
Efficient Biotransformation of Icariin to Baohuoside I Using Two Novel GH1 β-Glucosidases



Abstract
1. Introduction
2. Results and Discussion
2.1. Screening of 13 Active β-Glucosidases from Metagenomic Data
2.2. Purification and Enzymatic Characterization of DCF-bgl-26 and DCF-bgl-27
2.3. Bioinformatics Analysis of DCF-bgl-26 and DCF-bgl-27
2.4. Enzymatic Transformation of Icariin into Baohuoside I by DCF-bgl-26 and DCF-bgl-27
3. Materials and Methods
3.1. Strains, Plasmids, and Reagents
3.2. Screening of Novel β-Glucosidase Genes from Metagenomic Data of Cow Fecal Microbiota
3.3. Gene Expression and Catalytic Activity Determination of Candidate β-Glucosidase Genes
3.4. Expression and Purification of Active β-Glucosidases
3.5. Effects of Temperature and pH on DCF-bgl-26 and DCF-bgl-27 Activity
3.6. Enzymatic Transformation of Icariin into Baohuoside I
3.7. Bioinformatics Analyses of DCF-bgl-26 and DCF-bgl-27
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, X.; Li, Y.; Li, J.; Liang, H.; Zhang, J.; Chen, X.; Li, Q.; Pang, X.; Ding, Q.; Song, J.; et al. Bioactive metabolites reveal the therapeutic consistency of Epimedii folium from multi-plant sources for the treatment of kidney-yang deficiency. J. Ethnopharmacol. 2024, 319, 117215. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, F.; Huang, Y.; Chen, B.; Pei, C.; Huang, D.; Wang, X.; Wang, Y.; Kou, S.; Li, W.; et al. Epimedium for osteoporosis based on western and eastern medicine: An updated systematic review and meta-analysis. Front. Pharmacol. 2022, 13, 782096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.B.; Yan, Y.; He, J.; Wang, P.P.; Chen, X.; Lan, T.Y.; Guo, Y.X.; Wang, J.P.; Luo, J.; Yan, Z.R.; et al. Epimedii herba: An ancient Chinese herbal medicine in the prevention and treatment of rheumatoid arthritis. Front. Chem. 2022, 10, 1023779. [Google Scholar] [CrossRef]
- Qian, H.Q.; Wu, D.C.; Li, C.Y.; Liu, X.R.; Han, X.K.; Peng, Y.; Zhang, H.; Zhao, B.Y.; Zhao, Y. A systematic review of traditional uses, phytochemistry, pharmacology and toxicity of Epimedium koreanum Nakai. J. Ethnopharmacol. 2024, 318, 116957. [Google Scholar] [CrossRef]
- Liu, Y.M.; Liu, S.S.; Xiong, Y.Z.; Shen, Z.B.; Wang, X.; Qi, Z.C.; Zhan, H.S.; Du, G.Q. Formulation rules and main health functions of health products containing Epimedii Folium based on data mining and network pharmacology. Chin. Tradit. Herb Drugs 2024, 55, 2027–2039. [Google Scholar]
- Zhang, X.; Tang, B.; Wen, S.; Wang, Y.; Pan, C.; Qu, L.; Yin, Y.; Wei, Y. Advancements in the biotransformation and biosynthesis of the primary active flavonoids derived from Epimedium. Molecules 2022, 28, 7173. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, W.W.; Ding, B.; Guo, M.; Liang, M.; Pang, H.; Wei, Y.T.; Huang, R.B.; Du, L.Q. Highly efficient bioconversion of icariin to icaritin by whole-cell catalysis. Microb. Cell Factories 2023, 22, 64. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yang, J.; Zhao, Y.; Zheng, W.; Pang, X.; Wang, B.; Wang, J.; Li, Q.; Chen, X.G.; Zhang, J.; et al. Comprehensive analysis and quality assessment of Herba Epimedii from multiple botanical origins based on ultra-high performance supercritical fluid chromatography coupled with quadrupole time-of-flight mass spectrometry and photodiode array detector. J. Supercrit. Fluids 2019, 149, 1–9. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2020; Volume 1. [Google Scholar]
- Deng, L.; Ouyang, B.; Shi, H.; Yang, F.; Li, S.; Xie, C.; Du, W.; Hu, L.; Wei, Y.; Dong, J. Icariside II attenuates bleomycin-induced pulmonary fibrosis by modulating macrophage polarization. J. Ethnopharmacol. 2023, 317, 116810. [Google Scholar] [CrossRef]
- Wu, H.; Kim, M.; Han, J. Icariin metabolism by human intestinal microflora. Molecules 2016, 21, 1158. [Google Scholar] [CrossRef]
- Xie, J.; Xu, H.; Jiang, J.; Zhang, N.; Yang, J.; Zhao, J.; Wei, M. Characterization of a novel thermostable glucose-tolerant GH1 β-glucosidase from the hyperthermophile Ignisphaera aggregans and its application in the efficient production of baohuoside I from icariin and total Epimedium flavonoids. Bioorganic Chem. 2020, 104, 104296. [Google Scholar] [CrossRef]
- Ma, M.; Fan, A.Y.; Liu, Z.; Yang, L.Q.; Huang, J.M.; Pang, Z.Y.; Yin, F. Baohuoside I inhibits osteoclastogenesis and protects against ovariectomy-induced bone loss. Front. Pharmacol. 2022, 13, 874952. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.S.; Thakur, K.; Hu, F.; Zhang, J.G.; Wei, Z.J. Icariside II inhibits tumorigenesis via inhibiting AKT/Cyclin E/CDK 2 pathway and activating mitochondria-dependent pathway. Pharmacol. Res. 2020, 152, 104616. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Zhang, Y.; Zhang, T.; Lu, L.; Ding, Y.; Zhao, Y. Comparative pharmacokinetics study of icariin and icariside II in rats. Molecules 2015, 20, 21274–21286. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Ji, H.; Zhang, Q.W.; Tu, P.F.; Wang, Y.T.; Guo, B.L.; Li, S.P. A rapid method for simultaneous determination of 15 flavonoids in Epimedium using pressurized liquid extraction and ultra-performance liquid chromatography. J. Pharm. Biomed. Anal. 2008, 46, 226–235. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, H.; Lu, Y.; Xu, L.; Yin, H.; Tam, J.P.; Yang, H.; Jia, X.B. Construction of a novel catalysis system for clean and efficient preparation of baohuoside I from icariin based on biphase enzymatic hydrolysis. J. Clean Prod. 2018, 170, 727–734. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, M.; Zhou, J.; Chen, Y.; Xu, L.; Wu, M.; Xia, G.; Tam, J.P.; Yu, J.; Teng, X.; et al. Eco-efficient biphasic enzymatic hydrolysis for the green production of rare baohuoside I. Enzym. Microb. Technol. 2019, 131, 109431. [Google Scholar] [CrossRef]
- Zhang, X.; Miao, Q.; Pan, C.; Yin, J.; Wang, L.; Qu, L.; Yin, Y.; Wei, Y. Research advances in probiotic fermentation of Chinese herbal medicines. iMeta 2023, 2, e93. [Google Scholar] [CrossRef]
- Ouyang, B.; Wang, G.; Zhang, N.; Zuo, J.; Huang, Y.; Zhao, X. Recent advances in β-glucosidase sequence and structure engineering: A brief review. Molecules 2023, 28, 4990. [Google Scholar] [CrossRef]
- Erkanli, M.E.; El-Halabi, K.; Kim, J.R. Exploring the diversity of β-glucosidase: Classification, catalytic mechanism, molecular characteristics, kinetic models, and applications. Enzym. Microb. Technol. 2014, 173, 110363. [Google Scholar] [CrossRef]
- Yang, W.; Su, Y.; Wang, R.; Zhang, H.; Jing, H.; Meng, J.; Zhang, G.; Huang, L.; Guo, L.; Wang, J.; et al. Microbial production and applications of β-glucosidase-a review. Int. J. Biol. Macromol. 2024, 256, 127915. [Google Scholar] [CrossRef] [PubMed]
- de Giuseppe, P.O.; Souza, T.d.A.; Souza, F.H.; Zanphorlin, L.M.; Machado, C.B.; Ward, R.J.; Jorge, J.A.; Furriel, R.d.P.; Murakami, M.T. Structural basis for glucose tolerance in GH1 β-glucosidases. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zou, K.; Guo, B.; Pei, J.; Wang, Z.; Xiao, W.; Zhao, L. One-step purification and immobilization of thermostable β-glucosidase on Na-Y zeolite based on the linker and its application in the efficient production of baohuoside I from icariin. Bioorganic Chem. 2022, 121, 105690. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Yang, J.; Zhang, T.; Yang, Y.S.; Ding, Y. Optimized biotransformation of icariin into icariside II by β-glucosidase from Trichoderma viride using central composite design method. Biomed. Res. Int. 2016, 2016, 5936947. [Google Scholar] [CrossRef]
- Wang, J.; Liang, J.; Li, Y.; Tian, L.; Wei, Y. Characterization of efficient xylanases from industrial-scale pulp and paper wastewater treatment microbiota. AMB Express 2021, 11, 19. [Google Scholar] [CrossRef]
- Neun, S.; Brear, P.; Campbell, E.; Tryfona, T.; El Omari, K.; Wagner, A.; Dupree, P.; Hyvönen, M.; Hollfelder, F. Functional metagenomic screening identifies an unexpected β-glucuronidase. Nat. Chem. Biol. 2022, 18, 1096–1103. [Google Scholar] [CrossRef]
- Miao, Q.; Zhang, X.; Wang, Y.; Li, X.; Wang, Z.; Tian, L.; Qu, L.; Wei, Y. Characterization of novel pectinolytic enzymes derived from the efficient lignocellulose degradation microbiota. Biomolecules 2022, 12, 1388. [Google Scholar] [CrossRef]
- Zhang, X.; Miao, Q.; Tang, B.; Mijakovic, I.; Ji, X.J.; Qu, L.; Wei, Y. Discovery of novel alkaline-tolerant xylanases from fecal microbiota of dairy cows. Biotechnol. Biofuels Bioprod. 2023, 16, 182. [Google Scholar] [CrossRef]
- Sun, J.; Wang, W.; Ying, Y.; Hao, J. A novel glucose-tolerant GH1 β-glucosidase and improvement of its glucose tolerance using site-directed mutation. Appl. Biochem. Biotechnol. 2020, 192, 999–1015. [Google Scholar] [CrossRef]
- Liew, K.J.; Lim, L.; Woo, H.Y.; Chan, K.G.; Shamsir, M.S.; Goh, K.M. Purification and characterization of a novel GH1 beta-glucosidase from Jeotgalibacillus malaysiensis. Int. J. Biol. Macromol. 2018, 115, 1094–1102. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, Y.; Shi, P.; Ma, R.; Yang, H.; Xia, W.; Cui, Y.; Luo, H.; Bai, Y.; Yao, B. A highly glucose-tolerant GH1 β-glucosidase with greater conversion rate of soybean isoflavones in monogastric animals. J. Ind. Microbiol. Biotechnol. 2018, 45, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Kaenying, W.; Tagami, T.; Suwan, E.; Pitsanuwong, C.; Chomngam, S.; Okuyama, M.; Kongsaeree, P.; Kimura, A.; Kongsaeree, P.T. Structural and mutational analysis of glycoside hydrolase family 1 Br2 β-glucosidase derived from bovine rumen metagenome. Heliyon 2023, 9, e21923. [Google Scholar] [CrossRef] [PubMed]
- Renchinkhand, G.; Magsar, U.; Bae, H.C.; Choi, S.H.; Nam, M.S. Identification of β-glucosidase activity of Lentilactobacillus buchneri URN103L and its potential to convert ginsenoside Rb1 from Panax ginseng. Foods 2022, 11, 529. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Long, L.; Liang, M.; Li, H.; Chen, Y.; Zheng, M.; Ni, H.; Li, Q.; Zhu, Y. Characterization of a glucose-stimulated β-glucosidase from Microbulbifer sp. ALW1. Microbiol. Res. 2021, 251, 126840. [Google Scholar] [CrossRef]

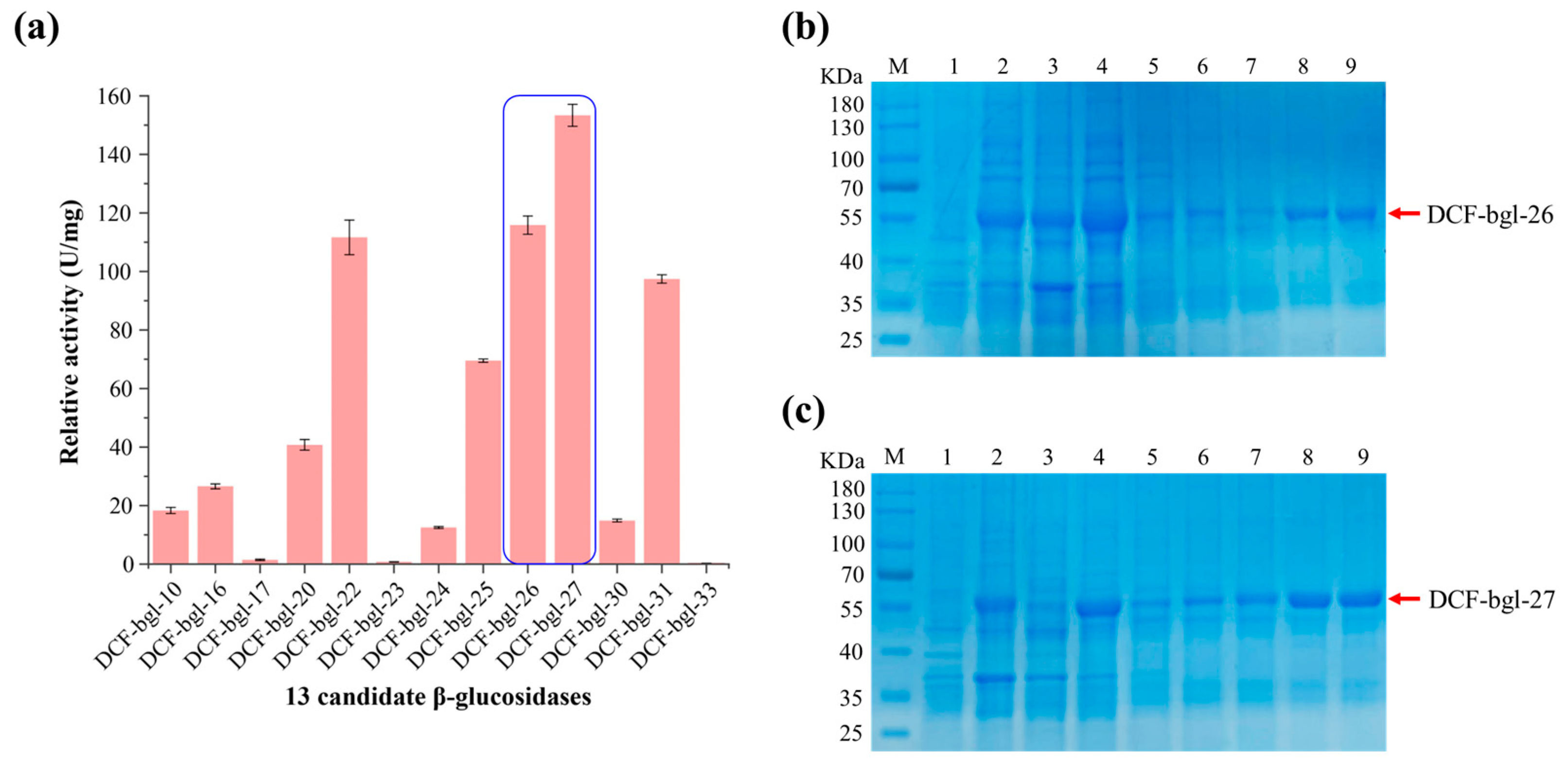
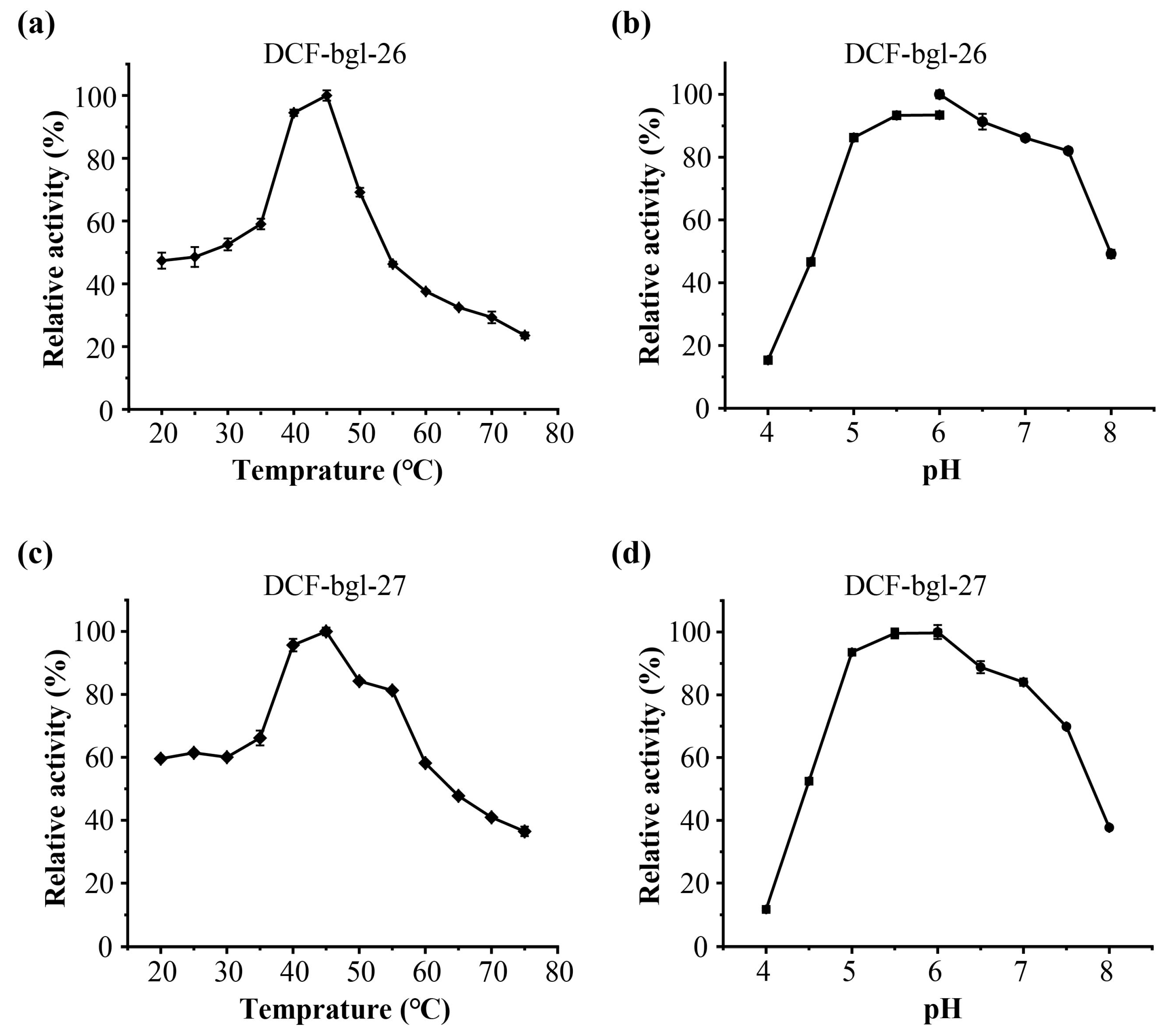
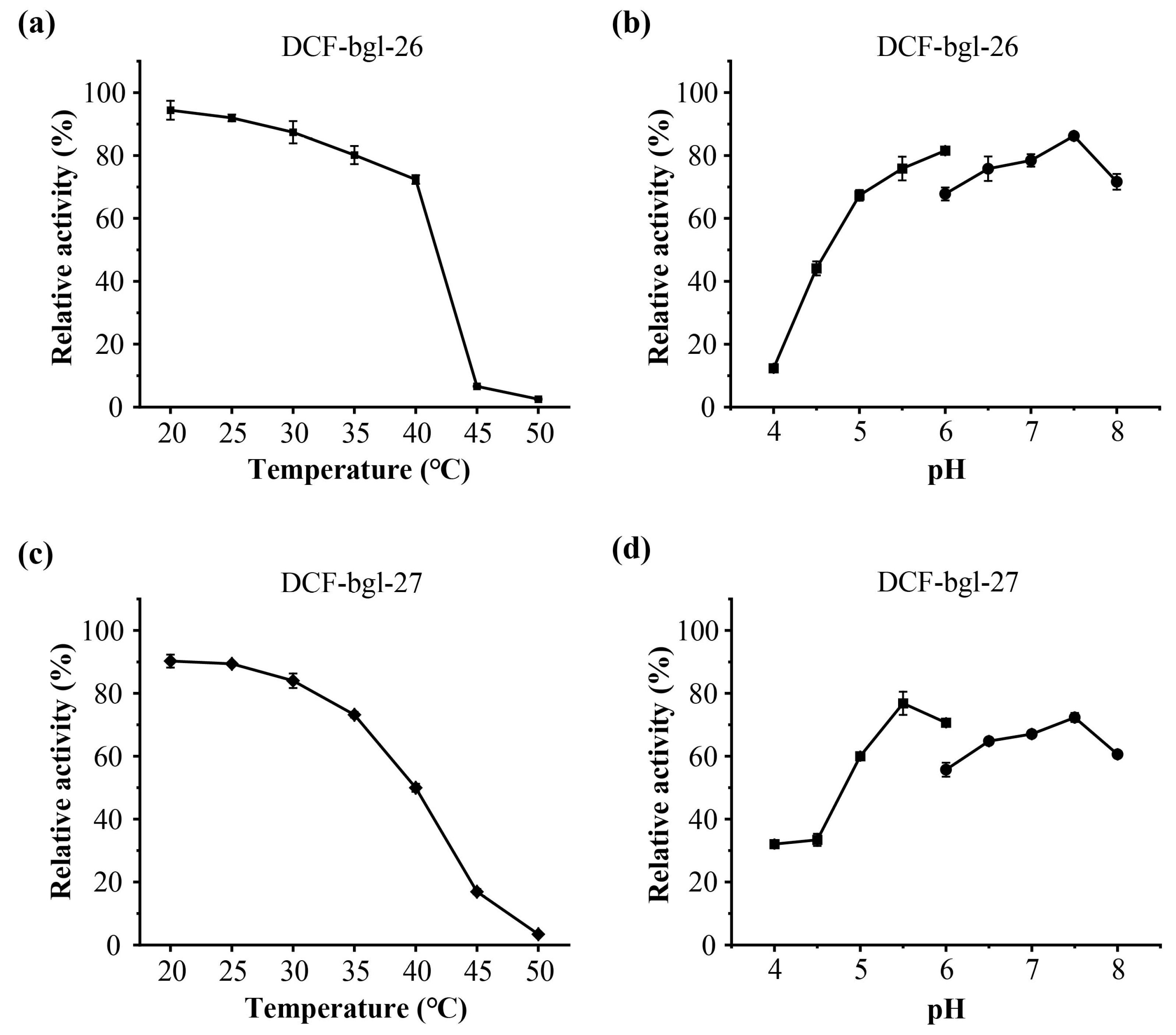

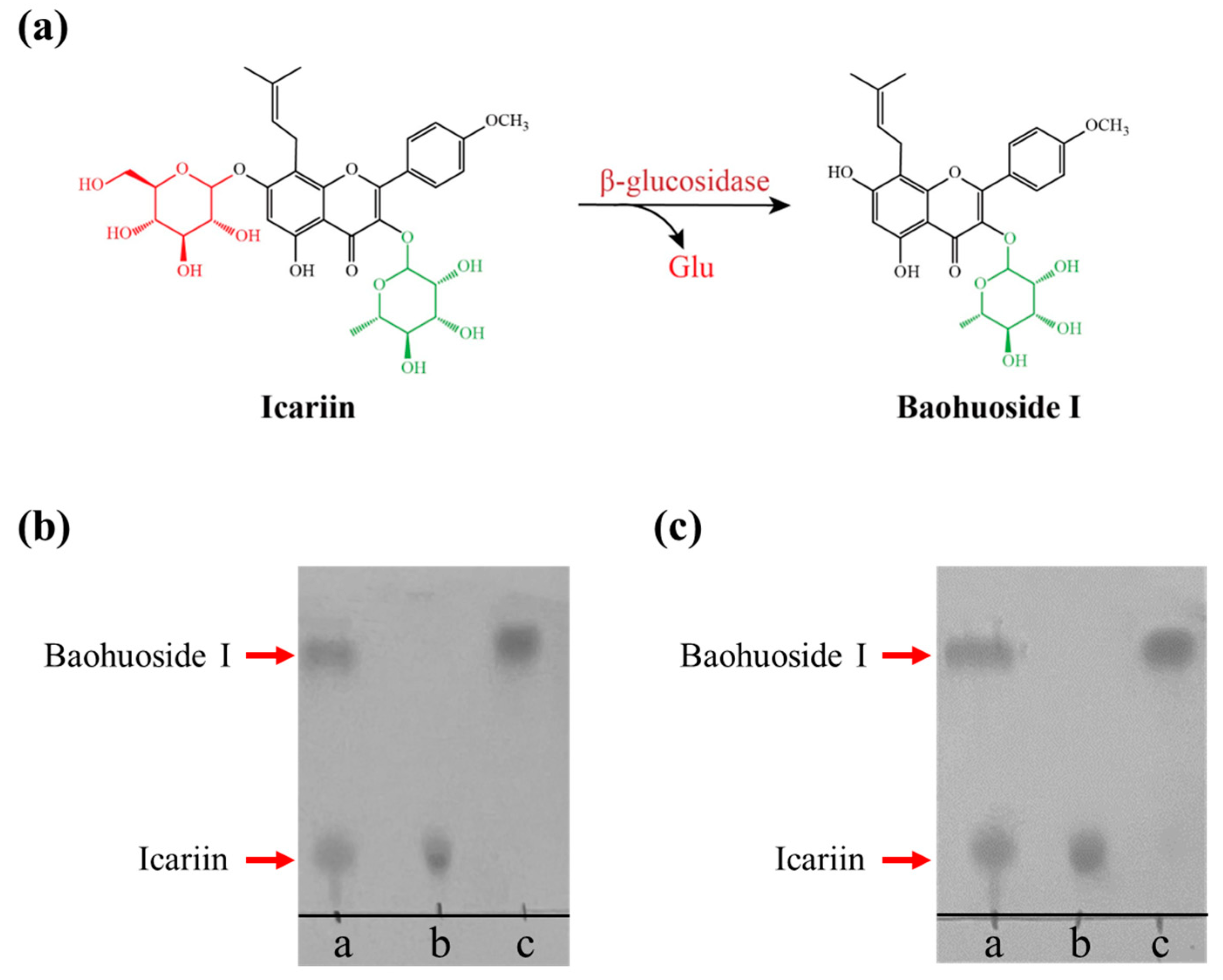

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 33 Candidate β-Glucosidase Genes | Blast Alignment Results | The Alignment Sequence ID | Identity |
|---|---|---|---|
| DCF-bgl-1 | Beta-glucosidase [Treponema sp.] | MBO5118277.1 | 95.31% |
| DCF-bgl-2 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBR5144157.1 | 85.47% |
| DCF-bgl-3 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBO5199890.1 | 69.72% |
| DCF-bgl-5 | Beta-glucosidase [Oscillospiraceae bacterium] | MBO5129153.1 | 74.28% |
| DCF-bgl-4 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBQ9780681.1 | 84.24% |
| DCF-bgl-6 | Glycoside hydrolase family 1 protein [Lentisphaeria bacterium] | MBR1999354.1 | 99.54% |
| DCF-bgl-7 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBR6574904.1 | 93.49% |
| DCF-bgl-8 | Glycoside hydrolase family 1 protein [Lentisphaeria bacterium] | MBO5681597.1 | 75.52% |
| DCF-bgl-9 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBQ8689522.1 | 80.05% |
| DCF-bgl-10 | Glycosyl hydrolase family protein [Clostridiales bacterium] | MBE7090053.1 | 94.04% |
| DCF-bgl-11 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBO5758315.1 | 96.82% |
| DCF-bgl-12 | Glycosyl hydrolase family protein [Clostridiales bacterium] | MBE7081073.1 | 94.56% |
| DCF-bgl-13 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBR5124729.1 | 93.20% |
| DCF-bgl-14 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBP3571607.1 | 93.03% |
| DCF-bgl-15 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBQ7909166.1 | 91.46% |
| DCF-bgl-16 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBR1974365.1 | 87.39% |
| DCF-bgl-17 | GH1 family beta-glucosidase [Psychrobacillus psychrotoler] | WP_093537721.1 | 99.55% |
| DCF-bgl-18 | Family 1 glycosyl hydrolase [Clostridia bacterium] | MBQ3125168.1 | 88.14% |
| DCF-bgl-19 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBQ1272506.1 | 68.23% |
| DCF-bgl-20 | Glycoside hydrolase family 1 protein [Clostridia bacterium] | MBQ4323209.1 | 98.37% |
| DCF-bgl-21 | Beta-glucosidase [Clostridia bacterium] | MBO7345591.1 | 68.15% |
| DCF-bgl-22 | Beta-glucosidase [Clostridia bacterium] | MBQ8740605.1 | 72.28% |
| DCF-bgl-23 | Beta-glucosidase [Oscillospiraceae bacterium] | MBR3978613.1 | 72.22% |
| DCF-bgl-24 | Beta-glucosidase [Oscillospiraceae bacterium] | MBR3978613.1 | 71.78% |
| DCF-bgl-25 | Beta-glucosidase [Clostridia bacterium] | MBQ9846934.1 | 99.78% |
| DCF-bgl-26 | Beta-glucosidase [Clostridia bacterium] | MBQ1284466.1 | 99.78% |
| DCF-bgl-27 | Beta-glucosidase [Clostridia bacterium] | MBQ9846934.1 | 97.57% |
| DCF-bgl-28 | Beta-glucosidase [Enterococcus casseliflavus] | VTT36984.1 | 75.32% |
| DCF-bgl-29 | Glycoside hydrolase family 1 protein [Turicibacter sp.] | MBQ1787025.1 | 99.57% |
| DCF-bgl-30 | GH1 family beta-glucosidase [Sarcina sp. DSM 11001] | WP_092994021.1 | 99.35% |
| DCF-bgl-31 | Family 1 glycosyl hydrolase [Lachnospiraceae bacterium] | MBR3186943.1 | 86.30% |
| DCF-bgl-32 | Beta-glucosidase [Oscillospiraceae bacterium] | MBE6980119.1 | 77.17% |
| DCF-bgl-33 | Beta-glucosidase [Roseburia sp.] | HAT88334.1 | 74.84% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, Y.; Zhang, T.; Yuan, Z.; Wei, Y. Efficient Biotransformation of Icariin to Baohuoside I Using Two Novel GH1 β-Glucosidases. Molecules 2024, 29, 5280. https://doi.org/10.3390/molecules29225280
Zhang X, Wang Y, Zhang T, Yuan Z, Wei Y. Efficient Biotransformation of Icariin to Baohuoside I Using Two Novel GH1 β-Glucosidases. Molecules. 2024; 29(22):5280. https://doi.org/10.3390/molecules29225280
Chicago/Turabian StyleZhang, Xiaoling, Yitong Wang, Tiantian Zhang, Ziqiao Yuan, and Yongjun Wei. 2024. "Efficient Biotransformation of Icariin to Baohuoside I Using Two Novel GH1 β-Glucosidases" Molecules 29, no. 22: 5280. https://doi.org/10.3390/molecules29225280
APA StyleZhang, X., Wang, Y., Zhang, T., Yuan, Z., & Wei, Y. (2024). Efficient Biotransformation of Icariin to Baohuoside I Using Two Novel GH1 β-Glucosidases. Molecules, 29(22), 5280. https://doi.org/10.3390/molecules29225280