Abstract
Taking the endophytic fungus UJ3-2, isolated from Urtica fissa, as the experimental material, this study aimed to explore the composition of its metabolites and the underlying mechanisms by which it inhibits Staphylococcus aureus. Initially, the MIC, MBC, inhibitory curves, biofilm growth, and extracellular nucleic acids and proteins of S. aureus in response to the metabolites were measured. Secondly, PI staining and SEM were used to evaluate the impact of the metabolites on the integrity of the cell wall and overall morphology of S. aureus. Additionally, UPLC-MS was employed to analyze the composition of the secondary metabolites. The UJ3-2 strain was identified as Xylaria grammica based on ITS sequencing and designated as Xylaria grammica UJ3-2. Our results revealed that the metabolites of UJ3-2 exhibited excellent in vitro antibacterial activity against S. aureus, with both MIC and MBC values of 3.125 mg/mL. The inhibitory curve confirmed that 1 MIC of UJ3-2 metabolites could completely inhibit the growth of S. aureus within 24 h. With increasing concentrations of UJ3-2 metabolites, the growth of S. aureus biofilms was significantly suppressed, and obvious leakage of nucleic acids and proteins was observed. PI fluorescence staining indicated that various concentrations of UJ3-2 metabolites disrupted the integrity of the S. aureus cell membrane. SEM observation revealed that the treated S. aureus surfaces became rough, and the bacteria shrank and adhered to each other, showing a dose-dependent effect. UPLC-MS analysis suggested that the main components of the fermented metabolites were 6-oxocineole (17.92%), (S)-2-acetolactate (9.91%), 3-methyl-cis,cis-muconate (4.36%), and 8-oxogeranial (3.17%). This study demonstrates that the endophytic fungus UJ3-2 exhibits remarkable in vitro antibacterial effects against S. aureus, primarily by enhancing the permeability of the S. aureus cell membrane, causing the leakage of its intracellular contents, and altering the bacterial surface morphology to inhibit the pathogen. The endophytic fungus UJ3-2 has a good antibacterial effect on S. aureus, which gives it certain application prospects in the screening and industrial production of new and efficient natural antibacterial active substances.
1. Introduction
Staphylococcus aureus (S. aureus) continues to be one of the most involved bacteria in animal and human diseases. This bacterium is found in the normal skin microbiota of both animals and humans, with a carriage rate of between 20 and 30% in the healthy human population [1]. Antibiotic resistance develops quickly in S. aureus, and the rise in multidrug-resistant forms is a major problem [2]. It has been reported that the annual mortality toll from antibiotic-resistant diseases has surpassed 10 million and that by 2050, it will outnumber cancer deaths [3]. Methicillin-resistant S. aureus (MRSA) represents a prevalent cause of multidrug-resistant infections, resulting in considerable morbidity and mortality [4]. The clinical manifestations and predisposing factors for infection differ among HA-MRSA, CA-MRSA, and LA-MRSA strains [5]. The morbidity and mortality consequences reinforce the need to urgently discover new effective solutions due to the inefficiency of traditional antibiotics. Therefore, alternative treatments represent a promising field of investigation due to the lack of new antibiotic classes.
Endophytes refer to bacteria or fungi that inhabit plant organs, are able to colonize within plant tissues, and do not cause significant harm to the host [6]. Plant–endophyte associations represent an inexhaustible source of novel metabolites, exhibiting significance from environmental, agricultural, and pharmaceutical perspectives [7,8]. Endophytic fungi are an important reservoir of therapeutically active compounds [9]. Hydrolytic enzymes from Amazonian endophytic fungi Myrcia guianensis effectively eradicate S. aureus biofilms, with combinations of lipase and protease showing total degradation, highlighting their potential as a tool against microbial biofilms [10]. Penicillium brevicompactum ANT13 from Abies numidica is a promising source of novel antimicrobials, effectively inhibiting multidrug-resistant S. aureus and dermatophytes, with ethyl acetate extract exhibiting significant activity [11]. A new nucleoside, cyclohexenone, and benzofuranone derivatives from Aspergillus polyporicola R2, with compounds 11-dehydrocurvularin, 11α-hydroxycurvularin, and 2,5-dimethyl-3(2H)-benzofuranone, exhibit antimicrobial activity against MRSA, Salmonella, and Fusarium graminearum, respectively [12].
Faced with the increasingly severe challenge of antibiotic resistance, this global issue has posed a significant threat to human health that cannot be ignored. Therefore, actively exploring and deeply mining the resources of endophytic fungi in plants has become one of the most important ways for us to find and develop new and efficient antibacterial drugs [13,14]. Based on the previously screened endophytic fungus UJ3-2 from Urtica fissa, with antibacterial activity against S. aureus, Escherichia coli, and Salmonella, using tetracycline as a positive drug, it was found that UJ3-2 exhibited the best antibacterial effect against S. aureus, but its antibacterial components and mechanisms need further research. Therefore, this experiment aims to explore the antibacterial components and their mechanisms of action in the metabolites of the endophytic fungus UJ3-2 by determining its antibacterial activity against S. aureus and its effects on cell wall and membrane integrity, biofilm, and bacterial structure. It is expected to provide practical evidence for the study of the antibacterial mechanism of pathogens and a theoretical basis for screening novel and natural antibacterial active substances.
2. Result
2.1. Identification of Endophytic Fungus UJ3-2
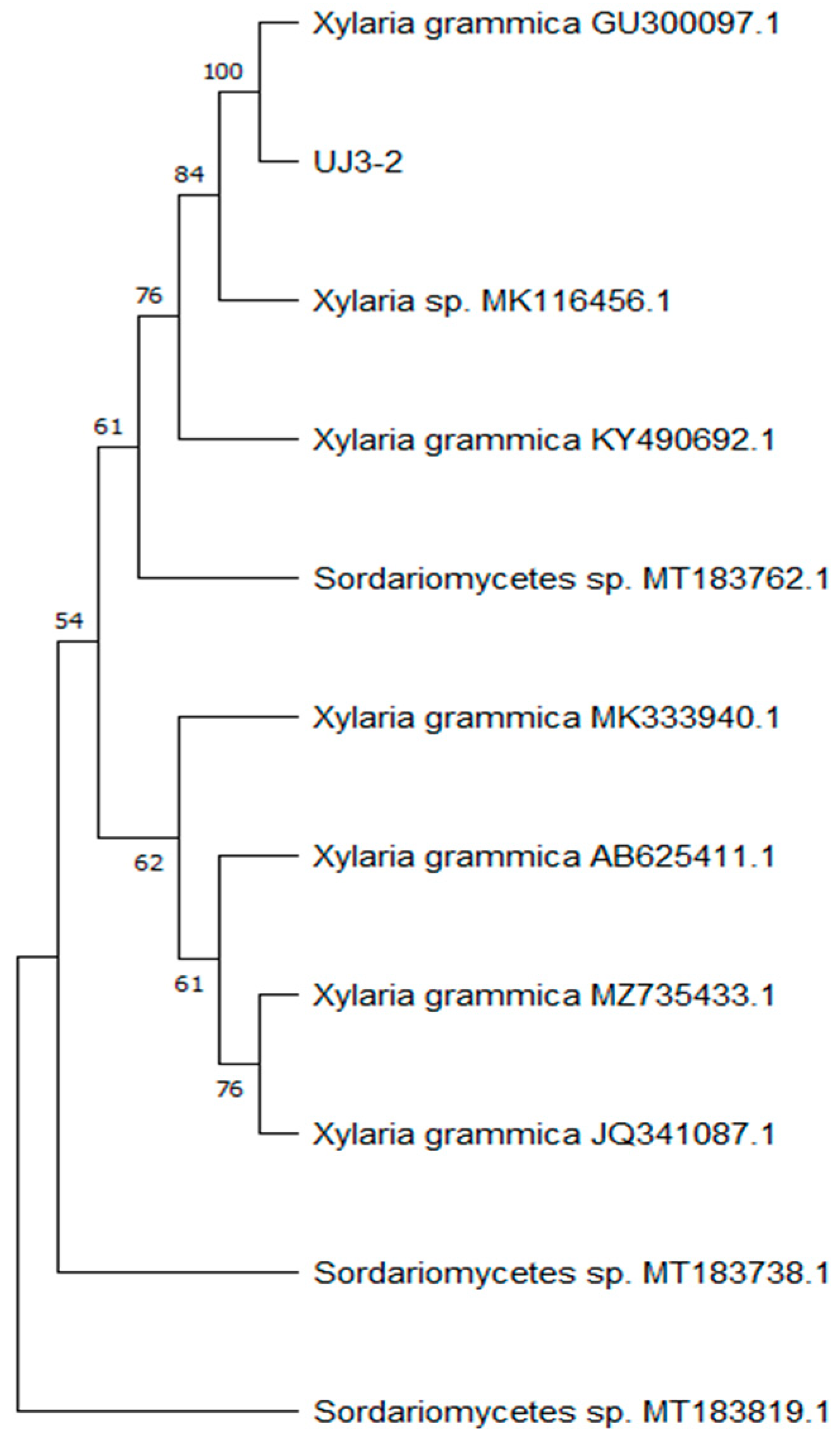
After extracting the fungal genomic DNA, the ITS sequence was amplified, and the amplified PCR products were sent for sequencing. A phylogenetic tree of endophytic fungi was constructed, as shown in Figure 1, which indicated 100% homology with Xylaria grammica, thus identifying it as Xylaria grammica. Meanwhile, the sequencing data were uploaded to the NCBI database under sequence number PP957465.1.
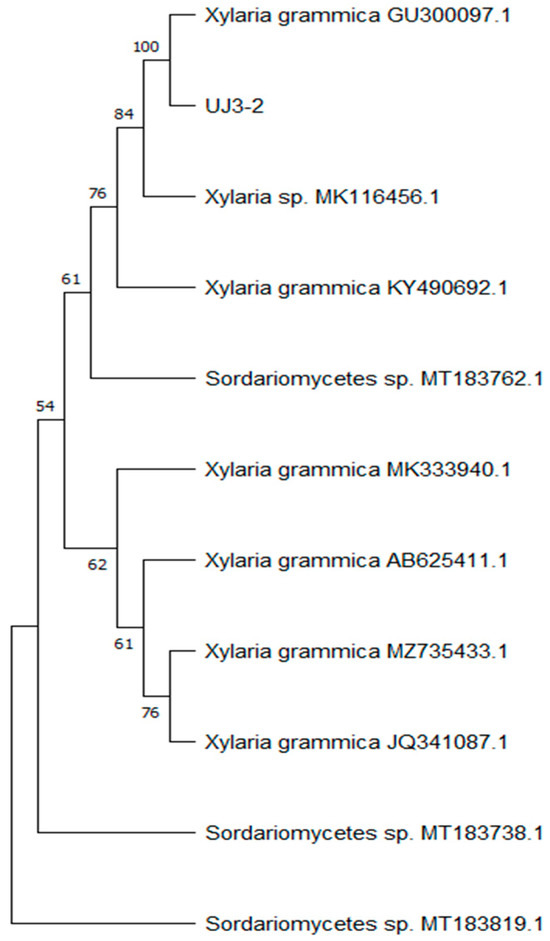
Figure 1.
Phylogenetic tree based on ITS rDNA sequence.
2.2. Metabolome Analysis of UJ3-2 Fermentation Products
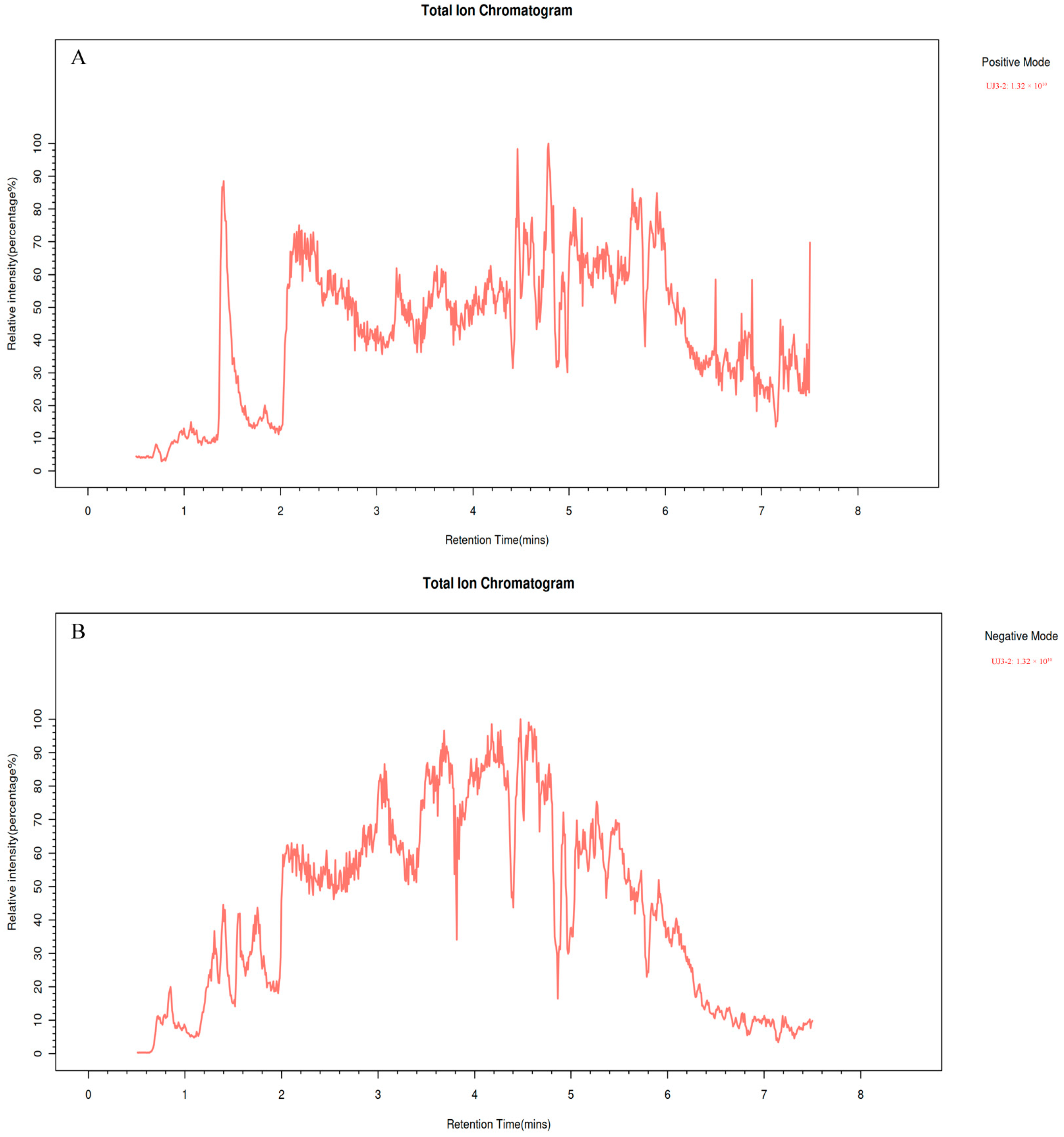
UPLC-MS/MS was used for identification of the fermentation product of UJ3-2 (Figure 2). The identified compounds mainly included low-molecular-weight phenolic compounds, alkaloids, flavonoids, organic acids, amino acids, sugars, lipids, sugar alcohols, etc. Among them, 6-oxocineole (17.92%), (s)-2-acetolactate (9.91%), 3-methyl-cis,cis-muconate (4.36%), and 8-oxogeranial (3.17%) had the highest concentrations (Table 1). Compounds such as solasodine, alpha-curcumene, bornyl isovalerate, 4-hydroxybenzaldehyde, and naringenin exhibit antibacterial activity, while aesculetin exhibits anti-inflammatory and antitumor activities. Additionally, 8-oxogeranial serves as a substrate for the synthesis of the basic chemical structure (iridoid ring scaffold) of iridoid compounds.
Figure 2.
Total ion chromatogram, TIC. (A) Positive mode; (B) Negative mode.

Table 1.
Main components of fermentation product of UJ3-2.
2.3. MIC and MBC of Fermentation Product of UJ3-2 against S. aureus
As can be seen from Table 2, for S.aureus, when the concentration of UJ3-2 fermentation product is 1.563 mg/mL, the liquid medium is cloudy, indicating that there is significant growth of S. aureus. However, when the mass concentration is 3.125 mg/mL, the culture medium is clear, indicating no significant growth of the strain. Therefore, the MIC of the UJ3-2 fermentation product against this indicator bacterium can be determined to be 3.125 mg/mL. According to Table 2, the MBC of the UJ3-2 fermentation product against this indicator bacterium can also be determined to be 3.125 mg/mL.
Table 2.
MIC and MBC of UJ3-2 fermentation product against S. aureus.
2.4. Effects of UJ3-2 Fermentation Products on Growth of S. aureus
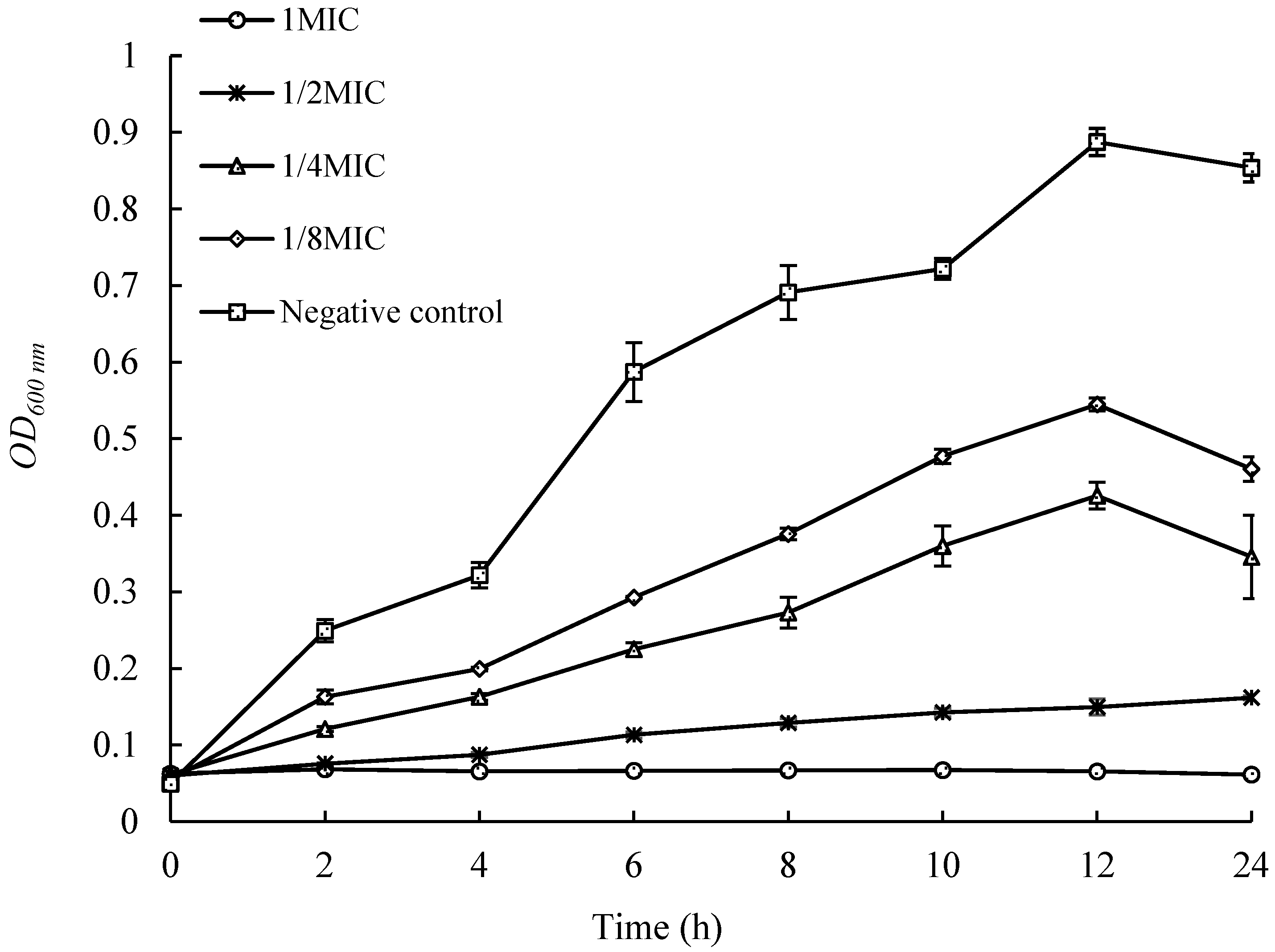
As shown in Figure 3, the control group strain grew rapidly within 12 h, reaching an OD600nm of 0.887 at 12 h. The growth of strains with the addition of different concentrations of extracts was significantly inhibited, and their OD values, at the same time, were significantly lower than those of the control group. When the concentration of the added extract reached 1 MIC, the growth of the strain was almost completely inhibited, with no significant change in OD600nm within 24 h. This indicates that when the concentration of the extract from UJ3-2 metabolites reaches 1 MIC, it can effectively inhibit the growth and reproduction of S. aureus strains in liquid culture.
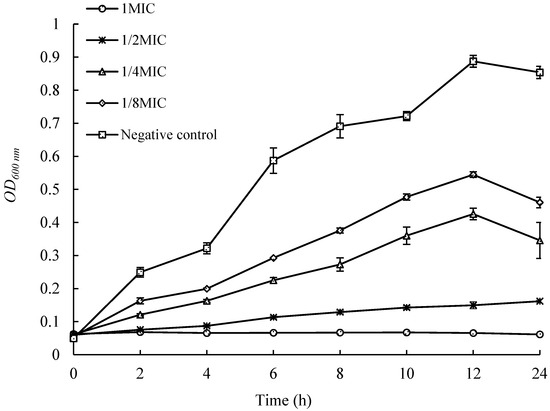
Figure 3.
Effects of UJ3-2 fermentation products on growth of S. aureus.
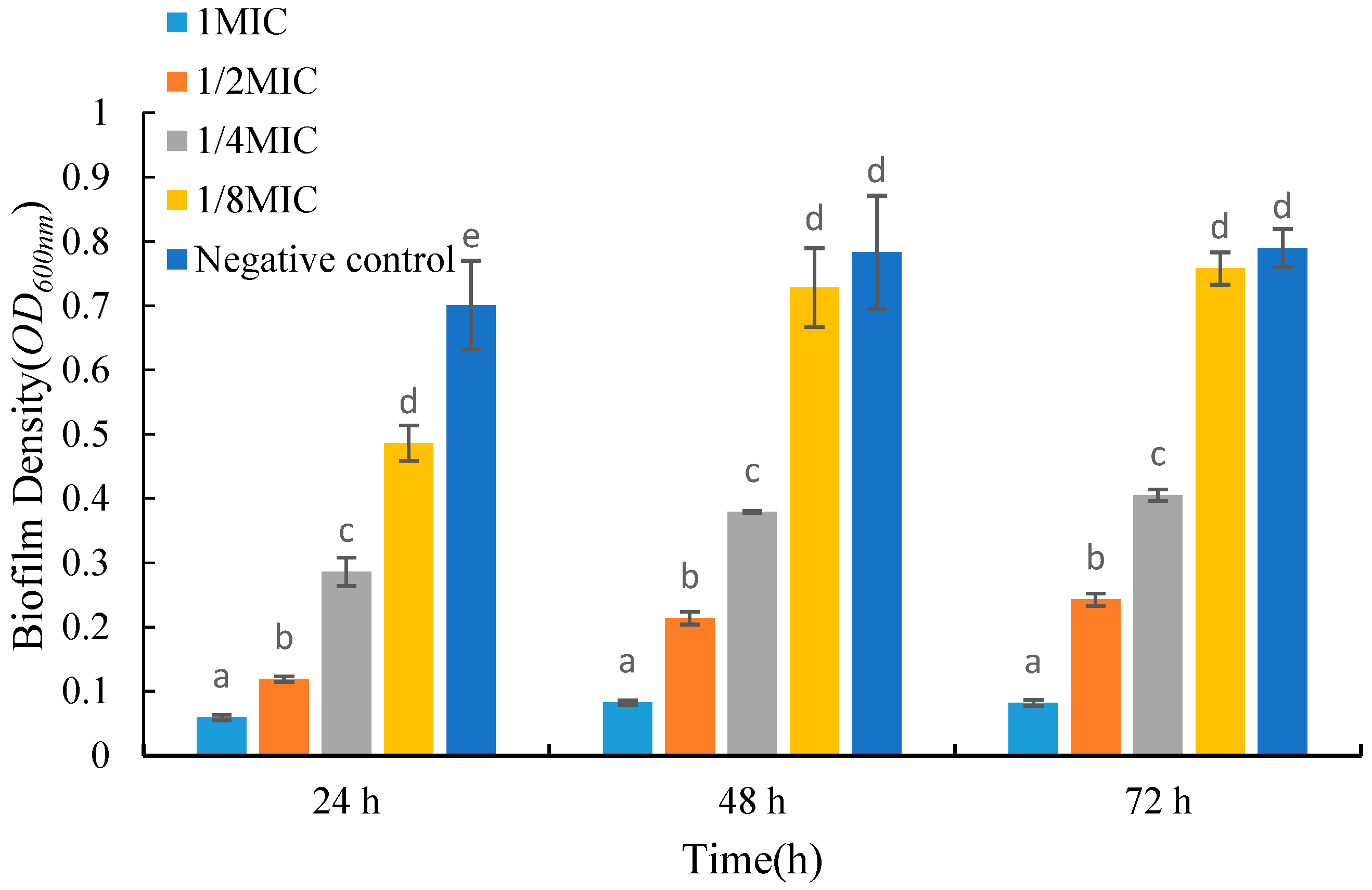
2.5. Effects of UJ3-2 Fermentation Products on Growth of S. aureus Biofilm
As shown in Figure 4, the biomass of biofilms decreased significantly after adding the fermentation product to the medium. For biofilms grown in the same period, the biomass gradually decreased with an increase in the concentration of the fermentation product. Compared with the control group, the absorbance values of 24 h old biofilms treated with low and high concentrations of fermentation products decreased by 30.7% and 91.5%, respectively. After 72 h of growth, the absorbance of biofilms treated with low and high concentrations of fermentation products decreased by 4.01% and 89.6%, respectively. From Figure 4, it can be seen that when treated with the same concentration of fermentation product, the biomass of 48 h old biofilms is larger than that of 24 h old biofilms, but there is no significant trend in the biomass of 72 h old biofilms compared to 48 h old biofilms.
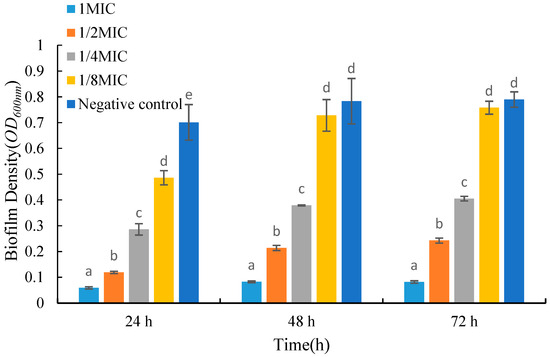
Figure 4.
Effects of UJ3-2 fermentation products on growth of S. aureus biofilm. At the same time point, different letters represented significant difference (p < 0.05).
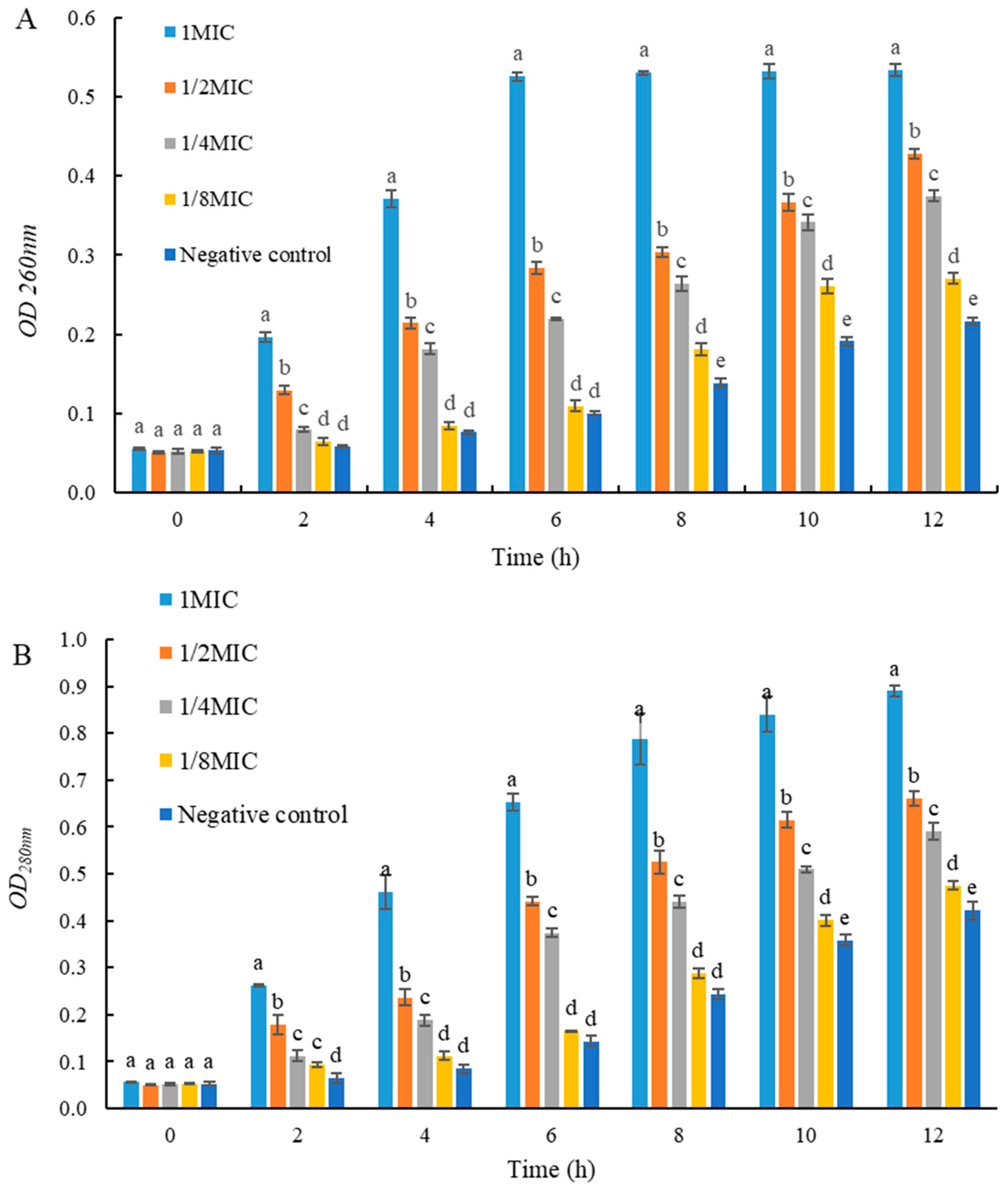
2.6. Effects of UJ3-2 Fermentation Products on Extracellular Nucleic Acid and Protein of S. aureus
As shown in Figure 5, the concentrations of extracellular nucleic acids and proteins in S. aureus in the 1 MIC and 2 MIC groups were significantly higher than those in the control group (p < 0.05). This result indicates that the fermentation product of endophytic fungus UJ3-2 can disrupt the cell membrane structure of S. aureus, causing the leakage of intracellular nucleic acids and proteins from the bacteria.
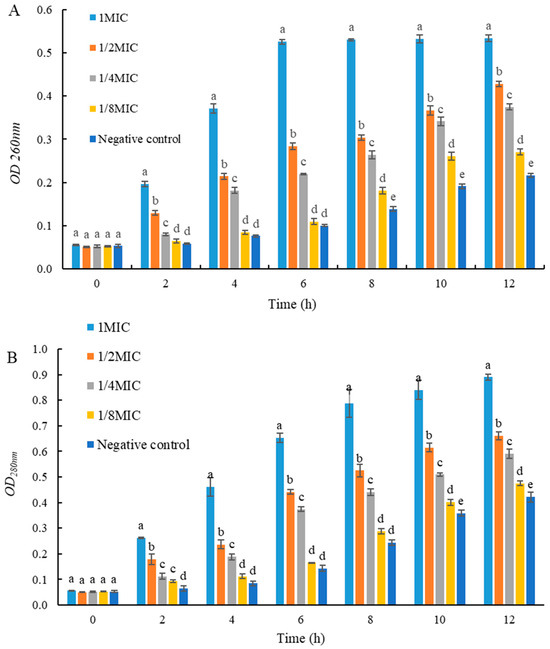
Figure 5.
Effects of UJ3-2 fermentation products on extracellular nucleic acid and protein of S. aureus. (A) OD260nm; (B) OD280nm. At the same time point, different letters represented significant difference (p < 0.05).
2.7. Effects of UJ3-2 Fermentation Products on Cell Membrane Integrity of S. aureus
Propidium iodide (PI), as a membrane-impermeable fluorescent dye, possesses a unique mechanism of action [15]. When the integrity of the cell membrane is compromised, it can penetrate the membrane, bind to genetic material, and emit red fluorescence. Therefore, the integrity of the cell membrane can be reflected by detecting PI fluorescence through fluorescence microscopy. As shown in Figure 6, after PI staining, the control group exhibited virtually no observable fluorescence, while the S. aureus group treated with 0.5 MIC metabolites for 4 h emitted sporadic red fluorescence. With an increase in extract concentration, the red fluorescence gradually intensified. When the mass concentration reached 2 MIC, a large amount of red fluorescence was observed, indicating that after treatment with the extract, the cell membrane of S. aureus was disrupted, allowing a significant amount of PI to enter the cells and bind to genetic material, resulting in the emission of red fluorescence.

Figure 6.
Effects of different concentrations of UJ3-2 fermentation product on cell membrane integrity of S. aureus (fluorescence microscope). Note: (A) negative control group ((A1) bright field, (A2) dark field); (B) 1/4 MIC group ((B1) bright field, (B2) dark field); (C) 1/2 MIC group ((C1) bright field, (C2) dark field); (D) 1 MIC group ((D1) bright field, (D2) dark field).
2.8. Effect of Fermentation Products on S. aureus Was Observed by Scanning Electron Microscope
Observations from scanning electron microscopy revealed that compared to the control group (Figure 7A,B), S. aureus treated with the fermentation product of UJ3-2 exhibited bacterial morphological deformations, including blurred edges and rough surfaces. This indicates that the fermentation product of UJ3-2 can affect the overall morphology of S. aureus and alter its bacterial surface structure.

Figure 7.
Effect of 1/2 MIC of UJ3-2 fermentation product on morphology of S. aureus cells. Note: (A,B) normal controls; (C,D) processing group.
3. Discussion
The secondary metabolites of microorganisms, as natural antibacterial substances, are important sources for the development and creation of new drugs [16]. Endophytic fungi in plants are highly diverse groups in microbial classification [17]. These fungi can synthesize medicinal substances. Therefore, they may be excellent sources of new antibacterial compounds for the treatment of human and animal pathogen infections [18]. However, the study of these microbial resources is boundless and requires the extensive attention of microbiologists and medicinal chemists. In this study, an endophytic fungus, UJ3-2, was isolated from fresh plants of Urtica fissa (U. fissa). By means of ITS sequencing and alignment, it was identified as Xylaria grammica fungus. U. fissa is commonly used to treat rheumatism, arthritis, and urticaria. Many chemical compounds with anti-inflammatory properties have recently been identified [19,20,21,22,23]. Fungi belonging to the genus Xylaria are concentrated in temperate, subtropical, and tropical regions, and their main chemical components are secondary metabolites such as terpenes, alkaloids, sterols, and polyketides, which possess a variety of pharmacological activities, including antioxidation, antibiosis, antitumor activity, etc. [24]. Xylaria that symbiotizes with plants can also produce the same or similar physiologically active substances as its host plants [25]. An endolichenic fungus, Xylaria grammica strain EL000614, exhibited potent nematicidal activity against the plant-pathogenic nematode Meloidogyne incognita through the production of grammicin [26,27].
The strain UJ3-2 obtained in this study exhibited significant inhibitory effects on S. aureus. The MIC of the extract from its fermentation broth against S. aureus was 3.125 mg/mL. When the concentration of the extract was 1/8 MIC, it significantly inhibited the growth of S. aureus, with a notably lower OD600 value compared to the control group at the same time point. At a concentration of 1 MIC, it completely inhibited the growth of S. aureus. When the extract concentration was 1/4 MIC, it significantly inhibited the biofilm formation of S. aureus. To investigate the effect of the UJ3-2 fermentation broth on the cell membrane integrity of S. aureus, the bacteria were stained with PI and observed under a fluorescence microscope. PI cannot penetrate intact cell membranes, but when the cell membrane is damaged, PI can enter the cell, stain the DNA, and emit red fluorescence [28]. As the concentration of the fermentation broth increased, the damage to the cell membrane of S. aureus became more severe. When the concentration of the fermentation broth reached 1 MIC, the cell membrane of S. aureus was severely damaged, resulting in a significant number of bacterial deaths. Additionally, scanning electron microscopy revealed that S. aureus treated with the fermentation product of UJ3-2 exhibited morphological deformations, including blurred edges and rough surfaces, indicating that the fermentation product of UJ3-2 can affect the overall morphology and alter the bacterial surface structure of S. aureus. Microbial biofilm is an aggregate of microbial cells embedded in the extracellular polymeric substance (EPS) matrix produced by themselves. Biofilm is resistant to extreme environments and can protect microorganisms from ultraviolet (UV) radiation, extreme temperatures, extreme pH values, high salinity, high pressure, malnutrition, antibiotics, etc., by acting as an “aprotective suit” [29]. Therefore, it is speculated that the antibacterial mechanism of the UJ3-2 fermentation broth against S. aureus may be attributed to the increased concentration of the fermentation broth, which leads to enhanced permeability of the cell membrane of S. aureus, damaging the integrity of the cell membrane. This results in the influx of a large number of exogenous substances and the leakage of cytoplasm, causing intracellular osmotic pressure disorders that prevent normal growth and reproduction of the bacteria.
UPLC-MS was used to analyze and identify the components of the UJ3-2 fermentation broth extract. The identification results showed that the UJ3-2 fermentation broth extract mainly contained compounds such as organic acids, flavonoids, phenols, coumarins, and alkaloids. The content of 6-oxocineole (17.92%), (S)-2-acetolactate (9.91%), 3-methyl-cis,cis-muconate (4.36%), and 8-oxogeranial (3.17%) was the highest. 6-oxocineole is a cineole. Cineole, a terpene oxide inherent in a myriad of plant essential oils, boasts notable anti-inflammatory, analgesic, and antioxidant properties [30,31]. 8-Oxogeranial serves as a precursor in the biosynthesis of iridoids. Iridoid synthases (IRIS), members of the PRISE subclass within the SDR protein family, orchestrate the pivotal step in iridoid formation: the cyclization of 8-oxogeranial into nepetalactol, the ubiquitous biosynthetic precursor for all iridoids [32]. The fundamental chemical architecture of iridoids in plants, namely the iridoid ring scaffold, is biologically synthesized within the plant by the enzyme iridoid synthase, employing 8-oxogeranial as its primary substrate [33]. Iridoids exhibit a promising capacity to modulate numerous biological processes in a wide array of diseases [34]. Among them, compounds such as solasodine, alpha-Curcumene, Bornyl isovalerate, 4-hydroxybenzaldehyde, and naringenin exhibit antibacterial activity. Da Silva has shown that solasodine may be acting on the mechanism of action of the antibiotic, as it has shown a potentiating effect when associated with antibiotics, inducing a reduction in the MIC against Gram-positive and Gram-negative bacteria [35]. Solasodine can be used as a potent anti-mycobacterial agent for combating Paratuberculosis [36]. Zhang et al. have shown that ginger essential oil can disrupt the cell membrane and biofilm structure of Staphylococcus, demonstrating its potential as a natural food preservative. Among its components, alpha-curcumene is the main active compound of ginger essential oil [37]. 4-hydroxybenzaldehyde sensitizes Acinetobacter baumannii to amphenicols [38]. Naringenin has antidiabetic, anticancer, antimicrobial, antiobesity, gastroprotective, immunomodulator, cardioprotective, nephroprotective, and neuroprotective properties [39,40]. This indicates that the components of the UJ3-2 fermentation extract are complex and abundant, exerting an antibacterial effect through multiple components.
4. Materials and Methods
4.1. Materials
The endophytic fungus UJ3-2 of Urtica fissa was previously isolated by our laboratory; the strain of S. aureus (CMCC(B)26003) was purchased from Shanghai Luwei Technology Co., Ltd. (Shanghai, China); Casein Hydrolysate (MH) broth medium and LB agar medium were provided by Qingdao Haibo Biotechnology Co., Ltd. (Qingdao, China); propidium iodide (PI) was obtained from Solarbio (Beijing, China); and anhydrous ethanol, ethyl acetate, ether, and n-butanol (analytical pure) were obtained from Aladdin (Shanghai, China).
4.2. Methods
4.2.1. Identification of Endophytic Fungi
The identification of isolated endophytic fungi was carried out using the ITS (Internal Transcribed Spacer) sequence of Urtica fissa. Universal fungal primers were employed to amplify the ITS region [41]: ITS1: 5′-TCCGTAGGTGAACCTGCGG-3′, and ITS4: 5′-TCCTCCGCTTATTGATATGC-3′. The resulting ITS sequences were then submitted to the GeneBank database, and BLAST on the NCBI website was used to search for the DNA sequence with the highest similarity. Together with the sequencing results, the software Mega11 version 11.0.11 was utilized to construct an evolutionary tree using the neighbor-joining (NJ) method, in order to identify the generic classification of each endophytic fungal strain.
4.2.2. Preparation of UJ3-2 Fermentation Products
The fungus was inoculated in a conical flask containing Potato Dextrose Broth and cultured in a constant-temperature shaker for 15 days (28 °C, 120 r/min) to obtain a fermented broth for later use. The fermented broth was centrifuged (4 °C, 8000 r/min) for 10 min, and the supernatant was retained. On a sterile working bench, the supernatant was filtered through a 0.22 μm filter membrane to obtain a sterile fermented broth. The sterile fermented broth was then extracted with an equal volume of ethyl acetate for 30 min each time, and this was repeated three times. The combined extracts were concentrated using a rotary evaporator under reduced pressure and then dried in a vacuum oven to obtain the fermented metabolites of UJ3-2.
4.2.3. Ultra-High-Performance Liquid Chromatography–Mass Spectrometry (UPLC-MS)
Chromatographic Separation: A Thermo Vanquish (Thermo Fisher Scientific, Waltham, MA, USA) ultra-high-performance liquid chromatography (UHPLC) system was employed, utilizing an ACQUITY UPLC® HSS T3 (2.1 × 100 mm, 1.8 µm) column (Waters, Milford, MA, USA) with a flow rate of 0.3 mL/min and a column temperature set at 40 °C. A sample volume of 2 μL was injected. For the positive ion mode, the mobile phases consisted of 0.1% formic acid in acetonitrile (B2) and 0.1% formic acid in water (A2), with the following gradient elution program: 0–1 min, 10% B2; 1–5 min, 10%–98% B2; 5–6.5 min, 98% B2; 6.5–6.6 min, 98%–10% B2; 6.6–8 min, 10% B2. For the negative ion mode, the mobile phases were acetonitrile (B3) and 5 mM ammonium formate in water (A3), with the following gradient elution program: 0–1 min, 10% B3; 1–5 min, 10%–98% B3; 5–6.5 min, 98% B3; 6.5–6.6 min, 98%–10% B3; 6.6–8 min, 10% B3.
Mass Spectrometry Acquisition: A Thermo Q Exactive Focus mass spectrometer (Thermo Fisher Scientific, USA), equipped with an Electrospray Ionization (ESI) source, was used to acquire data in both positive and negative ion modes. The spray voltages were set at 3.50 kV for positive ions and −2.50 kV for negative ions. The sheath gas flow rate was 40 arb, and the auxiliary gas flow rate was 10 arb. The capillary temperature was maintained at 325 °C. A first-order full scan was performed at a resolution of 70,000, with an ion scan range of m/z 100–1000. Higher-energy collisional dissociation (HCD) was employed for second-order fragmentation, with a collision energy of 30 eV and a second-order resolution of 17,500. The top 3 most abundant ions were selected for fragmentation, and dynamic exclusion was applied to remove unnecessary MS/MS information [42].
4.2.4. Measurement of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
Using the two-fold dilution method, 100 μL of culture medium was pre-added to wells 2 to 8 in a 96-well plate. A total of 200 μL of extract solution was added to well 1, and 100 μL of the sample solution was withdrawn and added to well 2; this pattern was followed for serial two-fold dilutions until well 8. To wells 1 to 8, 10 μL of 1 × 105 CFU/mL bacterial suspension was added sequentially. Well 9 contained an equal volume of culture medium, bacterial suspension, and normal saline, with the latter serving as the control group. Well 10 contained only culture medium, serving as the blank control. After completing the sample addition, the plates were incubated at 37 °C for 24 h. The MIC was determined by visually inspecting the wells for turbidity. For each group, this process was repeated three times. Based on the MIC test results, 10 μL of solution from each well was plated, and the plates were incubated at 37 °C for 24 h to observe colony growth on the agar plates. The MBC of the fermentation extract was defined as the lowest drug concentration at which no colonies grew [43].
4.2.5. Determination of Growth Curve of S. aureus by Fermentation Extract of UJ3-2
We added 100 μL of bacterial suspension (1 − 2 × 107 CFU/mL) to a 96-well cell culture plate, and then added 100 μL of extract at concentrations of 2 MIC, 1.0 MIC, and 0.5 MIC to the respective wells, resulting in final mass concentrations of 0.25 MIC, 0.5 MIC, and 1.0 MIC of the fermentation extract in the wells. We used culture medium instead of extract as a control, with 4 wells for each concentration. We incubated the 96-well cell culture plate at 35 °C for 16 h, measuring OD600nm every 2 h. We plotted a growth curve with time as the horizontal axis and optical density values as the vertical axis [44].
4.2.6. Determination of the Biofilm of S. aureus by Fermentation Extract of UJ3-2
To measure the inhibitory effect of UJ3-2 fermentation extract on S. aureus biofilm, 2.97 mL of TSB (Tryptic Soy Broth) medium was added to surface-treated 12-well plates and inoculated with 30 μL of S. aureus suspension at 109 CFU/mL. Fermentation products were then added to achieve final concentrations of 0.5, 1, and 5 mg/mL, respectively. The wells without fermentation products were considered the control group. The 12-well plates were incubated at 37 °C for 24, 48, and 72 h. After incubation, excess bacterial suspension was aspirated, and the wells were washed with PBS to remove free bacteria. Subsequently, 50 μL of methanol was added to each well to fix the biofilm for 30 min. Following fixation, the methanol was discarded, and the wells were rinsed with PBS. Then, 500 μL of 0.5% (w/v) crystal violet solution was added to stain the biofilm for 15 min. After staining, excess crystal violet solution was discarded, and the wells were washed with PBS. The plates were dried, and 300 μL of 70% ethanol was added. Once the crystal violet in the wells was fully dissolved, the absorbance at OD600nm was measured [45].
4.2.7. Detection of Extracellular Nucleic Acid and Protein of S. aureus by Fermentation Extract of UJ3-2
The bacterial solution cultured to the logarithmic growth phase was washed with sterile PBS, resuspended, and divided into four tubes. UJ3-2 fermentation extract was added to the resuspended solution at final concentrations of 1, 1/2, 1/4, and 1/8 MIC, respectively, and sterile PBS was used to set up a negative control. The samples were incubated in a 37 °C water bath for 12 h. At 0, 2, 4, 6, 8, 10, and 12 h, 200 μL of the samples was taken and centrifuged at 4000 rpm for 5 min. The supernatants were collected to measure the OD260 and OD280 values [46].
4.2.8. Effects of UJ3-2 Fermentation Extract on Membrane Integrity of S. aureus
Using the propidium iodide (PI) staining method, 500 μL of bacterial suspension was added to 2 mL centrifuge tubes. Then, 500 μL of the extract solutions at concentrations of 1 MIC, 2 MIC, and 4 MIC was added to the centrifuge tubes, resulting in final extract concentrations of 0.5 MIC, 1 MIC, and 2 MIC, respectively. Sterile deionized water was used instead of the extract as a control, with 3 tubes for each concentration. The samples were incubated at 28 °C for 4 h. The cultured bacterial suspensions were centrifuged at 5000 rpm for 10 min at 4 °C, and the supernatants were discarded. The obtained bacterial cells were resuspended and washed twice with PBS, followed by centrifugation to discard the supernatant again. The bacterial cells were resuspended in PBS, and PI dye (final concentration of 10 μg/mL) was added. The samples were then incubated in the dark at 4 °C for 30 min. Afterward, the bacterial suspensions were resuspended, washed twice with PBS, centrifuged to discard the supernatant, and resuspended in PBS. Finally, 10 μL of bacterial suspension was taken and observed under a fluorescence microscope with an excitation wavelength of 535 nm and an emission wavelength of 615 nm [47].
4.2.9. Cell Membrane Integrity of S. aureus
The bacterial suspension cultured to the logarithmic growth phase was washed and resuspended using sterile PBS. Fungal fermentation products were added to achieve a final concentration of 1/2 MIC, while sterile PBS was used to set up a negative control. The samples were incubated in a 37 °C water bath for 4 h, and then centrifuged at 4000 rpm for 5 min. The supernatants were discarded, and the bacterial cells were washed twice with sterile PBS. The bacterial cells were resuspended in electron microscopy fixative, fixed at room temperature for 2 h, and then stored at 4 °C. After being left overnight, the bacteria were washed with PBS 3 times and fixed in 1% osmic acid for 30 min, and then gradient elution with different concentrations of ethanol. Finally, the samples were added to a special slide and attached to a field emission scanning electron microscope (SEM) (Hitachi, Tokyo, Japan) stage. After drying for 8 h with a critical point dryer, the samples were gold-plated. The morphology of bacterial cells was observed with the SEM [48].
4.3. Statistical Analyses
Microsoft Office 2016 software was used for creating figures, and SPSS Statistics 20.0 (IBM Corp., Armonk, NY, USA) was used for conducting an ANOVA analysis and t-test on the data. The experiment was repeated three times, and the average of the measured results was taken. The data are presented as means ± SD.
5. Conclusions
The present study underscores the remarkable antibacterial prowess of the metabolites sourced from Xylaria grammica UJ3-2 against S. aureus, a prevalent cause of infections worldwide. By effectively targeting and inhibiting the formation of robust S. aureus biofilms, as well as disrupting the intricate architecture and stability of the bacterial cell wall, these metabolites elicit the release of crucial intracellular constituents, such as nucleic acids and proteins, to varying degrees. This leakage not only disrupts the normal metabolic activities but also significantly impedes the growth and propagation of the bacteria. Thus, this study delves into the initial stages of elucidating the mechanistic pathways underpinning the antibacterial effects of Xylaria grammica UJ3-2 on S. aureus, offering valuable insights and serving as a cornerstone for future endeavors aimed at harnessing natural compounds as innovative antimicrobial agents to counter the rising tide of antibiotic resistance among bacterial pathogens.
Author Contributions
F.L. wrote and revised the manuscript and constructed the tables and figures. J.H., R.L. and Y.H. conceived the study and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Ecological Poultry Science and Technology Innovation team of higher education institutions in Guizhou Province (Grant No. 2023098), the Qingzhen City Science and Technology Project Support Program (Grant No. 2023-25), and Research on Biodiversity Conservation and Development in Karst Regions (2024RW332).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hanselman, B.A.; Kruth, S.A.; Rousseau, J.; Weese, J.S. Coagulase positive staphylococcal colonization of humans and their household pets. Can. Vet. J. 2009, 50, 954–958. [Google Scholar] [PubMed]
- Balcucho, J.; Narváez, D.M.; Castro-Mayorga, J.L. Antimicrobial and Biocompatible Polycaprolactone and Copper Oxide Nanoparticle Wound Dressings against Methicillin-Resistant Staphylococcus aureus. Nanomaterials 2020, 10, 1692. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Mansour, N.; Loubet, P.; Pouget, C.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.P.; Molle, V. Staphylococcus aureus Toxins: An Update on Their Pathogenic Properties and Potential Treatments. Toxins 2021, 13, 677. [Google Scholar] [CrossRef]
- Huang, S.S.; Hinrichsen, V.L.; Datta, R.; Spurchise, L.; Miroshnik, I.; Nelson, K.; Platt, R. Methicillin-resistant Staphylococcus aureus infection and hospitalization in high-risk patients in the year following detection. PLoS ONE 2011, 6, e24340. [Google Scholar] [CrossRef]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.S.; Jain, R.; Bhardwaj, P.; Thakur, A.; Kumari, M.; Bhushan, S.; Kumar, S. Plant probiotics—Endophytes pivotal to plant health. Microbiol. Res. 2022, 263, 127148. [Google Scholar] [CrossRef]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.X.; Ng, S.B. Fungal Endophytes: A Promising Frontier for Discovery of Novel Bioactive Compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef]
- Tiwari, P.; Srivastava, Y.; Bae, H. Trends of Pharmaceutical Design of Endophytes as Anti-infective. Curr. Top. Med. Chem. 2021, 21, 1572–1586. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Matias, R.R.; Sepúlveda, A.M.G.; Batista, B.N.; de Lucena, J.; Albuquerque, P.M. Degradation of Staphylococcus aureus Biofilm Using Hydrolytic Enzymes Produced by Amazonian Endophytic Fungi. Appl. Biochem. Biotechnol. 2021, 193, 2145–2161. [Google Scholar] [CrossRef]
- Sadrati, N.; Zerroug, A.; Demirel, R.; Harzallah, D. Anti-multidrug-resistant Staphylococcus aureus and anti-dermatophyte activities of secondary metabolites of the endophytic fungus Penicillium brevicompactum ANT13 associated with the Algerian endemic plant Abies numidica. Arch. Microbiol. 2023, 205, 110. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Huang, R.; Zhang, S.; Xu, T.; Hu, K.; Wu, S. Antimicrobial secondary metabolites from an endophytic fungus Aspergillus polyporicola. Fitoterapia 2022, 162, 105297. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Ma, S.; Shi, X.; Xue, W.; Liu, C.; Ding, H. Isolation, diversity, and antimicrobial activity of fungal endophytes from Rohdea chinensis (Baker) N.Tanaka (synonym Tupistra chinensis Baker) of Qinling Mountains, China. PeerJ 2020, 17, e9342. [Google Scholar] [CrossRef] [PubMed]
- Perrine-Walker, F.; Le, K. Propidium iodide enabled live imaging of Pasteuria sp.-Pratylenchus zeae infection studies under fluorescence microscopy. Protoplasma 2021, 258, 279–287. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, L.; Zhu, G.; Zuo, M.; Gong, Q.; He, W.; Li, M.; Yuan, C.; Hao, X.; Zhu, W. New phenylpyridone derivatives from the Penicillium sumatrense GZWMJZ-313, a fungal endophyte of Garcinia multiflora. Chin. Chem. Lett. 2019, 30, 431–434. [Google Scholar] [CrossRef]
- Zhong, F.; Fan, X.; Ji, W.; Hai, Z.; Hu, N.; Li, X.; Liu, G.; Yu, C.; Chen, Y.; Lian, B.; et al. Soil Fungal Community Composition and Diversity of Culturable Endophytic Fungi from Plant Roots in the Reclaimed Area of the Eastern Coast of China. J. Fungi 2022, 8, 124. [Google Scholar] [CrossRef]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef]
- Li, K.; Zhang, H.; Shi, M.; Zhang, Y.; Cong, C.; Chang, X.; Duan, L.; Ding, Y. The complete chloroplast genome sequence of Urtica fissa. Mitochondrial DNA B Resour. 2022, 7, 1005–1007. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Guo, Z.; Li, X.; Wang, M. Chemical constituents from Urtica fissa stem and their inhibitory effects on α-glucosidase activity. Nat. Prod. Res. 2021, 35, 3011–3017. [Google Scholar] [CrossRef]
- Feng, X.; Wang, M.; Cheng, J.; Li, X. Two new secolignans with in vitro anti-inflammatory activities from Urtica fissa rhizomes. J. Nat. Med. 2017, 71, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Bao, T.; Tao, G.; Hu, Y.; Han, C. In vitro evaluation of the composition and acaricidal efficacy of Urtica fissa leaf ethyl acetate extract against Sarcoptes scabiei mites. Vet. Med. 2023, 68, 200–207. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Wang, M.Y.; Chen, S.; Li, X.B. The chemical constituents with cytotoxicity from Urtica fissa seed. J. Asian Nat. Prod. Res. 2019, 21, 1068–1074. [Google Scholar] [CrossRef]
- Cong, G.A.O.; Jun, L.U.O.; Xia, L.I.U.; Lin, M.A.; Xiao-Hong, Y. Recent advances in the studies of chemical compositions and bioactivities of the genus Xylaria. Mycosystema 2016, 35, 767–781. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jeon, J.; Kim, J.A.; Jeon, M.J.; Yu, N.H.; Kim, S.; Park, A.R.; Kim, J.C.; Lee, Y.; Kim, Y.; et al. Draft Genome Sequence of Xylaria grammica EL000614, a Strain Producing Grammicin, a Potent Nematicidal Compound. Mycobiology 2021, 49, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Park, A.R.; Duraisamy, K.; Vo, D.D.; Kim, J.C. Elucidation of the nematicidal mode of action of grammicin on Caenorhabditis elegans. Pestic. Biochem. Physiol. 2022, 188, 105244. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium iodide staining underestimates viability of adherent bacterial cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef]
- Chen, L.; Tao, D.; Yu, F.; Wang, T.; Qi, M.; Xu, S. Cineole regulates Wnt/β-catenin pathway through Nrf2/keap1/ROS to inhibit bisphenol A-induced apoptosis, autophagy inhibition and immunosuppression of grass carp hepatocytes. Fish Shellfish Immunol. 2022, 131, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tao, D.; Qi, M.; Wang, T.; Jiang, Z.; Xu, S. Cineole alleviates the BPA-inhibited NETs formation by regulating the p38 pathway-mediated programmed cell death. Ecotoxicol. Environ. Saf. 2022, 237, 113558. [Google Scholar] [CrossRef]
- Lindner, S.; Geu-Flores, F.; Bräse, S.; Sherden, N.H.; O’Connor, S.E. Conversion of substrate analogs suggests a Michael cyclization in iridoid biosynthesis. Chem. Biol. 2014, 21, 1452–1456. [Google Scholar] [CrossRef]
- Grover, P.; Mehta, L.; Malhotra, A.; Kapoor, G.; Nagarajan, K.; Kumar, P.; Chawla, V.; Chawla, P.A. Exploring the Multitarget Potential of Iridoids: Advances and Applications. Curr. Top. Med. Chem. 2023, 23, 371–388. [Google Scholar]
- Wang, C.; Gong, X.; Bo, A.; Zhang, L.; Zhang, M.; Zang, E.; Zhang, C.; Li, M. Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics. Molecules 2020, 25, 287. [Google Scholar] [CrossRef]
- da Silva, A.R.P.; Costa, M.D.S.; Araújo, N.J.S.; de Freitas, T.S.; de Almeida, R.S.; Barbosa Filho, J.M.; Tavares, J.F.; de Souza, E.O.; de Farias, P.A.M.; Pinheiro, J.C.A.; et al. Potentiation of Antibiotic Action and Efflux Pump Inhibitory Effect on Staphylococcus aureus Strains by Solasodine. Antibiotics 2022, 11, 1309. [Google Scholar] [CrossRef]
- Navabharath, M.; Srivastava, V.; Gupta, S.; Singh, S.V.; Ahmad, S. Ursolic Acid and Solasodine as Potent Anti-Mycobacterial Agents for Combating Paratuberculosis: An Anti-Inflammatory and in Silico Analysis. Molecules 2022, 28, 274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xie, Y.; Qiu, W.; Mei, J.; Xie, J. Antibacterial and Antibiofilm Efficacy and Mechanism of Ginger (Zingiber officinale) Essential Oil against Shewanella putrefaciens. Plants 2023, 12, 1720. [Google Scholar] [CrossRef]
- Shin, B.; Park, C.; Imlay, J.A.; Park, W. 4-Hydroxybenzaldehyde sensitizes Acinetobacter baumannii to amphenicols. Appl. Microbiol. Biotechnol. 2018, 102, 2323–2335. [Google Scholar] [CrossRef]
- Uçar, K.; Göktaş, Z. Biological activities of naringenin: A narrative review based on in vitro and in vivo studies. Nutr. Res. 2023, 119, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Górecki, M.; Rzepecka-Stojko, A.; Balwierz, R.; Stojko, J. Bee Products in Dermatology and Skin Care. Molecules 2020, 25, 556. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Zhang, W.; Wu, C.; Feng, H.; Peng, Y.; Shahid, H.; Cui, Z.; Ding, P.; Shan, T. Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbiol. 2021, 21, 155. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens 2021, 10, 165. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, R.; Li, L.; Zhou, X.; Li, Z.; Jia, R.; Song, X.; Zou, Y.; Yin, L.; He, C.; et al. The Antibacterial Mechanism of Terpinen-4-ol Against Streptococcus agalactiae. Curr. Microbiol. 2018, 75, 1214–1220. [Google Scholar] [CrossRef]
- Yang, X.; Lan, W.; Xie, J. Antimicrobial and anti-biofilm activities of chlorogenic acid grafted chitosan against Staphylococcus aureus. Microb. Pathog. 2022, 173, 105748. [Google Scholar] [CrossRef]
- Yang, H.; Li, G.; Huai, R.; Yang, D.; Lei, L.; Ouyang, Y.; Xiong, L. Isolation, identification and antibacterial activity of endophytes of Coptis chinensis Franch. Nat. Prod. Res. Dev. 2022, 34, 1456–1471. [Google Scholar]
- Zhou, Y.; Yao, Q.; Zhang, T.; Chen, X.; Wu, Z.; Zhang, N.; Shao, Y.; Cheng, Y. Antibacterial activity and mechanism of green tea polysaccharide conjugates against Escherichia coli. Ind. Crops Prod. 2020, 152, 112464. [Google Scholar] [CrossRef]
- Gowrishankar, S.; Kamaladevi, A.; Ayyanar, K.S.; Balamurugan, K.; Pandian, S.K. Bacillus amyloliquefaciens-secreted cyclic dipeptide—cyclo(l-leucyl-l-prolyl) inhibits biofilm and virulence production in methicillin-resistant Staphylococcus aureus. RSC Adv. 2015, 5, 95788–95804. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).