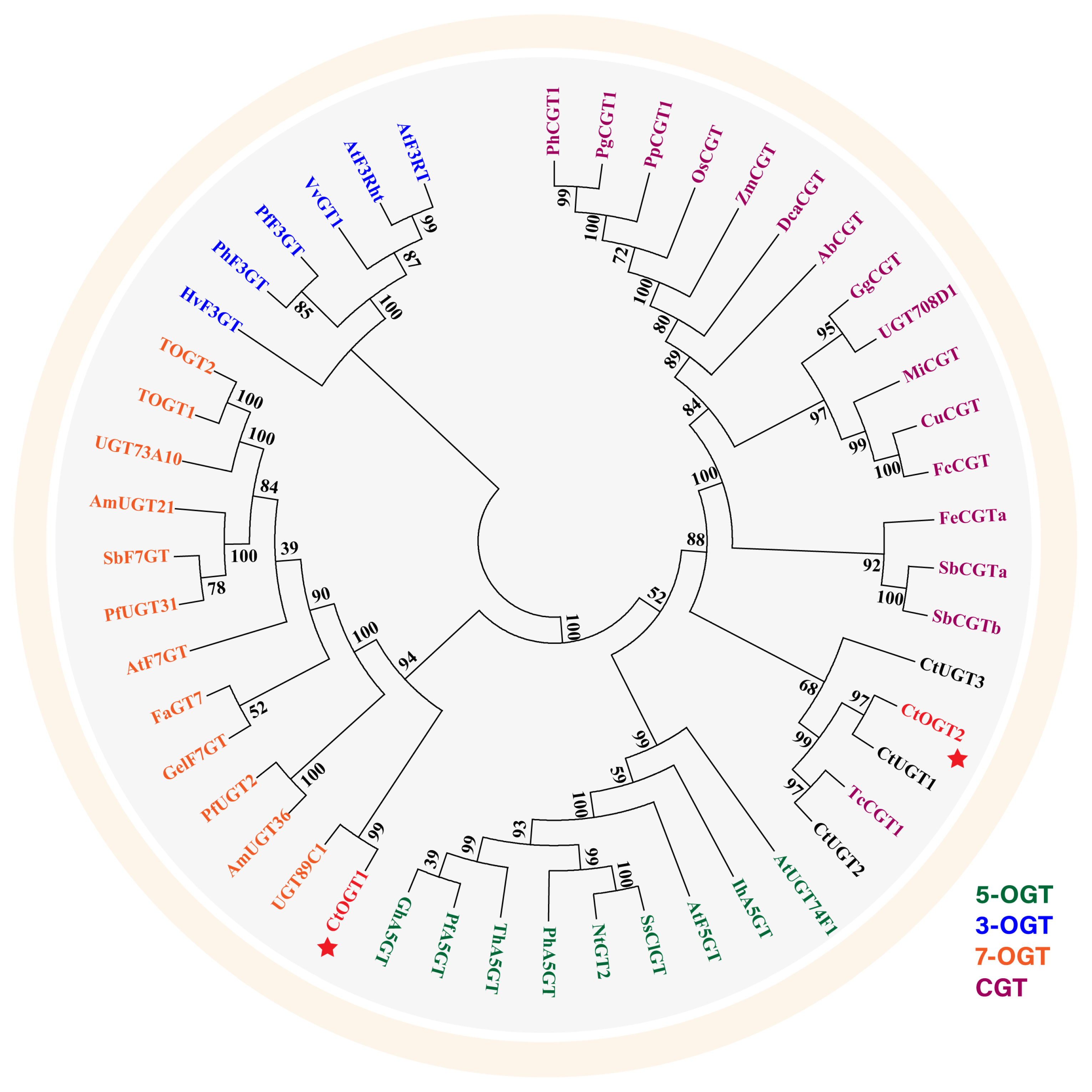
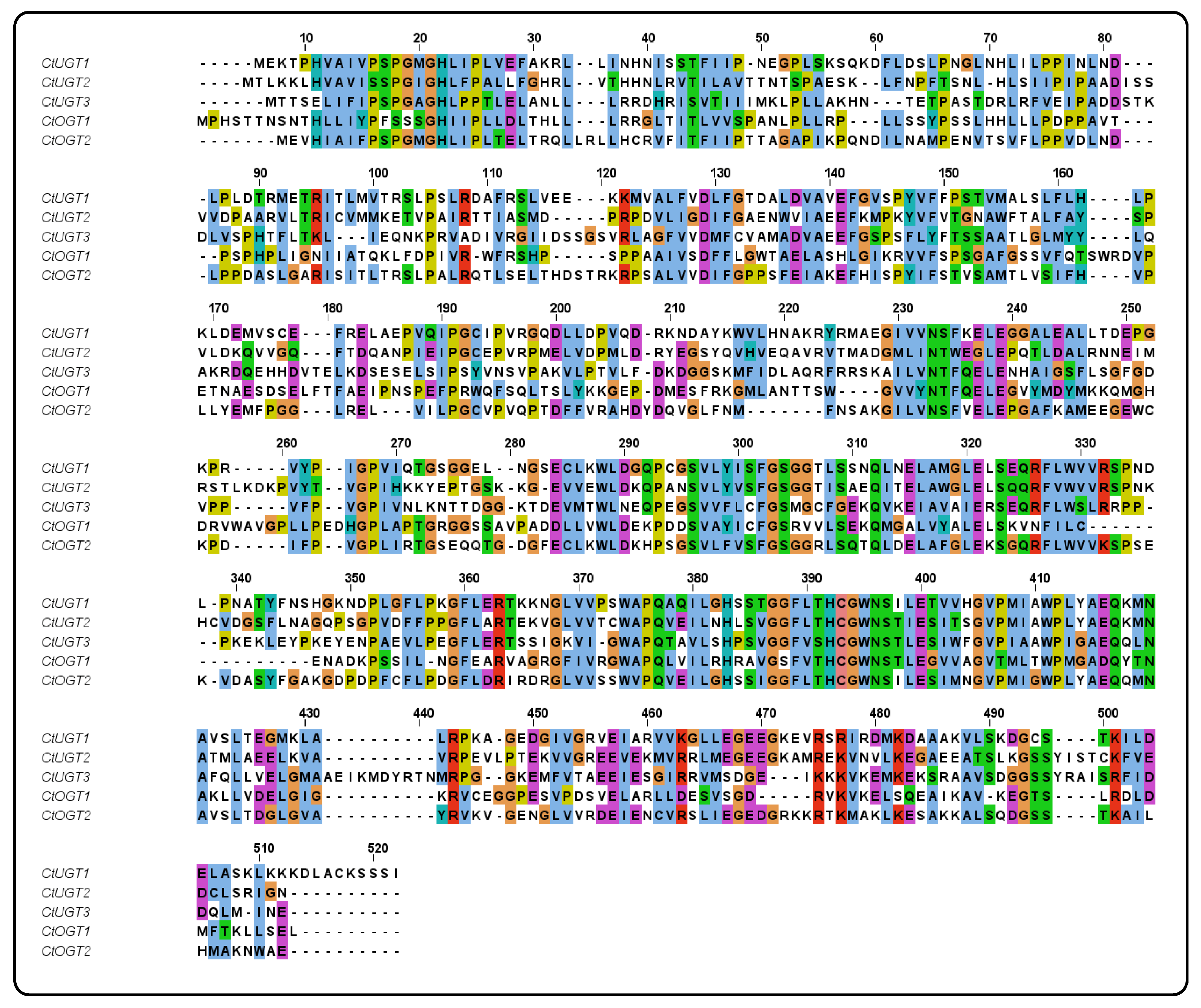
Using MEGA 6.0 software, a systematic evolutionary analysis of 48 glycosyltransferases, including the CtOGT1 and CtOGT2 genes, was conducted based on the neighbor-joining method with ClustalW multiple alignments [28]. The phylogenetic analysis revealed that UGTs can be divided into four main branches, including 3-O-glycosyltransferases, 5-O-glycosyltransferases, 7-O-glycosyltransferases, and C-glycosyltransferases, as shown in Figure 1. The results indicated that CtOGT1 clusters with the 7-O-glycosyltransferases branch, while CtOGT2 clusters with CtUGT1, CtUGT2, and CtUGT3, which were already reported to have O-glycosyltransferase activity in safflower, as well as TcCGT1, which had both C- and O-glycosyltransferase activity [29]. This predicted result indicates that the genes CtOGT1 and CtOGT2 belonged to O-glycosyltransferases in safflower. The amino acid sequence identity of CtOGT1, CtOGT2, CtUGT1, CtUGT2, and CtUGT3 was 48.39% (Figure 2).
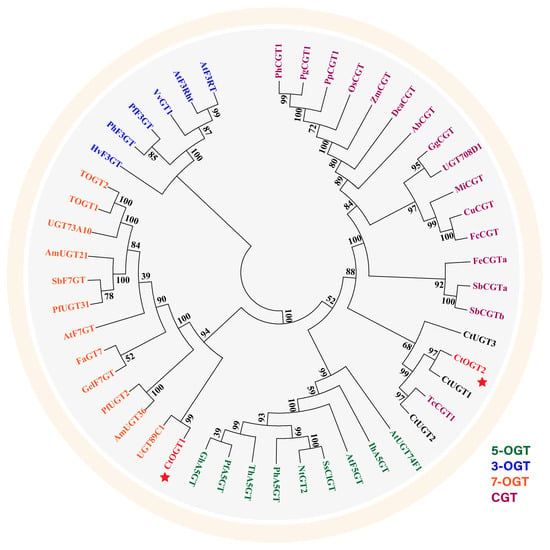
Figure 1.
Phylogenetic analyses of CtOGT1 and CtOGT2 with 46 other reported glycosyltransferases. PhCGT1 (MK616588, Phyllostachys heterocycla); PgCGT1 (MK616592, Phyllostachys glauca); PpCGT1 (MK616593, Phyllostachys prominens); OsCGT (FM179712, Oryza sativa); ZmCGT (NP_001132650, Zea mays); DcaCGT (QOD39011, Dendrobium catenatum); AbCGT (MN747045, Aloe barbadensis); GgCGT (QGL05036, Glycyrrhiza glabra); UGT708D1 (LC003312, Glycine max); MiCGT (KT200208, Glycyrrhiza glabra); CuCGT (LC131334, Citrus unshiu); FcCGT (LC131333, Fortunella crassifolia); FeCGTa (AB909375, Fagopyrum esculentum); SbCGTa (MK894443, Scutellaria baicalensis); SbCGTb (MK894444, Scutellaria baicalensis); TcCGT1 (MK644229, Trollius chinensis); CtUGT1 (OQ354214, Carthamus tinctorius L.); CtUGT2 (OQ354222, Carthamus tinctorius L.); CtUGT3 (OQ354223, Carthamus tinctorius L.); AtUGT74F1 (NP973682, Arabidopsis thaliana); IhA5GT (Q767C8, Iris hollandica); AtF5GT (AAM91686, Arabidopsis thaliana); SsClGT (AAK54465, Solanum sogarandinum); NtGT2 (BAB88935, Nicotiana tabacum); PhA5GT (BAA89009, Petunia hybrida); ThA5GT (BAC54093, Torenia hybrida); PfA5GT (BAA36421, Perilla frutescens); GhA5GT (BAA36423, Glandularia x hybrida); UGT89C1 (AAP31923, Arabidopsis thaliana); AmUGT36 (BAG16513, Antirrhinum majus); PfUGT2 (BAG31951, Perilla frutescens); GelF7GT (BAC78438, Glycyrrhiza echinate); AtF7GT (AAL90934, Arabidopsis thaliana); FaGT7 (Q2V6J9, Fragaria ananassa); PfUGT31 (BAG31952, Perilla frutescens); SbF7GT (BAA83484, Scutellaria baicalensis); UGT73A10 (BAG80536, Lycium barbarum); TOGT1 (AAK28303, Nicotiana tabacum); TOGT2 (AAK28304, Nicotiana tabacum); HvF3GT (CAA33729, Hordeum vulgare); PhF3GT (BAA89008, Petunia hybrida); PfF3GT (BAA19659, Perilla frutescens); VvGT1 (AAB81682, Vitis vinifera); AtF3Rht (AAM65321, Arabidopsis thaliana); AtF3RT (AAM91139, Arabidopsis thaliana). Note: The red stars represent glycosyltransferases characterized from safflower in this article.
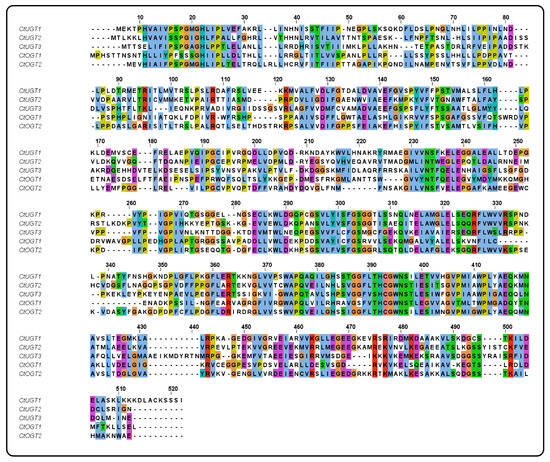
Figure 2.
The sequence alignment of genes CtOGT1, CtOGT2, CtUGT1, CtUGT2, and CtUGT3. Note: Different colors represent the homology of different sequences.
2.3. Cloning and Functional Characterization of CtOGTs
Using safflower cDNA as a template, PCR amplification was performed using high-fidelity polymerase Prime Start Max to obtain the complete ORF fragments of CtOGT1 and CtOGT2, which were 1365 bp and 1383 bp in length, respectively. These fragments were cloned into the pET28-MBP vector, resulting in recombinant expression plasmids pET28-MBP-CtOGT1 and pET28-MBP-CtOGT2. The recombinant plasmids were transformed into BL21(DE3) competent cells, induced with IPTG at 16 °C for 18 h, and purified using His-tag affinity chromatography. The purification of the recombinant pET28-MBP-CtOGT1 and pET28-MBP-CtOGT2 proteins was confirmed using SDS-PAGE analysis, as shown in Figure S2.
To verify the activity of CtOGT1 and CtOGT2 towards flavonoid O-glycosyltransferases in safflower, we selected flavanols (kaempferol, quercetin), flavones (apigenin, scutellarein), dihydrochalcone (phloretin), isoflavones (genistein, daidzein), flavanones (naringenin, eriodictyol, glycyrrhizin), and flavanonol (dihydrokaempferol) as substrates for detection. The enzymatic reaction system consisted of 50 mM Na2HPO4-NaH2PO4 buffer (pH 8.0), 0.2 mM substrate, 0.5 mM UDP-Glc, and 10 μg of purified enzyme. After incubating at 37 °C for 6 h, the reaction was terminated with double the volume of methanol.
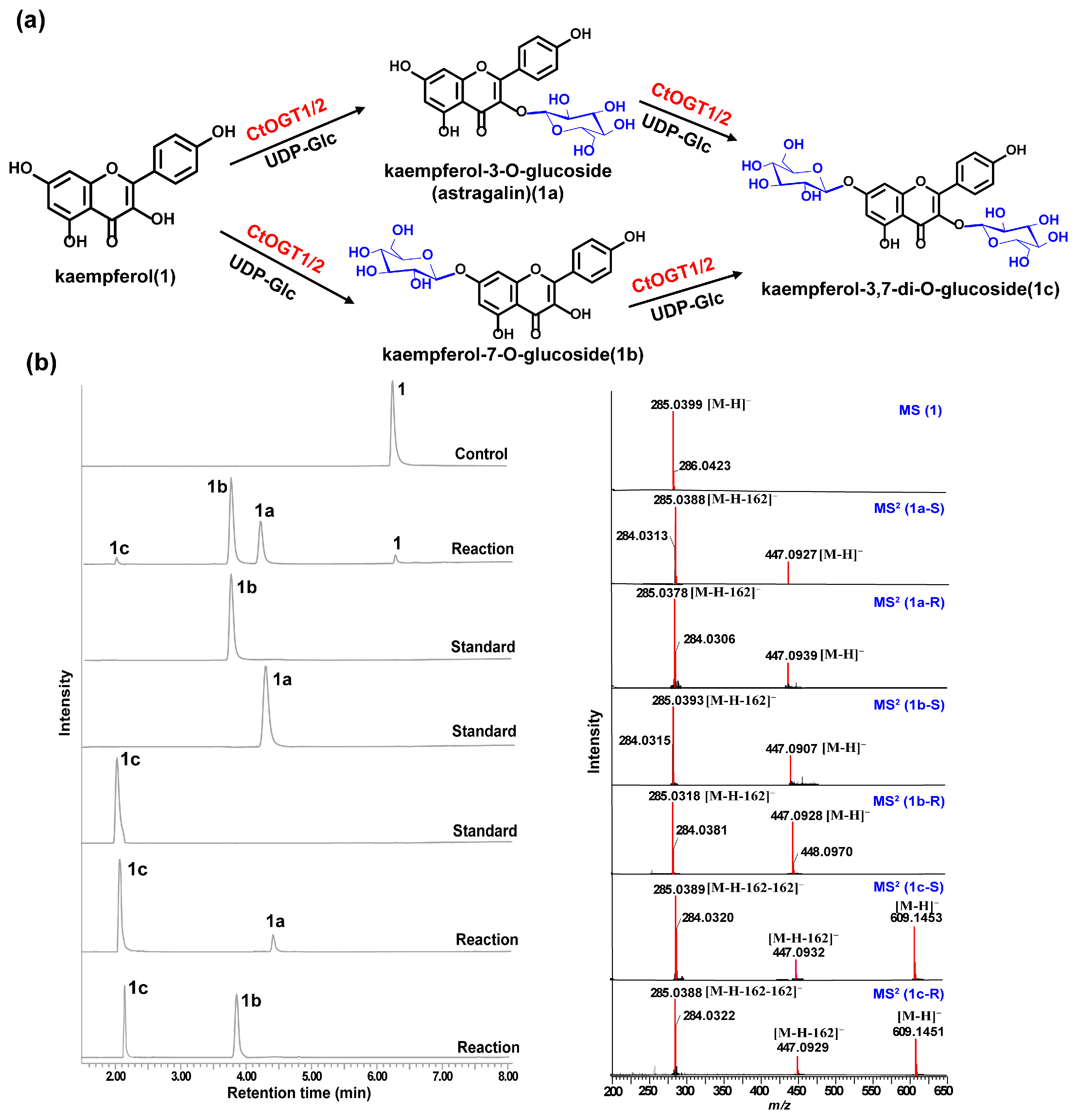
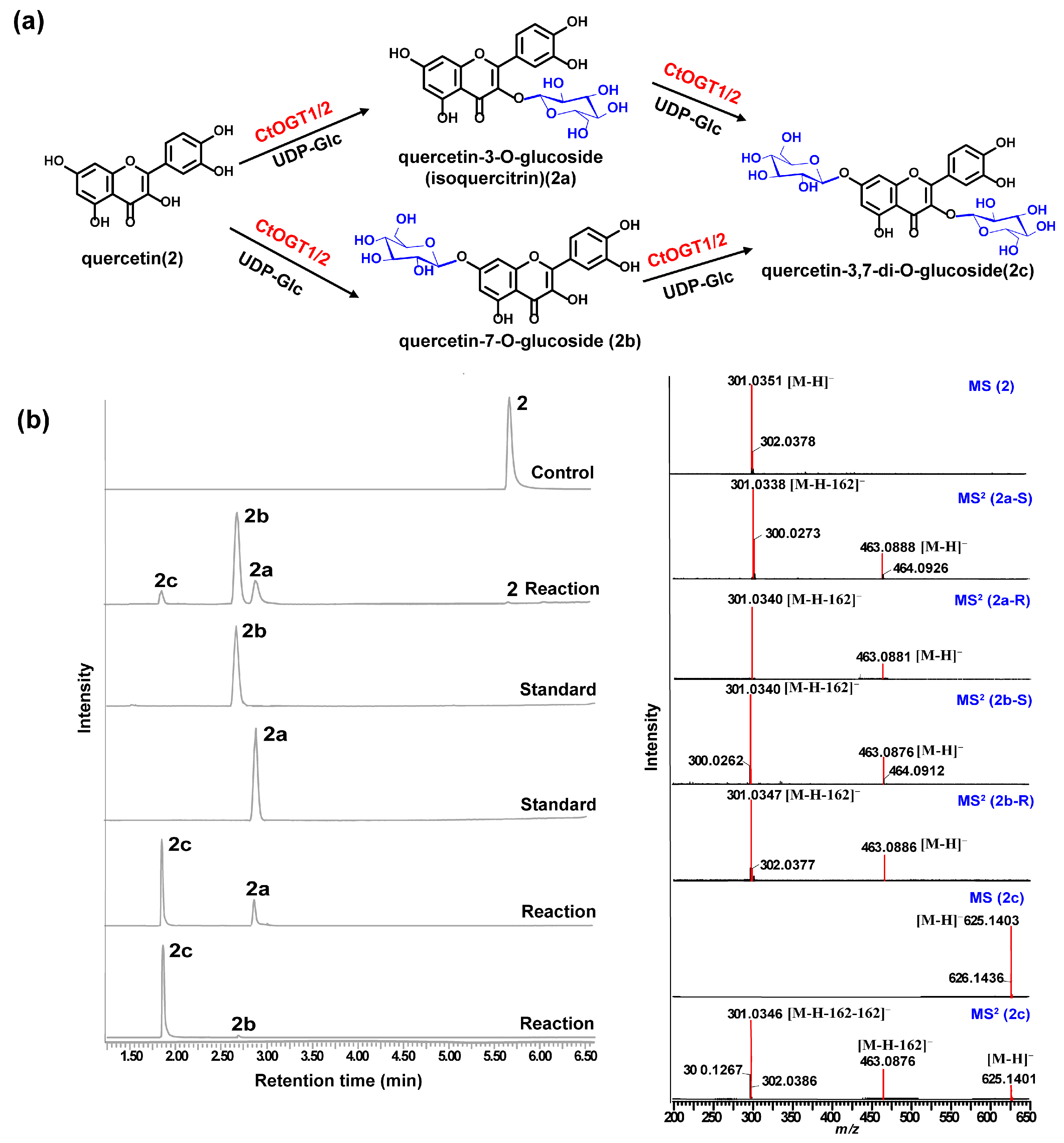
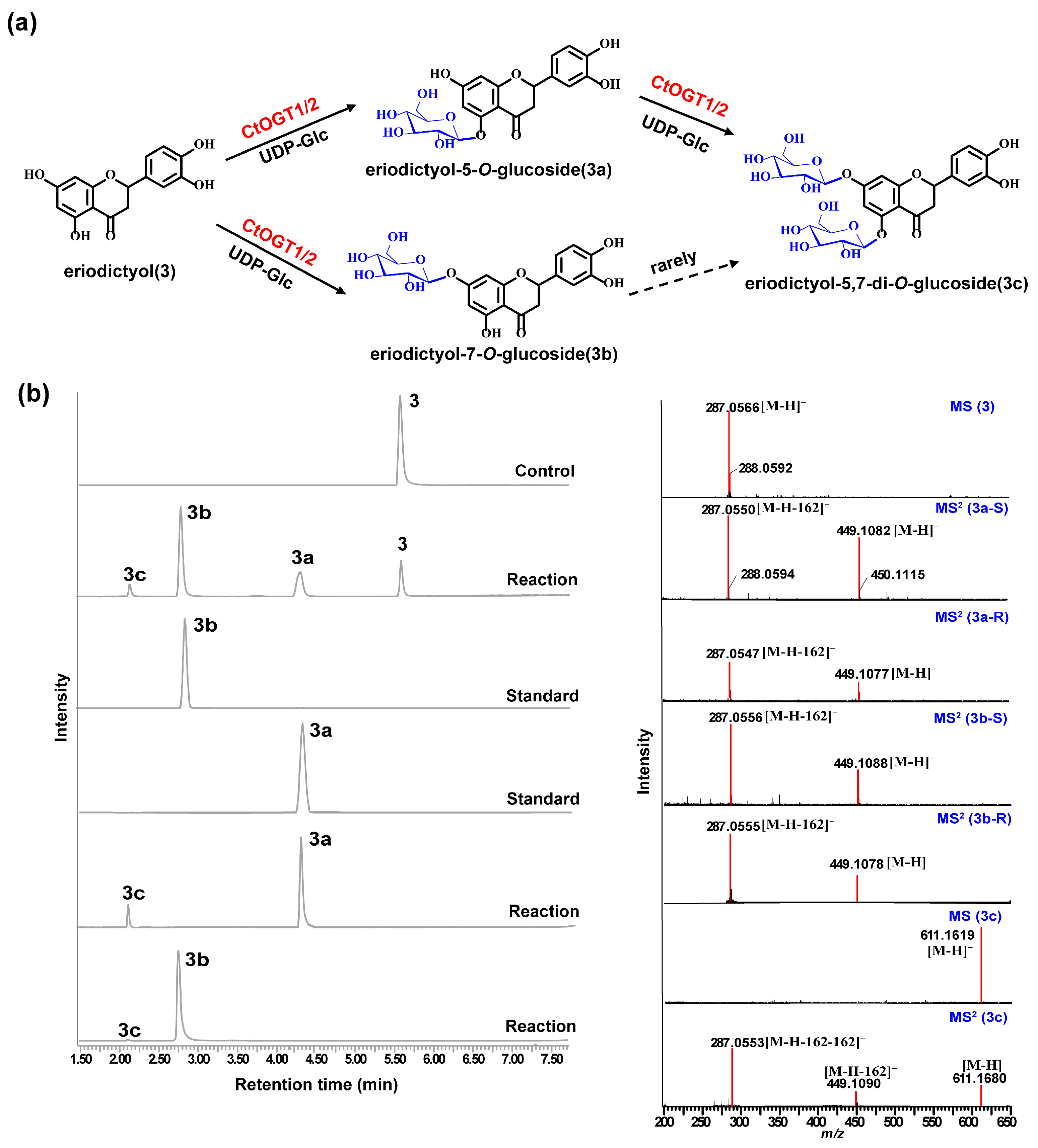
Through liquid chromatography combined with mass spectrometry (LC/MS) analysis and comparison with the standard references, the results showed that, compared to the control group, the genes CtOGT1 and CtOGT2 could respectively catalyze the conversion of kaempferol 3-OH and 7-OH into kaempferol-7-O-glucoside and kaempferol-3-O-glucoside (astragalin) (1a and 1b). Additionally, the presence of kaempferol-3,7-di-O-glucoside (1c) was detected. CtOGT1 exhibited better catalytic activity for di-O-glycosylation compared to CtOGT2. When quercetin was used as a substrate, both CtOGT1 and CtOGT2 catalyzed the conversion of quercetin 3-OH and 7-OH into isoquercitrin, quercetin-7-O-glucoside, and quercetin-3,7-di-O-glucoside (2a, 2b and 2c). When eriodictyol was used as a substrate, CtOGT1 and CtOGT2 catalyzed the formation of eriodictyol-5-O-glucoside, eriodictyol-7-O-glucoside, and eriodictyol-5,7-di-O-glucoside (3a, 3b and 3c), as shown in Figure 3, Figure 4 and Figure 5. Then, enzymatic reactions were conducted in vitro using substrates 1a, 1b, 2a, 2b, 3a, and 3b. As a result, it was found that compound 1c could be detected when using 1a and 1b as substrates, indicating that CtOGT1 and CtOGT2 can catalyze the formation of 1c from 1, 1a, and 1b, respectively. Similarly, compound 2c could also be detected when using 2, 2a, and 2b as substrates. However, when using 3b as a substrate, only a small amount of 3c was detected. This indicated that CtOGT1 and CtOGT2 could catalyze the formation of 3c from 3 and 3a, but it was difficult to catalyze the formation of 3c from 3b. Furthermore, CtOGT1 and CtOGT2 exhibited better O-glycosyltransferase activity towards substrates apigenin, scutellarein, phloretin, genistein, daidzein, naringenin, glycyrrhizin, and dihydrokaempferol (4–11), indicating a broad substrate specificity of CtOGT1 and CtOGT2, as shown in Figures S3–S10 (all reactions were plotted using CtOGT1 as an example). These in vitro enzymatic activity results were consistent with the predicted result, indicating that the genes CtOGT1 and CtOGT2 belonged to O-glycosyltransferases in safflower.
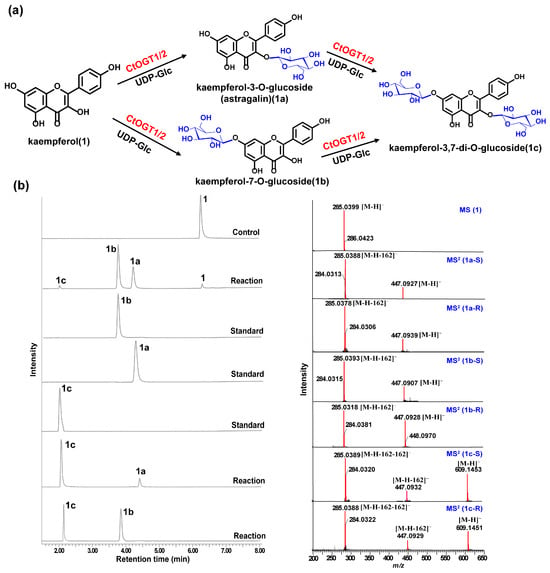
Figure 3.
(a) LC/MS analysis of CtOGT1- and CtOGT2-catalyzed product using kaempferol as the substrate. Compounds 1, 1a, 1b, and 1c represent kaempferol, kaempferol-7-O-glucoside, kaempferol-3-O-glucoside, and kaempferol-3,7-di-O-glucoside, respectively. (b) (−)-ESI-MS spectra of 1, (−)-ESI-MS2 spectra of 1a, 1b, and 1c. S: standard, R: reaction.
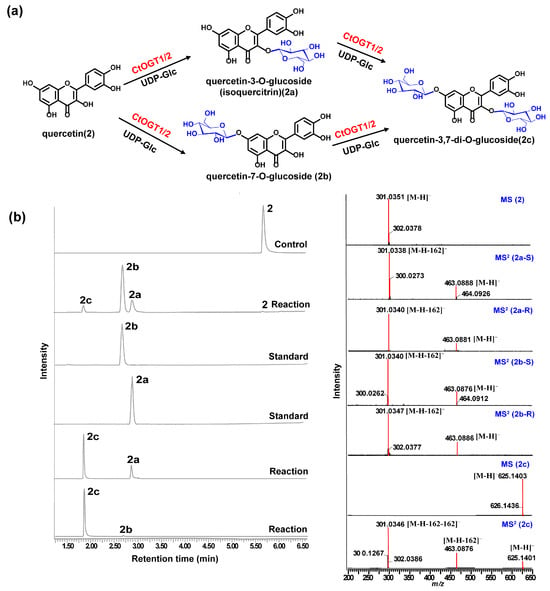
Figure 4.
(a) LC/MS analysis of CtOGT1- and CtOGT2-catalyzed product using quercetin as the substrate. Compounds 2, 2a, 2b, and 2c represent quercetin, isoquercitrin, quercetin-7-O-glucoside, and quercetin-3,7-di-O-glucoside respectively. (b) (−)-ESI-MS spectra of 2 and 2c, (−)-ESI-MS2 spectra of 2a, 2b, and 2c. S: standard, R: reaction.
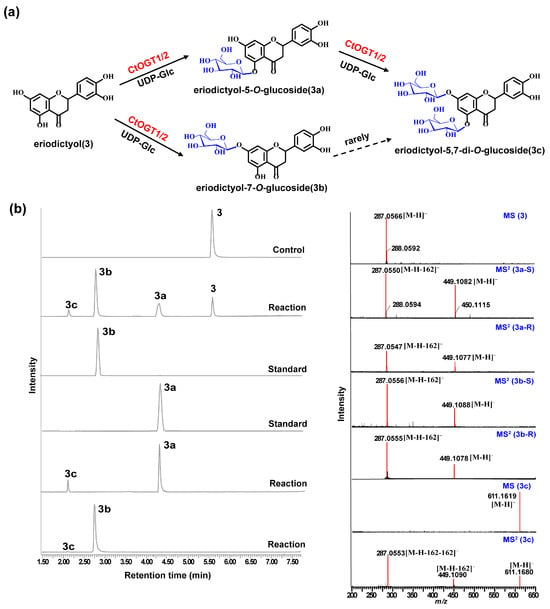
Figure 5.
(a) LC/MS analysis of CtOGT1- and CtOGT2-catalyzed product using eriodictyol as the substrate. Compounds 3, 3a, 3b, and 3c represent eriodictyol, eriodictyol-5-O-glucoside, eriodictyol-7-O-glucoside, and eriodictyol-5,7-di-O-glucoside, respectively. (b) (−)-ESI-MS spectra of 3 and 3c, (−)-ESI-MS2 spectra of 3a, 3b, and 3c. S: standard, R: reaction.
3. Discussion
Safflower contains quinone chalcone compounds with unique structures and significant therapeutic activities in cardiovascular and cerebrovascular diseases. Examples of these compounds include HSYA, safflor yellow A, and carthamin. These compounds are exclusively found in safflower and belong to the C-glycoside class. Therefore, the biosynthesis pathway of these special flavonoids in safflower has attracted researchers’ attention [17]. Common flavonoid compounds in safflower are also found in many other species and display various activities. These include kaempferol, naringenin, quercetin, apigenin, and luteolin, among others [30].
Xie et al. discovered a novel glycosyltransferase, UGT73AE1, from safflower. UGT73AE1 demonstrated the ability to glucosylate 19 structurally diverse acceptor molecules and generate O-, S-, and N-glycosides, making it the first reported plant glycosyltransferase with tri-functional activity [31]. Qi et al. found that CtUGT4 can catalyze the formation of quercetin-7-O-glucoside from quercetin and naringenin-3-O-glucoside from naringenin in vitro. Additionally, a range of new products were generated using various compounds as substrates, including flavones, flavonols, flavanones, chalcones, and chalcone glycosides [25]. Studies have shown that CtUGT3 catalyzes the glucosylation of flavones (apigenin), flavanones (naringenin), flavonols (kaempferol, quercetin, and isorhamnetin), and chalcones (naringenin chalcone), forming their respective O-glucosides. Furthermore, when flavonols were used as aglycones in vitro, CtUGT3 exhibited both 7-OH and 3-OH glycosylation activities simultaneously [24]. The most abundant flavonoid glycosides in plants are flavone O- or C-glycosides and flavonol O-glycosides, with glycosides at the 3-OH or 7-OH positions being the most common. According to reports, the genes UGT78K1 and Fh3GT1 both possess the activity of flavonoid 3-O-glycosyltransferase [32,33]. Wang et al. discovered a highly selective and donor-diverse 3-O-glycosyltransferase, Sb3GT1 (UGT78B4), from Scutellaria baicalensis. Sb3GT1 was capable of accepting five sugar donors (UDP-Glc/-Gal/-GlcNAc/-Xyl/-Ara) to catalyze the 3-O-glycosylation reaction of 17 flavonoid compounds, with conversion rates exceeding 98% [34]. CsUGT75L12 encoded a flavonoid 7-O-glucosyltransferase, which can specifically transfer the glucose moiety from UDP-glucose to the 7-hydroxyl position of flavonoids, generating the corresponding 7-O-glucosides. Furthermore, the expression pattern of the CsUGT75L12 gene was consistent with the accumulation pattern of 7-O-glucosides and 7-O-neohesperidoside in tea plants, indicating its involvement in the biosynthesis of bitter flavonoid 7-O-neohesperidoside [35]. In addition, genes such as NpUGT6, UGT73CD1, TwUGT3, and UGT71E5 have been found to exhibit catalytic activity in the O-glycosylation of various flavonoid compounds [36,37,38,39]. However, there is currently a relatively limited amount of research on glycosyltransferases in safflower. Despite the presence of abundant flavonoid compounds in safflower that are typically conjugated with sugar moieties to form glycosides, our understanding of the specific glycosyltransferases involved in catalyzing these reactions remains limited. Therefore, further in-depth studies on glycosyltransferases in safflower are necessary.
In this study, CtOGT1 and CtOGT2, as newly discovered O-glycosyltransferases in safflower, have shown significant versatility and catalytic activity. They exhibit O-glycosylation activity towards a wide range of flavonoid compounds, including flavanols (kaempferol, quercetin), flavones (apigenin, scutellarein), dihydrochalcone (phloretin), isoflavones (genistein, daidzein), flavanones (naringenin, eriodictyol, glycyrrhizin), and flavanonol (dihydrokaempferol). Notably, CtOGT1 and CtOGT2 demonstrate an ability to catalyze the addition of glucose moieties to specific hydroxyl positions (such as 7-OH, 3-OH, and 5-OH) of various flavonoid structures. By catalyzing the formation of specific glycosides, such as kaempferol-7-O-glucoside, astragalin, kaempferol-3,7-di-O-glucoside, quercetin-7-O-glucoside, isoquercitrin, quercetin-3,7-di-O-glucoside, eriodictyol-7-O-glucoside, eriodictyol-5-O-glucoside, and eriodictyol-5,7-di-O-glucoside, CtOGT1 and CtOGT2 contribute to the synthesis of diverse flavonoid glycosides with medicinal value. These glycosides play crucial roles in various biological processes and contribute to the unique properties, taste, and health benefits associated with safflower. The discovery and characterization of CtOGT1 and CtOGT2 shed light on the glycosylation pathways involved in safflower’s flavonoid metabolism. These findings not only enhance our understanding of secondary metabolite biosynthesis in safflower but also provide valuable insights into the regulation of bitterness and other sensory attributes in safflower-based products.
Synthetic biology has played a crucial role in the research of flavonoid O-glucosides. Through genetic engineering techniques, glucosyltransferases (GTs) from various sources can be introduced into host microorganisms to enable the enzymatic glucosylation of flavonoid compounds. Common expression hosts for these GTs include E. coli and Saccharomyces cerevisiae. By adjusting reaction conditions and substrate concentrations, the efficient and controllable synthesis of flavonoid O-glucosides can be achieved. Flavonoid 7-O-glucosides exhibit various biological activities; however, some are not abundant in nature. Therefore, a method for producing flavonoid 7-O-glucosides was investigated. E. coli-expressing tobacco-derived glucosyltransferase (Ec-NtGT2) converted several flavonoid compounds (apigenin, luteolin, quercetin, kaempferol, and naringenin) into their respective 7-O-glucosides, with conversion rates ranging from 67% to 98%. In scaled-up production, Ec-NtGT2 yielded 24 mg/L of apigenin 7-O-glucoside, 41 mg/L of luteolin 7-O-glucoside, 118 mg/L of quercetin 7-O-glucoside, 40 mg/L of kaempferol 7-O-glucoside, and 75 mg/L of naringenin 7-O-glucoside through the sequential addition of substrates within 4–9 h. The conversion rates for apigenin, luteolin, quercetin, kaempferol, and naringenin were 97%, 72%, 77%, 98%, and 96%, respectively. These results indicated that Ec-NtGT2 was a simple and efficient biotransformation system for the production of flavonoid 7-O-glucosides [40]. Zhao et al. achieved de novo biosynthesis of the isoflavones genistein in E. coli. The results showed that the LCA2G-LNR30R-LGN43 three-strain system was more suitable for synthesizing it. Under the optimal inoculation ratio of 2:1:4, the production of genistein reached 35.1 mg/L [41]. Li et al. biosynthesized baicalin and scutellarein in E. coli, with yields of 23.6 mg/L and 106.5 mg/L, respectively [42]. Liu et al. established a yeast cell factory for the production of breviscapine, with a titer reaching 108 mg/L [43]. Nielsen et al. constructed a de novo synthesis yeast cell factory for isoflavones biosynthesis, producing the core chemical scaffold genistein, with titers of 72.8 mg/L for puerarin and 73.2 mg/L for daidzein, by introducing glycosyltransferases in the engineered strains [44]. Synthetic biology provides a sustainable and efficient approach to the synthesis of flavonoid O-glucosides. It can be utilized not only for large-scale production but also for the targeted synthesis of specific structures of flavonoid O-glucosides. This is of great significance in studying the biological activities of flavonoid compounds and drug development, providing new insights and tools for further research and applications in related fields. As a precious and highly valued traditional Chinese medicine with immense medicinal value, it is of the utmost importance to decipher the biosynthetic pathways of the major compounds in safflower and achieve efficient heterologous production.
In summary, we successfully characterized two new di-O-glycosyltransferases, CtOGT1 and CtOGT2, from Carthamus tinctorius L., a medicinal plant. It demonstrated efficient catalytic activity in di-O-glycosylation of at least seven substrates containing flavonols (kaempferol, quercetin), flavone (scutellarein), dihydrochalcone (phloretin), and flavanones (naringenin, eriodictyol, glycyrrhizin), as well as mono-O-glycosylation of at least 11 flavonoid compounds. This study provides valuable insights for future research on the biosynthetic pathways of flavonoids in safflower.
4. Materials and Methods
4.1. Plant Materials
Safflower (Carthamus tinctorius L.) samples were obtained from the Henan Academy of Agricultural Sciences. The collected samples were washed with sterile deionized water and subsequently frozen in liquid nitrogen. They were then stored at −80 °C until RNA extraction.
4.2. Phylogenetic Analysis and Sequence Alignment
To perform sequence comparisons and analysis, MEGA 6.0 software was utilized. Subsequently, a phylogenetic tree was constructed using the neighbor-joining method for cluster analysis, incorporating CtOGT1 and CtOGT2 along with other glycosyltransferase sequences. The amino acid sequences of CtOGT1, CtOGT2, CtUGT1, CtUGT2, and CtUGT3 were aligned using DNAMAN 8.0 software for multiple sequence alignment [45].
4.3. Molecular Cloning
To identify potential OGT genes, a BLAST search was performed on the transcriptome database of Carthamus tinctorius L., using Vitis vinifera’s glycosyltransferase VvGT1 and Arabidopsis thaliana’s glycosyltransferase UGT89C1 as reference sequences. This search led to the selection of two candidate genes, namely CtOGT1 and CtOGT2, which possessed complete open reading frames (ORFs). For detailed sequence information, please refer to Table S2. The ExPASy online server (http://web.expasy.org/protparam/, accessed on 8 May 2023) was utilized to predict various characteristics encoded by CtOGT1 and CtOGT2. Furthermore, the protein structures of CtOGT1 and CtOGT2 were predicted using AlphaFold (https://alphafold.com/, accessed on 10 May 2023).
Total RNA was isolated from safflower samples using the TranZolTM kit (Transgen Biotech, Beijing, China). Reverse transcription was performed to convert the extracted RNA into cDNA using the TransScript II One-Step gDNA Removal and cDNA Synthesis SuperMix (Transgen Biotech, China). Specific primers were designed for amplifying the candidate O-glycosyltransferase genes. The amplification reactions were carried out using 2× Prime Start Max Enzyme under the following conditions: 5 min at 98 °C; 15 s at 98 °C, 15 s at 55 °C, 50 s at 72 °C for 40 cycles; and 7 min at 72 °C. The resulting PCR products were then ligated into the pEASY-Blunt Zero Cloning Kit vector (TsingKe, Beijing, China) and sequenced to confirm their integrity. Subsequently, using a positive plasmid as a template, the target gene with BamHI restriction sites was cloned. The BamHI-digested pET28a-MBP vector and the target gene fragment were seamlessly joined together using seamless cloning technique. The constructed recombinant plasmids were transformed into E. coli BL21 (DE3) cells (Transgen Biotech) for heterologous expression on LB plates supplemented with kanamycin (50 mg/L). Detailed primer sequences used in this study can be found in Table 2.
Table 2.
The design of primer sequences for candidate genes.
4.4. Expression and Purification of CtOGT1 and CtOGT2
The recombinant expression plasmids were separately transformed into BL21(DE3) competent cells. After the appearance of bacterial colonies, single clones were picked and expanded for cultivation until the OD600 reached 0.6–0.7. Then, isopropyl-β-d-thiogalactopyranoside (IPTG) was added, and the cells were induced at a low temperature of 16 °C for 18 h. Following induction, the cells were harvested through centrifugation at 8000 rpm for 5 min at 4 °C. The resulting cell pellet was resuspended in 5 mL of lysis buffer (50 mM Na2HPO4-NaH2PO4, pH 8.0, 300 mM NaCl) and sonicated in an ice bath. Subsequent removal of cell debris was accomplished by centrifugation at 12,000 rpm for 10 min at 4 °C, with the supernatant collected for further use. The collected supernatant was mixed with Ni-NTA resin (DP101, Transgen Biotech, China) and incubated with agitation for 1.5 h at 4 °C. Using a 50 mM Na2HPO4-NaH2PO4 buffer (pH 8.0) containing 20 mM imidazole and 300 mM NaCl, the resin was washed with 10–20 mL of buffer. Elution was performed sequentially using a range of imidazole concentrations in the buffer salt (20/50/150/200/250 mM), with 2–5 mL fractions collected at each step. Next, 15 μL of collected fractions was mixed with 5 μL of 4× protein loading buffer (LABLEAD, Beijing, China) for SDS-PAGE gel electrophoresis. The fractions containing the target protein were concentrated using a 30 kDa ultrafiltration tube (15 mL) at 4000 rpm to a final volume of 500 μL. The concentrated protein was exchanged with a 50 mM Na2HPO4-NaH2PO4 buffer salt (pH 8.0) to replace the imidazole elution solution. The purified CtOGT1 and CtOGT2 enzymes were stored at −80 °C in the presence of glycerol for long-term storage. The protein concentration was determined using the Protein Quantitative Kit (TransGen Biotech), employing bovine serum albumin (BSA) as the standard.
4.5. Enzyme Activity Assay
To assess the functionality of CtOGT1 and CtOGT2, various substrates (kaempferol, quercetin, apigenin, scutellarein, phloretin, genistein, daidzein, naringenin, eriodictyol, glycyrrhizin, and dihydrokaempferol) were employed. UDP-Glc served as the sugar donor in the glycosylation reactions. The reaction mixture comprised 0.5 mM UDP-Glc, 0.1 mM substrate, protein, and a buffer containing 50 mM Na2HPO4-NaH2PO4 and 300 mM NaCl (pH 8.0), making a final volume of 500 μL. Incubation of the reactions took place at 37 °C for 6 h, followed by termination with double the volume of methanol. Supernatants collected after centrifugation at 15,000 rpm for 15 min were subjected to LC/MS analysis. Each substrate underwent three parallel reactions.
4.6. General Procedures
Compounds 1–11, 1a, 1b, 2a, 2b, 3a, and 3b were acquired from Beite Renkang (Beijing, China), while UDP-glucose was obtained from Sigma-Aldrich (St. Louis, MO, USA). An UPLC-QTOF-MS system (Waters Technologies, Milford, MA, USA) was employed to analyze all products. The Acquity UPLC utilized a T3 column (2.1 mm × 100 mm, 1.8 μm particle size; Waters Technologies) held at 40 °C. The mobile phase consisted of water containing 0.1% (v/v) formic acid (A) and acetonitrile containing 0.1% (v/v) formic acid (B). A linear gradient elution program was applied with the following conditions: 0 min, 90% A; 1 min, 80% A; 3 min, 80% A; 4 min, 75% A; 8 min, 30% A; 9 min, 5% A; 12 min, 5% A; 12.2 min, 90% A; 15.2 min, 90% A. For MS analysis, the effluent was introduced into the ESI source of the mass spectrometer at a flow rate of 0.4 mL/min using a T-union splitter. The mass spectrometer operated in (−)-ESI mode.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules29030604/s1, Table S1: Physicochemical analysis of the amino acids encoded by CtOGT1 and CtOGT2; Table S2: The sequences of CtOGT1 and CtOGT2 in Carthamus tinctorius L.; Figure S1: (a) The three-dimensional protein structure prediction of CtOGT1; (b) the three-dimensional protein structure prediction of CtOGT2; Figure S2: SDS-PAGE analysis of recombinant CtOGT1 and CtOGT2 purified via affinity chromatography. Note: M: Marker; Figure S3: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using apigenin as the substrate; Figure S4: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using scutellarein as the substrate; Figure S5: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using phloretin as the substrate; Figure S6: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using genistein as the substrate; Figure S7: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using daidzein as the substrate; Figure S8: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using naringenin as the substrate; Figure S9: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using glycyrrhizin as the substrate; Figure S10: LC-MS analysis of CtOGT1- and CtOGT2-catalyzed product using dihydrokaempferol as the substrate.
Author Contributions
Project administration and funding acquisition, L.H. and P.S.; X.X., M.X., P.S. and Y.Z. were responsible for methodology, validated the data, performed formal analysis, and reviewed and edited the manuscript; X.X., Y.H., H.T., Y.C., X.S. and S.Y. performed experiments; X.X. prepared the original draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the scientific and technological innovation project of the China Academy of Chinese Medical Sciences (CI2021B014 and CI2023E002), Fundamental Research Funds of the China Academy of Chinese Medical Sciences (ZZ16-YQ-042), and Key project at central government level: The establishment of sustainable use for valuable Chinese medicine resources (2060302).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article or Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sung, J.S.; Ko, H.C.; Hur, O.S.; Sang, G.K.; Kyoung, Y.R. Morphological and oil compositions in safflower (Carthamus tinctorius L.) germplasm of different geographical groups. Korean J. Agric. Sci. 2016, 28, 84–91. [Google Scholar] [CrossRef]
- Delshad, E.; Yousefi, M.; Sasannezhad, P.; Rakhshandeh, H.; Ayati, Z. Medical uses of Carthamus tinctorius L. (Safflower): A comprehensive review from Traditional Medicine to Modern Medicine. Electron. Physician 2018, 4, 6672–6681. [Google Scholar] [CrossRef] [PubMed]
- Ao, H.; Feng, W.; Peng, C. Hydroxysafflor Yellow A: A promising therapeutic agent for a broad spectrum of diseases. Evid.-Based Complement. Altern. Med. 2018, 2018, 8259280. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Xue, Y.; Guo, D.; Sun, L.; Guo, M. Carthami Flos: A review of its ethnopharmacology, pharmacology and clinical applications. Rev. Bras. 2015, 25, 553–566. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J.; Kim, S.J.; Kim, J.H. Quercetin-3-O-glucoside suppresses pancreatic cancer cell migration induced by tumor-deteriorated growth factors in vitro. Oncol. Rep. 2016, 35, 2473–2479. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharmacother. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Elizabeth, T.D.; Diego, M.E.; Oswaldo, O.M.; Liliana, S.C. Rutin: Family farming products’ extraction sources, industrial applications and current trends in biological activity protection. Molecules 2023, 28, 5864. [Google Scholar]
- Caparica, R.; Júlio, A.; Araújo, M.E.M.; Baby, A.R.; Fonte, P.; Costa, J.G.; Santos, D.A.T. Anticancer activity of rutin and its combination with ionic liquids on renal cells. Biomolecules 2020, 10, 233. [Google Scholar] [CrossRef]
- Nishina, A.; Sato, D.; Yamamoto, J.; Kobayashi-Hattori, K.; Hirai, Y.; Kimura, H. Antidiabetic-like effects of naringenin-7-O-glucoside from edible chrysanthemum ‘Kotobuki’ and naringenin by activation of the PI3K/Akt pathway and PPARγ. Chem. Biodivers. 2019, 16, e1800434. [Google Scholar] [CrossRef]
- Penso, J.; Cordeiro, K.C.; da Cunha, C.R.; da Silva Castro, P.F.; Martins, D.R.; Lião, L.M.; Rocha, M.L.; de Oliveira, V. Vasorelaxant activity of 7-β-O-glycosides biosynthesized from flavonoids. Eur. J. Pharmacol. 2014, 733, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Z.; Gao, S.; Cheng, Y.N.; Sun, Y.Z.; Liu, W.; Tang, L.L.; Ren, D.M. Protective effect of naringenin-7-O-glucoside against oxidative stress induced by doxorubicin in H9c2 cardiomyocytes. Biosci. Trends 2012, 6, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Seo, Y. Antiadipogenic activity of isohamnetin 3-O-β-D-glucopyranoside from Salicornia herbacea. Immunopharmacol. Immunotoxicol. 2012, 34, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Devi, V.G.; Rooban, B.N.; Sasikala, V.; Sahasranamam, V.; Abraham, A. Isorhamnetin-3-glucoside alleviates oxidative stress and opacification in selenite cataract in vitro. Toxicol. In Vitro 2010, 24, 1662–1669. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.A.; Xu, X.; Wang, Y.; Shie, P.H.; Qiu, L. A new anti-inflammatory flavonoid glycoside from tetraena aegyptia. Nat. Prod. Res. 2021, 35, 1985–1990. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Islam, M.S.; Uddin, M.N.; Hasan, M.M.I.; Akanda, M.R. The potential health benefits of the isoflavone glycoside genistin. Arch. Pharm. Res. 2020, 43, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Tang, Y.; Li, S.; Duan, J.A. Chemical and biological properties of quinochalcone C-glycosides from the florets of Carthamus tinctorius. Molecules 2013, 18, 15220–15254. [Google Scholar] [CrossRef]
- Xian, B.; Wang, R.; Jiang, H.; Zhou, Y.; Yan, J.; Huang, X.; Chen, J.; Wu, Q.; Chen, C.; Xi, Z.; et al. Comprehensive review of two groups of flavonoids in Carthamus tinctorius L. Biomed. Pharmacother. 2022, 153, 113462. [Google Scholar] [CrossRef]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S.; et al. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1975. [Google Scholar] [CrossRef]
- Cornell, H.V.; Hawkins, B.A. Herbivore responses to plant secondary compounds: A test of phytochemical coevolution theory. Am. Nat. 2003, 161, 507–522. [Google Scholar] [CrossRef]
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yao, M.; Wang, Y.; Ding, M.; Zha, J.; Xiao, W.; Yuan, Y. Advances in engineering UDP-sugar supply for recombinant biosynthesis of glycosides in microbes. Biotechnol. Adv. 2020, 41, 107538. [Google Scholar] [CrossRef] [PubMed]
- Hoffmeister, D.; Dräger, G.; Ichinose, K.; Rohr, J.; Bechthold, A. The C-Glycosyltransferase UrdGT2 is unselective toward d- and l-configured nucleotide-bound rhodinoses. J. Am. Chem. Soc. 2003, 125, 4678–4679. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Xi, Z.; Xian, B.; Chen, C.; Huang, X.; Jiang, H.; Chen, J.; Peng, C.; Pei, J. Identification and characterization of CtUGT3 as the key player of astragalin biosynthesis in Carthamus tinctorius L. J. Agric. Food Chem. 2023, 71, 16221–16232. [Google Scholar] [CrossRef]
- Qi, S.; He, B.; Wang, H.; Duan, Y.; Wang, L.; Gao, Y.; Guo, M. A muti-substrate flavonol O-glucosyltransferases from safflower. Molecules 2023, 28, 7613. [Google Scholar] [CrossRef]
- Hiromoto, T.; Honjo, E.; Noda, N.; Tamada, T.; Kazuma, K.; Suzuki, M.; Blaber, M.; Kuroki, R. Structural basis for acceptor-substrate recognition of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase from Clitoria ternatea. Protein Sci. 2015, 24, 395–407. [Google Scholar] [CrossRef]
- Zong, G.; Li, J.; Gao, Y.; Fei, S.; Liu, X.; Wang, X.; Shen, Y. Overexpression, purification, biochemical and structural characterization of rhamnosyltransferase UGT89C1 from Arabidopsis thaliana. Protein Expr. Purif. 2019, 156, 44–49. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 12, 2725–2729. [Google Scholar] [CrossRef]
- He, J.B.; Zhao, P.; Hu, Z.M.; Liu, S.; Kuang, Y.; Zhang, M.; Li, B.; Yun, C.H.; Qiao, X.; Ye, M. Molecular and structural characterization of a promiscuous C-glycosyltransferase from Trollius chinensis. Angew. Chem. Int. Ed. Engl. 2019, 58, 11513–11520. [Google Scholar] [CrossRef]
- Zhang, L.L.; Tian, K.; Tang, Z.H.; Chen, X.J.; Bian, Z.X.; Wang, Y.T.; Lu, J.J. Phytochemistry and pharmacology of Carthamus tinctorius L. Am. J. Chin. Med. 2016, 44, 197–226. [Google Scholar] [CrossRef]
- Xie, K.; Chen, R.; Li, J.; Wang, R.; Chen, D.; Dou, X.; Dai, J. Exploring the catalytic promiscuity of a new glycosyltransferase from Carthamus tinctorius. Org. Lett. 2014, 16, 4874–4877. [Google Scholar] [CrossRef] [PubMed]
- Kovinich, N.; Saleem, A.; Arnason, J.T.; Miki, B. Functional characterization of a UDP-glucose: Flavonoid 3-O-glucosyltransferase from the seed coat of black soybean (Glycine max (L.) Merr.). Phytochemistry 2010, 71, 1253–1263. [Google Scholar] [CrossRef]
- Sun, W.; Liang, L.; Meng, X.; Li, Y.; Gao, F.; Liu, X.; Wang, S.; Gao, X.; Wang, L. Biochemical and molecular characterization of a flavonoid 3-O-glycosyltransferase responsible for anthocyanins and flavonols biosynthesis in Freesia hybrida. Front. Plant Sci. 2016, 7, 410. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.L.; Wang, S.; Xu, Z.; Li, M.; Chen, K.; Zhang, Y.; Hu, Z.; Zhang, M.; Zhang, Z.; Qiao, X.; et al. Highly promiscuous flavonoid 3-O-glycosyltransferase from Scutellaria baicalensis. Org. Lett. 2019, 21, 2241–2245. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Shi, X.; Yang, C.; Zhao, X.; Zhuang, J.; Liu, Y.; Gao, L.; Xia, T. Two UDP-glycosyltransferases catalyze the biosynthesis of bitter flavonoid 7-O-neohesperidoside through sequential glycosylation in tea plants. J. Agric. Food Chem. 2022, 70, 2354–2365. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; He, J.; Chen, K.; Wang, Z.L.; Liu, J.; Qiao, X.; Ye, M. Molecular cloning and biochemical characterization of a new flavonoid glycosyltransferase from the aquatic plant lotus. Biochem. Biophys. Res. Commun. 2019, 510, 315–321. [Google Scholar] [CrossRef]
- Huang, J.; Li, J.; Yue, J.; Huang, Z.; Zhang, L.; Yao, W.; Guan, R.; Wu, J.; Liang, J.; Duan, L.; et al. Functional characterization of a novel glycosyltransferase (UGT73CD1) from Iris tectorum Maxim. for the substrate promiscuity. Mol. Biotechnol. 2021, 63, 1030–1039. [Google Scholar] [CrossRef]
- Gao, J.; Ma, B.; Lu, Y.; Zhang, Y.; Tong, Y.; Guo, S.; Gao, W.; Huang, L. Investigating the catalytic activity of glycosyltransferase on quercetin from Tripterygium wilfordii. ACS Omega 2020, 5, 1414–1421. [Google Scholar] [CrossRef]
- Jung, J.; Liu, H.; Borg, A.J.E.; Nidetzky, B. Solvent engineering for nonpolar substrate glycosylation catalyzed by the UDP-glucose-dependent glycosyltransferase UGT71E5: Intensification of the synthesis of 15-hydroxy cinmethylin β-d-glucoside. J. Agric. Food Chem. 2023, 71, 13419–13429. [Google Scholar] [CrossRef]
- Dorjjugder, N.; Taguchi, G. Production of flavonoid 7-O-glucosides by bioconversion using Escherichia coli expressing a 7-O-glucosyltransferase from Tobacco (Nicotiana tabacum). Appl. Biochem. Biotechnol. 2022, 194, 3320–3329. [Google Scholar] [CrossRef]
- Liu, X.; Li, L.; Zhao, G.R. Systems metabolic engineering of Escherichia coli coculture for de novo production of genistein. ACS Synth. Biol. 2022, 11, 1746–1757. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tian, C.; Xia, Y.; Mutanda, I.; Wang, K.; Wang, Y. Production of plant-specific flavones baicalein and scutellarein in an engineered E. coli from available phenylalanine and tyrosine. Metab. Eng. 2019, 52, 124–133. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, J.; Zhang, G.; Ding, W.; Duan, L.; Yang, J.; Kui, L.; Cheng, X.; Ruan, J.; Fan, W.; et al. Engineering yeast for the production of breviscapine by genomic analysis and synthetic biology approaches. Nat. Commun. 2018, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, Y.; Li, G.; Savolainen, O.; Chen, Y.; Nielsen, J. De novo biosynthesis of bioactive isoflavonoids by engineered yeast cell factories. Nat. Commun. 2021, 12, 6085. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mu, Y.; Chang, X.; Li, G.; Dong, Z.; Sun, J.; Jin, S.; Wang, X.; Zhang, L.; Jin, S. Functional verification and screening of protein interacting with the slPHB3. Plant Signal Behav. 2022, 1, 2025678. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).