
Research Progress on the Mechanism for Improving Glucose and Lipid Metabolism Disorders Using Phenolic Acid Components from Medicinal and Edible Homologous Plants

 , ,
, ,
Abstract

1. Introduction
2. Mechanism of Action of MEHHs Phenolic Acids in Regulating Glucose and Lipid Metabolism Disorders
2.1. Regulates Glucose Metabolism
2.1.1. Inhibition of α-Amylase and α-Glucosidase Activity to Reduce Carbohydrate Cleavage
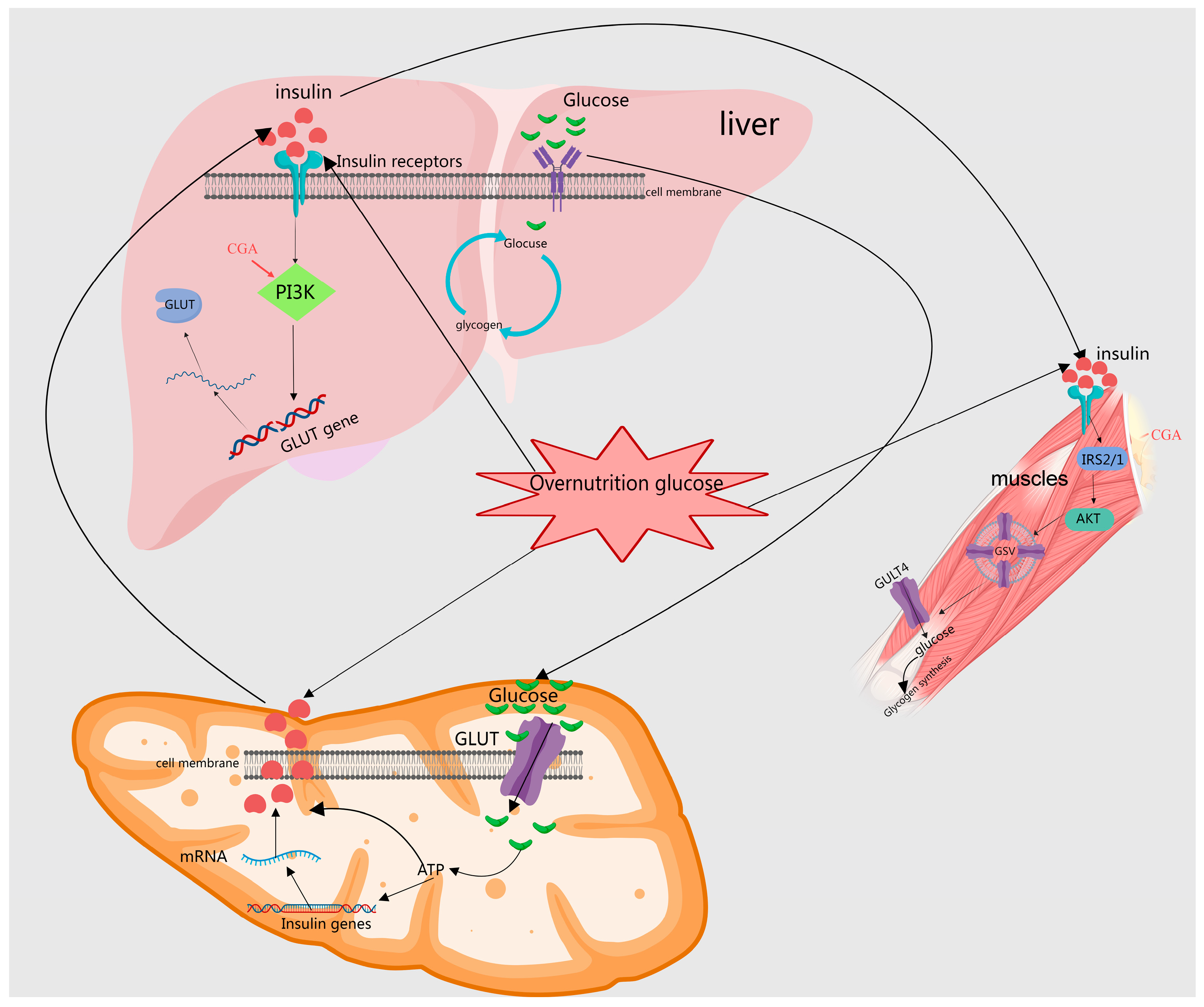
2.1.2. Enhancing the Expression or Translocation of Glucose Transporters (GLUT) to Augment Glucose Uptake
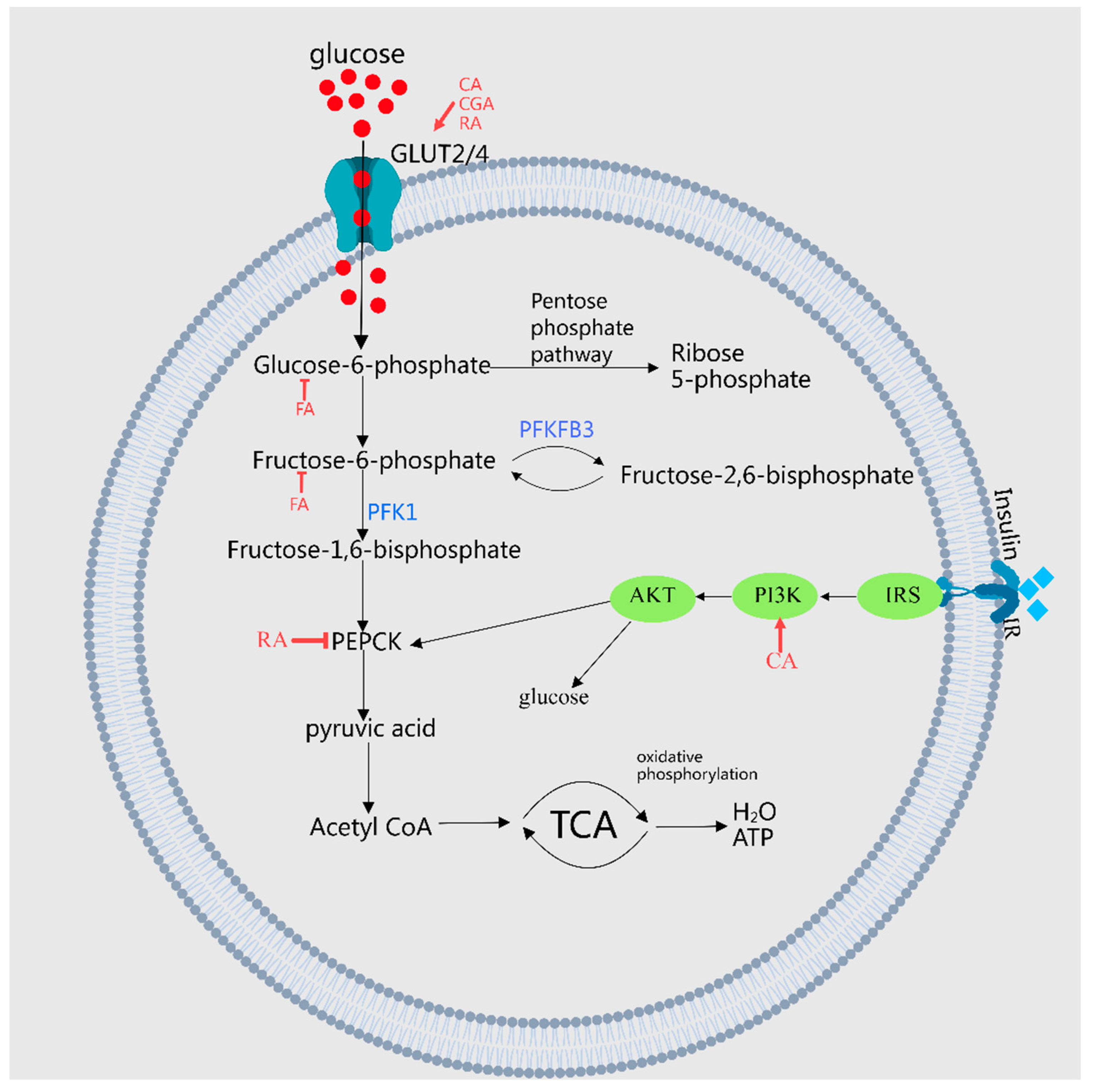
2.1.3. Inhibition of Gluconeogenesis, Promotion of Glycogen Synthesis, and Regulation of Blood Glucose Levels
2.2. Regulates Lipid Metabolism
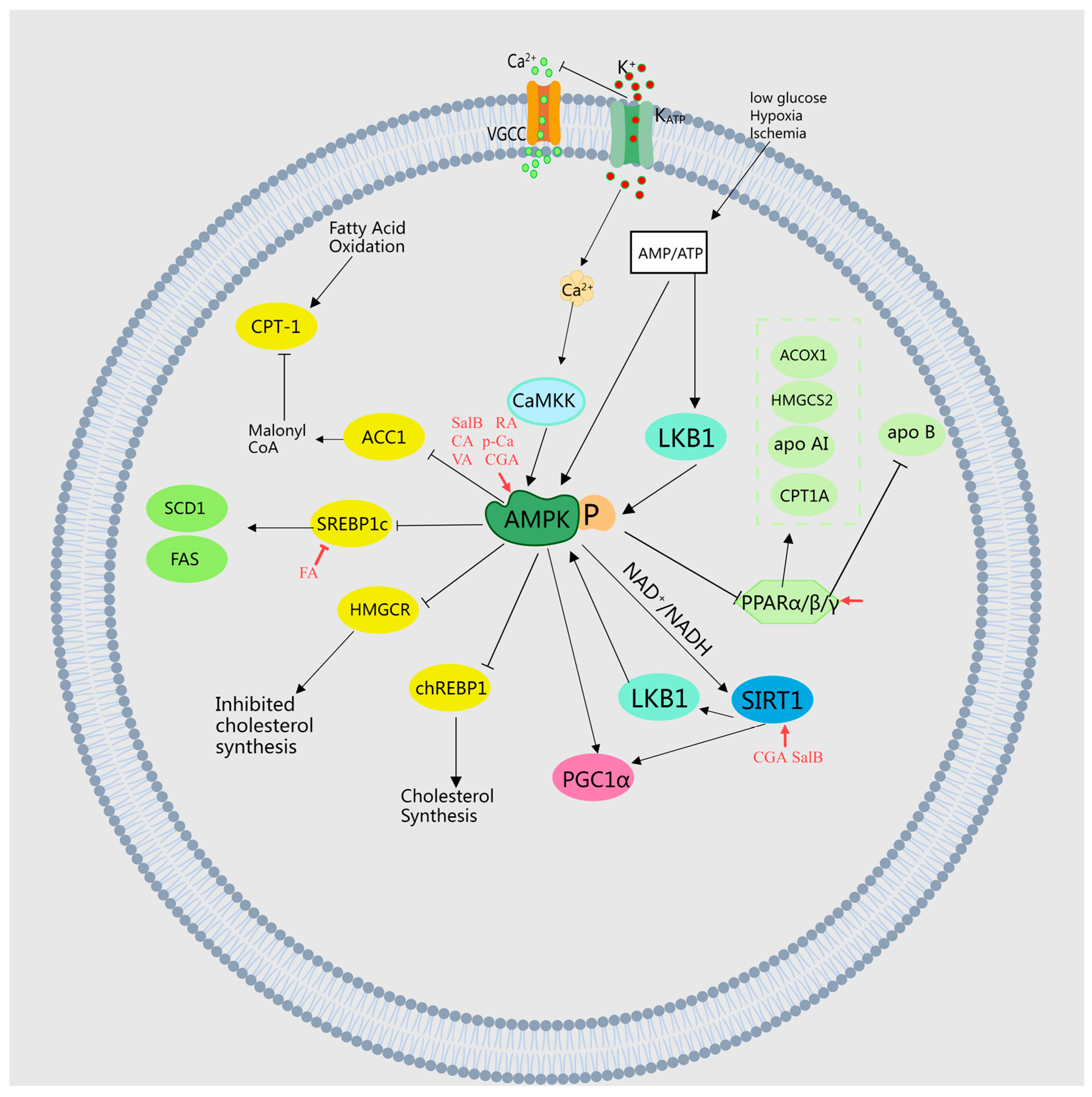
2.2.1. AMPK, SREBP1, and ACC Signaling Pathways
2.2.2. Enhancing the Oxidative Activity of Beta Fatty Acids and Expression of PPARs
2.3. Regulates Insulin Signaling, Improves Insulin Sensitivity and Improves Insulin Resistance
2.4. Inhibits Inflammatory Responses
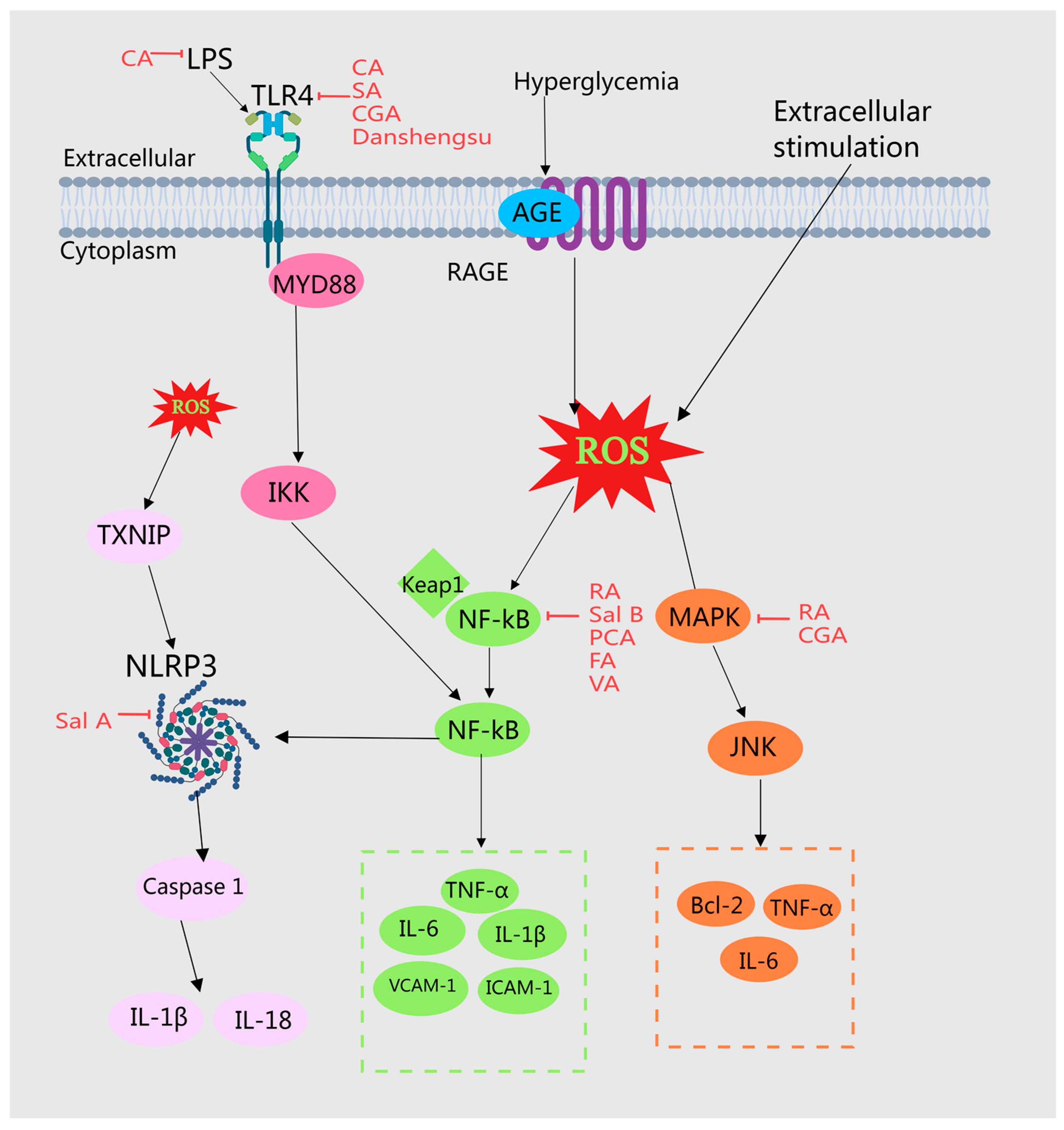
2.4.1. Accessing TLRs, NF-κB, and NLRP3 Pathways
2.4.2. MAPK Signaling Pathway
2.5. Inhibition of Oxidative Stress
2.5.1. The Nrf2 Signaling Pathway
2.5.2. Endoplasmic Reticulum Stress
2.5.3. Regulation of Oxidation-Related Factors
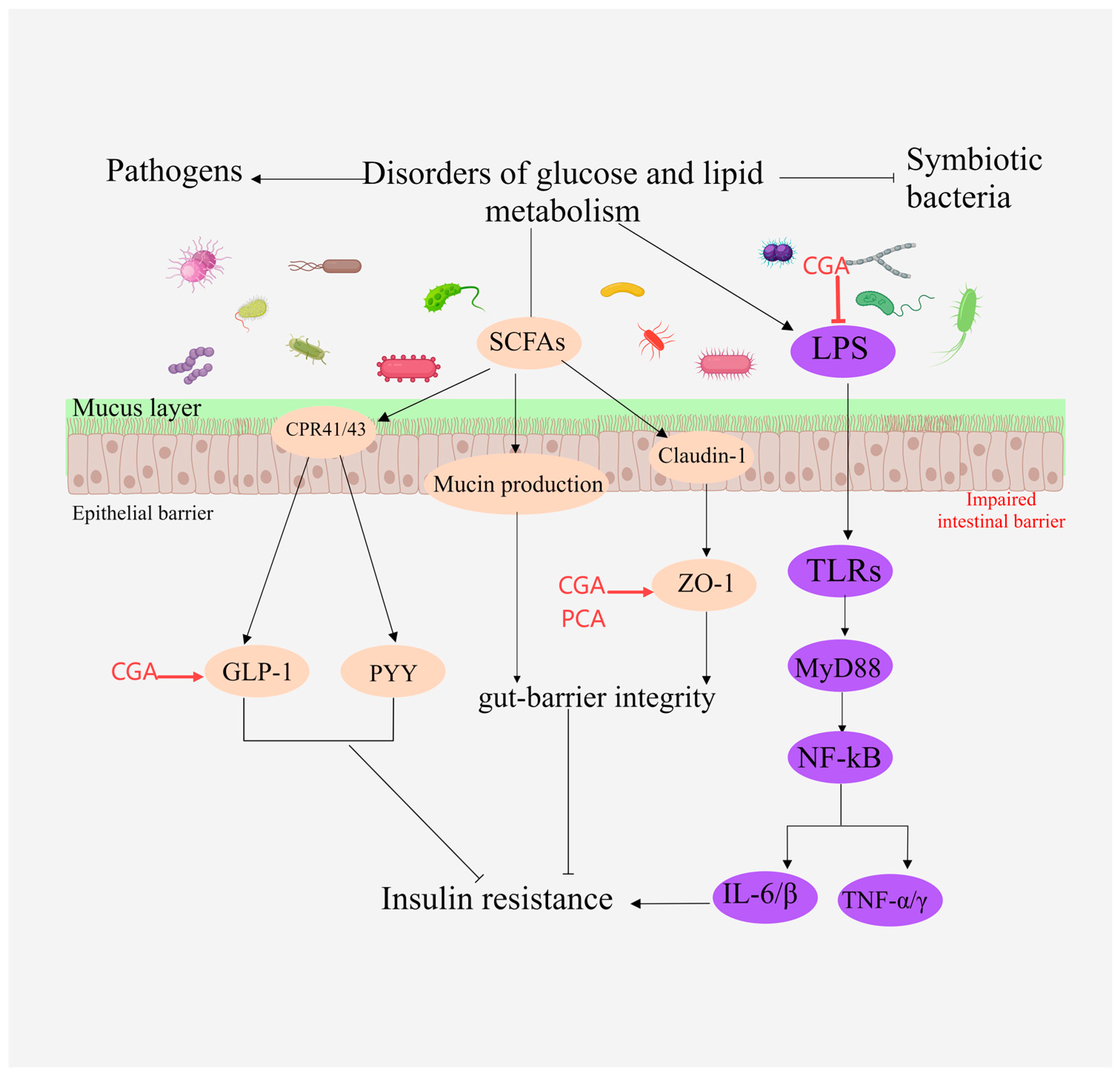
2.6. Regulates Intestinal Flora
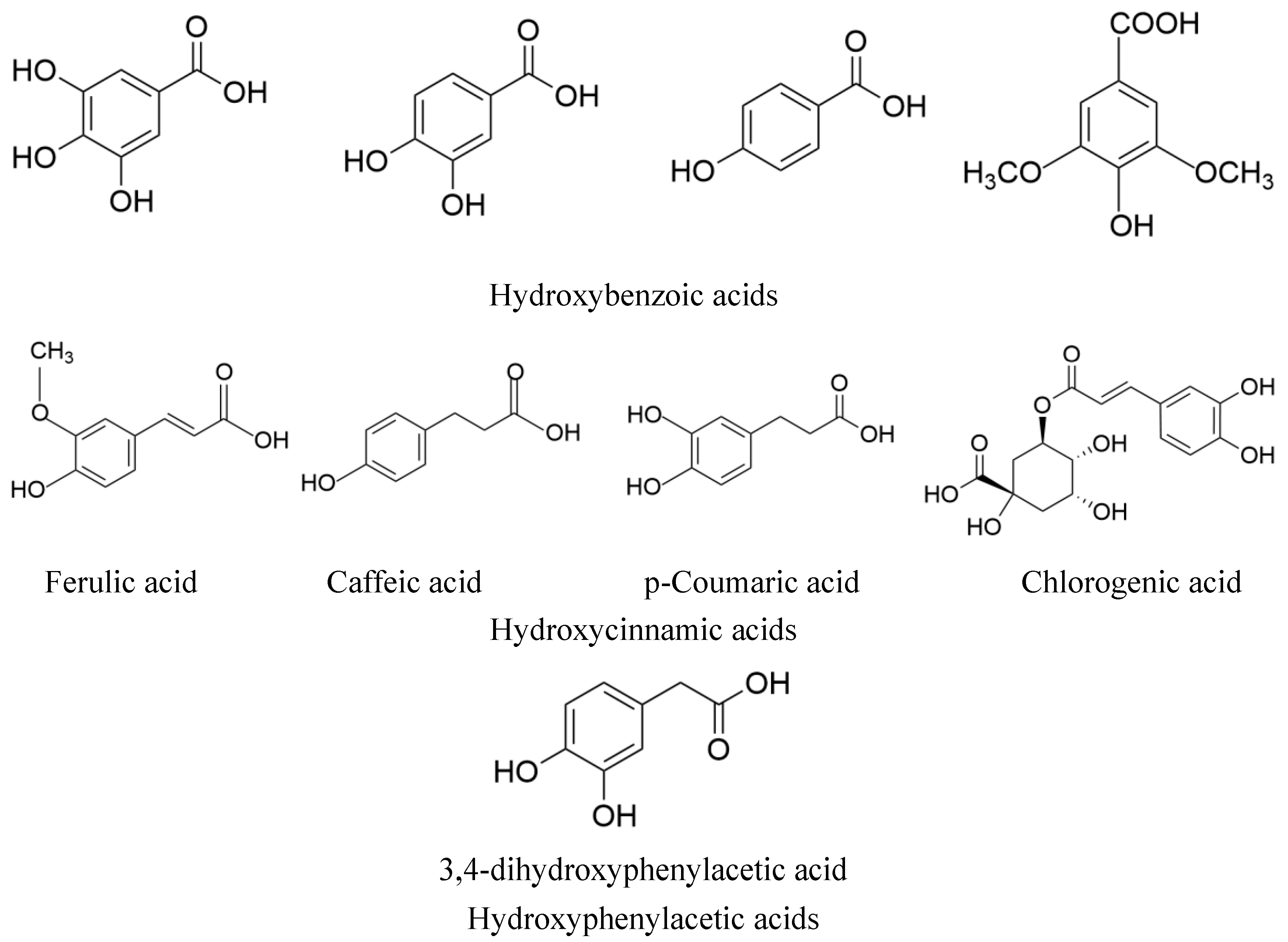
3. The Relationship between the Structure of MEHHs Phenolic Acids and the Regulation of Glucose and Lipid Metabolism Disorders
3.1. Hydroxybenzoic Acids
3.1.1. Number of Hydroxy Groups
3.1.2. Position of Hydroxyl Group
3.2. Hydroxycinnamic Acids
3.2.1. Number of Hydroxy Groups
3.2.2. Position of Hydroxyl Group
4. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AS atherosclerosis | MAPK mitogen-activated protein kinase |
| Apo B apolipoprotein B | MDA malonaldehyde |
| AGEs advanced glycosylation end products | Nrf2 nuclear factor erythroid2-related factor 2 |
| AMPK AMP-activated protein kinase | NAFLD non-alcoholic fatty liver disease |
| AchE acetylcholinesterase | NF-κB nuclear factor kappa-B |
| ACC acetyl CoA carboxylase | NLRP3 NOD-like receptor thermal protein domain associated protein 3 |
| CAT catalase | OS oxidative stress |
| CA caffeic acid | PCA protocatechuic acid |
| CGA chlorogenic acid | P-Ca p-coumaric acid |
| CPT-1 carnitine palmitoyl transterase-1 | PPARs peroxisome proliferators-activated receptors |
| DM diabetes mellitus | RA rosmarinus acid |
| FA ferulic acid | ROS reactive oxygen species |
| FFA non-esterified fatty acid | RNS reactive nitrogen species |
| GA gallic acid | SA salicylic acid |
| GSH-Px glutathione peroxidase | SOD super oxide dismutase |
| GSSG oxidized glutathione | STZ streptozotocin |
| GSH L-glutathione | SalA salvianolic acid A |
| GST glutathione S-transferase | SalB salvianolic acid B |
| HO-1 heme oxygenase-1 | T1D type1 diabetes mellitus |
| HUVECs human umbilical vein endothelial cells | T2D type 2 diabetes mellitus |
| IL-1β interleukin-1β | TNF-α tumor necrosis factor alpha |
| IL eukin-6 | TGF-β1 transforming growth factor-β |
| IR ischemia–reperfusion injury | TG triglyceride |
| ICAM-1 intercellular cell adhesion molecule-1 | TC serum total cholesterol |
| LDL-C low density lipoprotein-cholesterol | VCAM-1 vascular cell adhesion molecule-1 |
References
- Li, X.; Zhang, Y.; Wang, S.; Shi, C.; Wang, S.; Wang, X.; Lü, X. A review on the potential use of natural products in overweight and obesity. Phytother. Res. 2022, 36, 1990–2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, C.; Zhou, J.; Zhou, F.; Gui, A.; Chu, H.; Shao, Q. Chrysanthemum morifolium as a traditional herb: A review of historical development, classification, phytochemistry, pharmacology and application. J. Ethnopharmacol. 2024, 330, 118198. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Xiong, S.; Li, Y.; Xia, B.; Li, M.; Zhang, Z. Phenolic acids from medicinal and edible homologous plants: A potential anti-inflammatory agent for inflammatory diseases. Front. Immunol. 2024, 15, 1345002. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Ijaz, M.; Buabeid, M.; Kharaba, Z.J.; Yaseen, H.S.; Murtaza, G. Therapeutic Effects and Safe Uses of Plant-Derived Polyphenolic Compounds in Cardiovascular Diseases: A Review. Drug Des. Dev. Ther. 2021, 15, 4713–4732. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wei, X.; Yan, X.; Jin, T.; Ling, W. Protocatechuic acid, a metabolite of anthocyanins, inhibits monocyte adhesion and reduces atherosclerosis in apolipoprotein E-deficient mice. J. Agric. Food Chem. 2010, 58, 12722–12728. [Google Scholar] [CrossRef]
- Ikegami, H.; Hiromine, Y.; Noso, S. Insulin-dependent diabetes mellitus in older adults: Current status and future prospects. Geriatr. Gerontol. Int. 2022, 22, 549–553. [Google Scholar] [CrossRef]
- Schrack, J.A.; Dougherty, R.J.; Corkum, A.; Liu, F.; Wanigatunga, A.A. Impact of Time in Motion on Blood Pressure Regulation Among Patients with Metabolic Syndrome. Curr. Hypertens. Rep. 2022, 24, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, T.; Hasebe, N. Impact of risk factors related to metabolic syndrome on acute myocardial infarction in younger patients. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2022, 45, 1447–1458. [Google Scholar] [CrossRef]
- Rahman, M.M.; Dhar, P.S.; Sumaia; Anika, F.; Ahmed, L.; Islam, M.R. Exploring the plant-derived bioactive substances as antidiabetic agent: An extensive review. Biomed. Pharmacother. 2022, 152, 113217. [Google Scholar] [CrossRef]
- Tian, J.; Qin, S.; Han, J.; Meng, J.; Liang, A. A review of the ethnopharmacology, phytochemistry, pharmacology and toxicology of Fructus Gardeniae (Zhi-zi). J. Ethnopharmacol. 2022, 289, 114984. [Google Scholar] [CrossRef]
- Gao, S.; Feng, Q. The Beneficial Effects of Geniposide on Glucose and Lipid Metabolism: A Review. Drug Des. Dev. Ther. 2022, 16, 3365–3383. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, I.; Benariba, N.; Adjdir, S.; Fekhikher, Z.; Daoud, I.; Terki, M.; Benramdane, H.; Ghalem, S. In silico evaluation of phenolic compounds as inhibitors of A-amylase and A-glucosidase. J. Biomol. Struct. Dyn. 2021, 39, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Salau, V.F.; Erukainure, O.L.; Ijomone, O.M.; Islam, M.S. Caffeic acid regulates glucose homeostasis and inhibits purinergic and cholinergic activities while abating oxidative stress and dyslipidaemia in fructose-streptozotocin-induced diabetic rats. J. Pharm. Pharmacol. 2022, 74, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Pan, Y.; Wang, L.; Zhou, H.; Li, A. Research advances on the distribution and physiological activity of ferulic acid in grain. Sci. Technol. Cereals Oils Foods 2019, 27, 43–48. [Google Scholar]
- Salau, V.F.; Erukainure, O.L.; Olofinsan, K.O.; Bharuth, V.; Ijomone, O.M.; Islam, M.S. Ferulic acid improves glucose homeostasis by modulation of key diabetogenic activities and restoration of pancreatic architecture in diabetic rats. Fundam. Clin. Pharmacol. 2023, 37, 324–339. [Google Scholar] [CrossRef]
- Liao, C.C.; Ou, T.T.; Wu, C.H.; Wang, C.J. Prevention of diet-induced hyperlipidemia and obesity by caffeic acid in C57BL/6 mice through regulation of hepatic lipogenesis gene expression. J. Agric. Food Chem. 2013, 61, 11082–11088. [Google Scholar] [CrossRef]
- Roy, J.R.; Janaki, C.S.; Jayaraman, S.; Veeraraghavan, V.P.; Periyasamy, V. Hypoglycemic Potential of Carica papaya in Liver Is Mediated through IRS-2/PI3K/SREBP-1c/GLUT2 Signaling in High-Fat-Diet-Induced Type-2 Diabetic Male Rats. Toxics 2023, 11, 240. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Cao, H. Chlorogenic acid and caffeic acid from Sonchus oleraceus Linn synergistically attenuate insulin resistance and modulate glucose uptake in HepG2 cells. Food Chem. Toxicol. 2019, 127, 182–187. [Google Scholar] [CrossRef]
- Runtuwene, J.; Cheng, K.C.; Asakawa, A.; Amitani, H.; Amitani, M.; Morinaga, A.; Takimoto, Y.; Kairupan, B.H.; Inui, A. Rosmarinic acid ameliorates hyperglycemia and insulin sensitivity in diabetic rats, potentially by modulating the expression of PEPCK and GLUT4. Drug Des. Dev. Ther. 2016, 10, 2193–2202. [Google Scholar]
- Jung, E.H.; Kim, S.R.; Hwang, I.K.; Ha, T.Y. Hypoglycemic effects of a phenolic acid fraction of rice bran and ferulic acid in C57BL/KsJ-db/db mice. J. Agric. Food Chem. 2007, 55, 9800–9804. [Google Scholar] [CrossRef]
- Naowaboot, J.; Piyabhan, P.; Munkong, N.; Parklak, W.; Pannangpetch, P. Ferulic acid improves lipid and glucose homeostasis in high-fat diet-induced obese mice. Clin. Exp. Pharmacol. Physiol. 2016, 43, 242–250. [Google Scholar] [CrossRef]
- Huang, D.W.; Chang, W.C.; Wu, J.S.; Shih, R.W.; Shen, S.C. Gallic acid ameliorates hyperglycemia and improves hepatic carbohydrate metabolism in rats fed a high-fructose diet. Nutr. Res. 2016, 36, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Jiang, X.; Wang, Y.; Fan, Y. Effects of Salvianolic Acid B on Atherosclerotic Plaque Area and Expressions of AGEs/RAGE in ApoE-Gene Knock-out Mice Treated with STZ and High Fat Diet. Tianjin Med. J. 2010, 38, 777–780. [Google Scholar]
- Han, Y.; Hu, Z.; Cui, A.; Liu, Z.; Ma, F.; Xue, Y.; Liu, Y. Post-translational regulation of lipogenesis via AMPK-dependent phosphorylation of insulin-induced gene. Nat. Commun. 2019, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, Y.; Li, M.; Huang, Z.; Jiang, J.; Chen, Y. Ferulic Acid Ameliorates Atherosclerotic Injury by Modulating Gut Microbiota and Lipid Metabolism. Front. Pharmacol. 2021, 12, 621339. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, J.; Xu, F.; Chu, C.; Li, X.; Shi, X.; Zheng, W. Use of Ferulic Acid in the Management of Diabetes Mellitus and Its Complications. Molecules 2022, 27, 6010. [Google Scholar] [CrossRef]
- Lu, W.; Wen, Z.; Liu, J.; Liao, P. Improvement and mechanism of salvianolic acid B on non-alcoholic fatty liver in ApoE knockout mice. Chin. Pharmacol. Bull. 2020, 36, 31–37. [Google Scholar]
- Wang, S.J.; Chen, Q.; Liu, M.Y.; Yu, H.Y.; Xu, J.Q. Regulation effects of rosemary (Rosmarinus officinalis Linn.) on hepatic lipid metabolism in OA induced NAFLD rats. Food Funct. 2019, 10, 7356–7365. [Google Scholar] [CrossRef]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K.I.; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Mu, H.N.; Zhou, Q.; Yang, R.Y.; Tang, W.Q.; Li, H.X.; Wang, S.M. Caffeic acid prevents non-alcoholic fatty liver disease induced by a high-fat diet through gut microbiota modulation in mice. Food Res. Int. 2021, 143, 110240. [Google Scholar] [CrossRef]
- Kim, H.M.; Kim, Y.; Lee, E.S.; Huh, J.H.; Chung, C.H. Caffeic acid ameliorates hepatic steatosis and reduces ER stress in high fat diet-induced obese mice by regulating autophagy. Nutrition 2018, 55–56, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Mourya, A.; Ahuja, S.; Sah, S.P.; Kumar, A. Plausible anti-inflammatory mechanism of resveratrol and caffeic acid against chronic stress-induced insulin resistance in mice. Inflammopharmacology 2016, 24, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sharma, A.; Iqbal, M.S.; Srivastava, J.K. Therapeutic Promises of Chlorogenic Acid with Special Emphasis on its Anti-Obesity Property. Curr. Mol. Pharmacol. 2020, 13, 7–16. [Google Scholar] [CrossRef]
- Yoon, S.Y.; Lee, J.H.; Kwon, S.J.; Kang, H.J.; Chung, S.J. Ginkgolic acid as a dual-targeting inhibitor for protein tyrosine phosphatases relevant to insulin resistance. Bioorg. Chem. 2018, 81, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Shekari, S.; Khonsha, F.; Rahmati-Yamchi, M.; Nejabati, H.R.; Mota, A. Vanillic Acid and Non-Alcoholic Fatty Liver Disease: A Focus on AMPK in Adipose and Liver Tissues. Curr. Pharm. Des. 2021, 27, 4686–4692. [Google Scholar] [CrossRef]
- Luo, Z.; Li, M.; Yang, Q.; Zhang, Y.; Liu, F.; Gong, L.; Han, L.; Wang, M. Ferulic Acid Prevents Nonalcoholic Fatty Liver Disease by Promoting Fatty Acid Oxidation and Energy Expenditure in C57BL/6 Mice Fed a High-Fat Diet. Nutrients 2022, 14, 2530. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic acid improves high fat diet-induced hepatic steatosis and insulin resistance in mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef]
- Lin, C.Y.; Tsai, S.J.; Huang, C.S.; Yin, M.C. Antiglycative effects of protocatechuic acid in the kidneys of diabetic mice. J. Agric. Food Chem. 2011, 59, 5117–5124. [Google Scholar] [CrossRef]
- Luo, C.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Zhang, C.; Zhu, W.; Jin, Y. Rosmarinic acid exerts an antagonistic effect on nonalcoholic fatty liver disease by regulating the YAP1/TAZ-PPARγ/PGC-1α signaling pathway. Phytother. Res. 2021, 35, 1010–1022. [Google Scholar] [CrossRef]
- Li, S. The Regulation of Lipid Metabolism in Rats with Hyperlipidemia by Salvianic acid via Enhanced Expression of LCAT and CYP7A1. Mod. Food Sci. Technol. 2018, 34, 31–35+186. [Google Scholar]
- Chao, C.Y.; Mong, M.C.; Chan, K.C.; Yin, M.C. Anti-glycative and anti-inflammatory effects of caffeic acid and ellagic acid in kidney of diabetic mice. Mol. Nutr. Food Res. 2010, 54, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, Y.; He, H. Mechanism research of chlorogenie acids on insulin resistance in nutritional obese rats. Chongqing Med. J. 2015, 44, 2457–2460. [Google Scholar]
- Zhao, M.; Ren, Y.; Tao, S.; Zheng, S. Salvianolic acid B inhibits cardiomyocyte apoptosis in diabetic rats induced by fluctuating high glucose. Chin. Tradit. Pat. Med. 2021, 43, 2177–2180. [Google Scholar]
- Yang, Y.; Pei, K.; Zhang, Q.; Wang, D.; Feng, H.; Du, Z.; Zhang, C.; Gao, Z.; Yang, W.; Wu, J.; et al. Salvianolic acid B ameliorates atherosclerosis via inhibiting YAP/TAZ/JNK signaling pathway in endothelial cells and pericytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158779. [Google Scholar] [CrossRef]
- Gao, Y.; Tian, R.; Liu, H.; Xue, H.; Zhang, R.; Han, S.; Ji, L.; Huang, W.; Zhan, J.; You, Y. Research progress on intervention effect and mechanism of protocatechuic acid on nonalcoholic fatty liver disease. Crit. Rev. Food Sci. Nutr. 2022, 62, 9053–9075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kaur, G.; Kumar, M.; Kushwah, A.S.; Kabra, A.; Kainth, R. Caffeic Acid Prevents Vascular Oxidative Stress and Atherosclerosis against Atherosclerogenic Diet in Rats. Evid.-Based Complement. Altern. Med. 2022, 2022, 8913926. [Google Scholar] [CrossRef]
- Zeng, N.; Xu, M.; Zhuang, L. The effect of Danshensu on atherosclerosis in rats and its mechanisms. Immunol. J. 2020, 36, 884–889+896. [Google Scholar]
- Shi, A.; Li, T.; Zheng, Y.; Song, Y.; Wang, H.; Wang, N.; Dong, L.; Shi, H. Chlorogenic Acid Improves NAFLD by Regulating gut Microbiota and GLP-1. Front. Pharmacol. 2021, 12, 693048. [Google Scholar] [CrossRef]
- Yan, H.; Gao, Y.Q.; Zhang, Y.; Wang, H.; Liu, G.S.; Lei, J.Y. Chlorogenic acid alleviates autophagy and insulin resistance by suppressing JNK pathway in a rat model of nonalcoholic fatty liver disease. J. Biosci. 2018, 43, 287–294. [Google Scholar] [CrossRef]
- Liu, Y.; Song, Z.; Peng, B.; Xu, S. Effects of Chlorogenic Acid on Apoptosis-Related Gene Expressions in Rats with Non-Alcoholic Fatty Liver Disease Caused by High-Fat Diet. Chin. J. Anim. Nutr. 2015, 27, 2140–2149. [Google Scholar]
- Ham, J.R.; Lee, H.I.; Choi, R.Y.; Sim, M.O.; Seo, K.I.; Lee, M.K. Anti-steatotic and anti-inflammatory roles of syringic acid in high-fat diet-induced obese mice. Food Funct. 2016, 7, 689–697. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic Acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef]
- Zheng, J.; Li, Q.; He, L.; Weng, H.; Su, D.; Liu, X.; Ling, W.; Wang, D. Protocatechuic Acid Inhibits Vulnerable Atherosclerotic Lesion Progression in Older Apoe−/− Mice. J. Nutr. 2020, 150, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Ramar, M.; Manikandan, B.; Raman, T.; Priyadarsini, A.; Palanisamy, S.; Vaseeharan, B. Protective effect of ferulic acid and resveratrol against alloxan-induced diabetes in mice. Eur. J. Pharmacol. 2012, 690, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Yang, Q.; Chen, J.; Yu, C.; Zhang, L.; Zhou, W.; Chen, M. Salvianolic Acid A Ameliorates Early-Stage Atherosclerosis Development by Inhibiting NLRP3 Inflammasome Activation in Zucker Diabetic Fatty Rats. Molecules 2020, 25, 1089. [Google Scholar] [CrossRef]
- Ding, C.; Zhao, Y.; Shi, X.; Zhang, N.; Zu, G.; Li, Z.; Zhou, J. New insights into salvianolic acid A action: Regulation of the TXNIP/NLRP3 and TXNIP/ChREBP pathways ameliorates HFD-induced NAFLD in rats. Sci. Rep. 2016, 6, 28734. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Song, X.; Li, L.; Sun, J.; Jaiswal, Y.; Huang, J.; Liu, C.; Yang, W.; Williams, L. Protective effects of p-coumaric acid against oxidant and hyperlipidemia-an in vitro and in vivo evaluation. Biomed. Pharmacother. 2019, 111, 579–587. [Google Scholar] [CrossRef]
- Villalobos-Labra, R.; Subiabre, M.; Toledo, F.; Pardo, F.; Sobrevia, L. Endoplasmic reticulum stress and development of insulin resistance in adipose, skeletal, liver, and foetoplacental tissue in diabesity. Mol. Asp. Med. 2019, 66, 49–61. [Google Scholar] [CrossRef]
- Preetha Rani, M.R.; Salin Raj, P.; Nair, A.; Ranjith, S.; Rajankutty, K.; Raghu, K.G. In vitro and in vivo studies reveal the beneficial effects of chlorogenic acid against ER stress mediated ER-phagy and associated apoptosis in the heart of diabetic rat. Chem. Biol. Interact. 2022, 351, 109755. [Google Scholar] [CrossRef]
- Mohan, S.; Nair, A.; Poornima, M.S.; Raghu, K.G. Vanillic acid mitigates hyperinsulinemia induced ER stress mediated altered calcium homeostasis, MAMs distortion and surplus lipogenesis in HepG2 cells. Chem. Biol. Interact. 2023, 375, 110365. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zheng, S.; Sheng, Y.; Miao, T.; Xu, J.; Xu, W.; Huang, K.; Zhao, C. Chlorogenic acid ameliorates obesity by preventing energy balance shift in high-fat diet induced obese mice. J. Sci. Food Agric. 2021, 101, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Valanciene, E.; Jonuskiene, I.; Syrpas, M.; Augustiniene, E.; Matulis, P.; Simonavicius, A.; Malys, N. Advances and Prospects of Phenolic Acids Production, Biorefinery and Analysis. Biomolecules 2020, 10, 874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jia, X.; Xu, Y.; Xie, Q.; Zhu, M.; Zhang, H. Effects of Coix Seed Extract, Bifidobacterium BPL1, and Their Combination on the Glycolipid Metabolism in Obese Mice. Front. Nutr. 2022, 9, 939423. [Google Scholar] [CrossRef] [PubMed]
- Riyaphan, J.; Pham, D.C.; Leong, M.K.; Weng, C.F. In Silico Approaches to Identify Polyphenol Compounds as α-Glucosidase and α-Amylase Inhibitors against Type-II Diabetes. Biomolecules 2021, 11, 1877. [Google Scholar] [CrossRef]
- Saleem, F.; Khan, K.M.; Chigurupati, S.; Andriani, Y.; Solangi, M.; Hameed, S. Dicyanoanilines as potential and dual inhibitors of α-amylase and α-glucosidase enzymes: Synthesis, characterization, in vitro, in silico, and kinetics studies. Arab. J. Chem. 2022, 15, 103651. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Huang, D.; Chen, S.; Xia, Y.; Zhu, S. The inhibitory mechanism of chlorogenic acid and its acylated derivatives on α-amylase and α-glucosidase. Food Chem. 2022, 372, 131334. [Google Scholar] [CrossRef]
- Oršolić, N.; Sirovina, D.; Odeh, D.; Gajski, G.; Balta, V.; Šver, L. Efficacy of Caffeic Acid on Diabetes and Its Complications in the Mouse. Molecules 2021, 26, 3262. [Google Scholar] [CrossRef]
- Ismail, A.; Tanasova, M. Importance of GLUT Transporters in Disease Diagnosis and Treatment. Int. J. Mol. Sci. 2022, 23, 8698. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chan, M.H.; Yang, Y.F.; Li, C.H.; Hsiao, M. Glucose transporter 4: Insulin response mastermind, glycolysis catalyst and treatment direction for cancer progression. Cancer Lett. 2023, 563, 216179. [Google Scholar] [CrossRef]
- Rutter, G.A.; Georgiadou, E.; Martinez-Sanchez, A.; Pullen, T.J. Metabolic and functional specialisations of the pancreatic beta cell: Gene disallowance, mitochondrial metabolism and intercellular connectivity. Diabetologia 2020, 63, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Na, R.S.; Ma, C.; Liu, Q.R.; Wu, L.M.; Zheng, X.L. Itraconazole attenuates hepatic gluconeogenesis and promotes glucose uptake by regulating AMPK pathway. Exp. Ther. Med. 2018, 15, 2165–2171. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen metabolism in humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.T.; Fuchs, C.J.; Betts, J.A.; van Loon, L.J. Liver glycogen metabolism during and after prolonged endurance-type exercise. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E543–E553. [Google Scholar] [CrossRef]
- Bouskila, M.; Hunter, R.W.; Ibrahim, A.F.; Delattre, L.; Peggie, M.; van Diepen, J.A.; Voshol, P.J.; Jensen, J.; Sakamoto, K. Allosteric regulation of glycogen synthase controls glycogen synthesis in muscle. Cell Metab. 2010, 12, 456–466. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Pang, J.; Zhang, H.; Luo, J.; Qian, X.; Chen, Q.; Ling, W. Attenuation of Atherosclerosis by Protocatechuic Acid via Inhibition of M1 and Promotion of M2 Macrophage Polarization. J. Agric. Food Chem. 2019, 67, 807–818. [Google Scholar] [CrossRef]
- Zeng, W.; Shan, W.; Gao, L.; Gao, D.; Hu, Y.; Wang, G.; Zhang, N.; Li, Z. Inhibition of HMGB1 release via salvianolic acid B-mediated SIRT1 up-regulation protects rats against non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 16013. [Google Scholar] [CrossRef]
- Le, T.K.C.; Dao, X.D.; Nguyen, D.V.; Luu, D.H.; Bui, T.M.H. Insulin signaling and its application. Front. Endocrinol. 2023, 14, 1226655. [Google Scholar] [CrossRef]
- Saltiel, A.R. Insulin signaling in health and disease. J. Clin. Investig. 2021, 131, 1. [Google Scholar] [CrossRef]
- Zheng, S.; Zhu, Y.; Tao, S.; Zheng, H. Effect of salvianolic acid B on intermittent high glucose induced JNK activation and INS-1 cell apoptosis. Chin. Pharmacol. Bull. 2017, 33, 68–73. [Google Scholar]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Church, J.S.; Kigerl, K.A.; Lerch, J.K.; Popovich, P.G.; McTigue, D.M. TLR4 Deficiency Impairs Oligodendrocyte Formation in the Injured Spinal Cord. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 6352–6364. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. CMLS 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Anthoney, N.; Foldi, I.; Hidalgo, A. Toll and Toll-like receptor signalling in development. Development 2018, 145, dev156018. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]
- Paerewijck, O.; Lamkanfi, M. The human inflammasomes. Mol. Asp. Med. 2022, 88, 101100. [Google Scholar] [CrossRef]
- Aminzadeh, A. Protective effect of tropisetron on high glucose induced apoptosis and oxidative stress in PC12 cells: Roles of JNK, P38 MAPKs, and mitochondria pathway. Metab. Brain Dis. 2017, 32, 819–826. [Google Scholar] [CrossRef]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Du, J.; Xi, L.; Lei, B.; Liu, H.; Yao, X. Structural requirements of isoquinolones as novel selective c-Jun N-terminal kinase 1 inhibitors: 2D and 3D QSAR analyses. Chem. Biol. Drug Des. 2011, 77, 248–254. [Google Scholar] [CrossRef]
- Masenga, S.K.; Kabwe, L.S.; Chakulya, M.; Kirabo, A. Mechanisms of Oxidative Stress in Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 7898. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, D.; Korhonen, L.; Eriksson, O.; Kõks, S. Recent Insights into the Role of Unfolded Protein Response in ER Stress in Health and Disease. Front. Cell Dev. Biol. 2017, 5, 48. [Google Scholar] [CrossRef]
- Cybulsky, A.V. Endoplasmic reticulum stress, the unfolded protein response and autophagy in kidney diseases. Nat. Rev. Nephrol. 2017, 13, 681–696. [Google Scholar] [CrossRef]
- Toma, L.; Sanda, G.M.; Niculescu, L.S.; Deleanu, M.; Stancu, C.S.; Sima, A.V. Caffeic acid attenuates the inflammatory stress induced by glycated LDL in human endothelial cells by mechanisms involving inhibition of AGE-receptor, oxidative, and endoplasmic reticulum stress. BioFactors 2017, 43, 685–697. [Google Scholar] [CrossRef]
- Ren, X.; Zhang, J. Study on the influence of salvianolic acid B on vSIRT1/SIRT3 signaling pathway in non-alcoholic fatty liver cell model. J. Qiqihar Med. Univ. 2022, 43, 2313–2318. [Google Scholar]
- Fang, L.; Wang, Y.; He, G. Anti-diabetic effect of salvianolic acid A and the possible mechanisms in rats with diabetes mellitus. Chin. J. New Drugs 2011, 20, 2063–2068. [Google Scholar]
- Kwon, E.Y.; Do, G.M.; Cho, Y.Y.; Park, Y.B.; Jeon, S.M.; Choi, M.S. Anti-atherogenic property of ferulic acid in apolipoprotein E-deficient mice fed Western diet: Comparison with clofibrate. Food Chem. Toxicol. 2010, 48, 2298–2303. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Anti-atherosclerotic Activity of Para Methoxy Cinnamic Acid in High Fat Diet Induced Hyperlipidemia Model Rats. Appl. Biochem. Biotechnol. 2022, 194, 1911–1924. [Google Scholar] [CrossRef]
- Ghaddar, B.; Gence, L.; Veeren, B.; Bringart, M.; Bascands, J.L.; Meilhac, O.; Diotel, N. Aqueous Extract of Psiloxylon mauritianum, Rich in Gallic Acid, Prevents Obesity and Associated Deleterious Effects in Zebrafish. Antioxidants 2022, 11, 1309. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.; Sun, R.; Fu, R.; Sun, Y.; Zhu, M.; Geng, Y.; Gao, D. Activation of TAF9 via Danshensu-Induced Upregulation of HDAC1 Expression Alleviates Non-alcoholic Fatty Liver Disease. Front. Pharmacol. 2021, 12, 775528. [Google Scholar] [CrossRef] [PubMed]
- Chao, P.C.; Hsu, C.C.; Yin, M.C. Anti-inflammatory and anti-coagulatory activities of caffeic acid and ellagic acid in cardiac tissue of diabetic mice. Nutr. Metab. 2009, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Jeon, S.M. Antihyperglycemic and antioxidant properties of caffeic acid in db/db mice. J. Pharmacol. Exp. Ther. 2006, 318, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Karthikesan, K.; Pari, L.; Menon, V.P. Antihyperlipidemic effect of chlorogenic acid and tetrahydrocurcumin in rats subjected to diabetogenic agents. Chem. Biol. Interact. 2010, 188, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T. Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients 2018, 11, 23. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Wang, Z.; Lam, K.L.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Zeng, S.; Lin, S. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci. Nutr. 2019, 7, 579–588. [Google Scholar] [CrossRef]
- Wang, D.; Hou, J.; Wan, J.; Yang, Y.; Liu, S.; Li, X.; Li, W.; Dai, X.; Zhou, P. Dietary chlorogenic acid ameliorates oxidative stress and improves endothelial function in diabetic mice via Nrf2 activation. J. Int. Med. Res. 2021, 49, 300060520985363. [Google Scholar] [CrossRef]
- Alarcón Yempén, R.E.; Venzel, R.; Paulino Campos, M.C.; de Oliveira, L.P.; Lins, R.V.D.; Pessoni, A.M.; Fanaro, G.B.; de Oliveira Souza, A.; Calaza, K.D.C.; de Brito Alves, J.L.; et al. Gut microbiota: A potential therapeutic target for management of diabetic retinopathy? Life Sci. 2021, 286, 120060. [Google Scholar] [CrossRef]
- Guan, L.; Long, H.; Ren, F.; Li, Y.; Zhang, H. A Structure-Activity Relationship Study of the Inhibition of α-Amylase by Benzoic Acid and Its Derivatives. Nutrients 2022, 14, 1931. [Google Scholar] [CrossRef]
- Yu, M.; Zhu, S.; Huang, D.; Tao, X.; Li, Y. Inhibition of starch digestion by phenolic acids with a cinnamic acid backbone: Structural requirements for the inhibition of α-amylase and α-glucosidase. Food Chem. 2024, 435, 137499. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yang, W.; Sun, W.; Chen, S.; Ye, X. Inhibition of porcine pancreatic α-amylase activity by chlorogenic acid. J. Funct. Foods 2019, 64, 103587. [Google Scholar] [CrossRef]
- Kaur, N.; Kumar, V.; Nayak, S.K.; Wadhwa, P.; Kaur, P.; Sahu, S.K. Alpha-amylase as molecular target for treatment of diabetes mellitus: A comprehensive review. Chem. Biol. Drug Des. 2021, 98, 539–560. [Google Scholar] [CrossRef]
- Wu, W.M.; Lu, L.; Long, Y.; Wang, T.; Liu, L.; Chen, Q.; Wang, R. Free radical scavenging and antioxidative activities of caffeic acid phenethyl ester (CAPE) and its related compounds in solution and membranes: A structure–activity insight. Food Chem. 2007, 105, 107–115. [Google Scholar] [CrossRef]
- Chen, X. Study on the Structure Activity Relationship and Antioxidant Mechanism of Phenolic Acid Antioxidant Activity; NUC: Taiyuan, China, 2020. [Google Scholar]
- Zhu, H.; Chen, S.; Hao, S.; Zhang, Z.; Wang, W.; Yao, S.J.B.-G.S. Double roles of hydroxycinnamic acid derivatives in protection against lysozyme oxidation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2006, 1760, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Siquet, C.; Paiva-Martins, F.; Lima, J.L.; Reis, S.; Borges, F. Antioxidant profile of dihydroxy- and trihydroxyphenolic acids-A structure–activity relationship study. Free Radic. Res. 2009, 40, 433–442. [Google Scholar] [CrossRef]
- Cheng, J.C.; Dai, F.; Zhou, B.; Yang, L.; Liu, Z.L.J.F.C. Antioxidant activity of hydroxycinnamic acid derivatives in human low density lipoprotein: Mechanism and structure-activity relationship. Food Chem. 2007, 104, 132–139. [Google Scholar] [CrossRef]
- Shan, W.; Chen, S.; Liang, W.; Wang, Y. Synthesis and antioxidant structure-activity relationship of hydroxycinnamic acid derivatives. Sci. Technol. Food Ind. 2017, 38, 287–291+332. [Google Scholar]
- Sawai, Y.; Moon, J.H.; Sakata, K.; Watanabe, N. Effects of structure on radical-scavenging abilities and antioxidative activities of tea polyphenols: NMR analytical approach using 1,1-diphenyl-2-picrylhydrazyl radicals. J. Agric. Food Chem. 2005, 53, 3598–3604. [Google Scholar] [CrossRef]
- Szeląg, M.G.; Urbaniak, A.; Bluyssen, H.A. A theoretical antioxidant pharmacophore for natural hydroxycinnamic acids. Open Chem. 2014, 13, 17–31. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Components | MEHHs | Pathway | Mechanism Pathway | Reference |
|---|---|---|---|---|---|
| 1 | Caffeic acid | Cirsium setosum (Willd.) MB.; Portulaca oleracea L.; Phyllanthus emblica L.; Citrus medica L’ | Regulates glucose metabolism | Inhibits α-amylase and α-glucosidase activity | [12,13,14] |
| 2 | Chlorogenic acid | Cirsium setosum (Willd.) MB.; | Regulates glucose metabolism | Inhibits α-amylase and α-glucosidase activity | [12] |
| 3 | Ferulic acid | Dolichos lablab L.; Dimocarpus longan Lour.; Hippophae rhamnoides L. | Regulates glucose metabolism | Inhibits α-amylase and α-glucosidase activity | [13,15,16] |
| 4 | Caffeic acid | Lonicera japonica Thunb; Zingiber officinale Rosc.; Lycium barbarum L. | Regulates glucose metabolism | Enhances the expression and translocation of GLUT4 and GLUT2 proteins | [17,18,19] |
| 5 | Chlorogenic acid | Lycium barbarum L. | Regulates glucose metabolism | Enhances the expression and translocation of GLUT4 proteins | [19] |
| 6 | Rosmarinic acid | Perilla frutescens (L.) Britt. (leaf) | Regulates glucose metabolism | Reduces PEPCK expression in the liver and increases GLUT4 expression in muscles | [20] |
| 7 | Ferulic acid | Hippophae rhamnoides L.; Hovenia dulcis Thunb.; Morus alba L. (leaf) | Regulates glucose metabolism | Inhibits glycogen phosphorylase, glucose-6-phosphatase, and fructose-1,6-diphosphatase activity Inhibits the protein expression of hepatic gluconeoxygenase, PEPCK, and G6Pase | [16,21,22] |
| 8 | Gallic acid | Lycium barbarum L. | Regulates glucose metabolism | Downregulates the expression of fructose-1,6-phosphatase, upregulate the expression of hexokinase, phosphofructokinase, and aldolase | [23] |
| 9 | Salvianolic acid B | Angelica sinensis (Oliv.) Diels. | Regulates glucose metabolism | Reduces the expression of AGEs within plaques | [24] |
| 10 | Protocatechuic acid | Prunella vulgaris L. | Regulates glucose metabolism | Reduces the levels of AGEs, glycosylated albumin and type IV collagen in STZ induced diabetes mice | [25] |
| 11 | Ferulic acid | Thunb.; Morus alba L. (leaf); Curcuma Longa L.; Kaempferia galanga L. | Regulates lipid metabolism | Regulates the AMPK α/SREBP1/ACC1 signaling pathway | [22,26,27] |
| 12 | Salvianolic acid B | Mentha haplocalyx Briq. | Regulates lipid metabolism | Regulates the AMPK pathway, enhances liver autophagy levels in ApoE−/− mice, reduces liver oxidative stress and inflammation levels, and alleviates liver damage | [28] |
| 13 | Rosmarinic acid | Prunella vulgaris L. | Regulates lipid metabolism | Regulates the AMPK/SREBP1c signaling pathway | [29] |
| 14 | Caffeic acid | Nelumbo nucifera Gaertn. (leaf); Chrysanthemum morifolium Ramat. | Regulates lipid metabolism | Inhibits the expression of SREBP1, Fas, ACC, and SCD1 in the liver tissue of obese mice | [30,31,32] |
| 15 | Chlorogenic acid | Cinnamomum cassia Presl; Zanthoxylum bungeanum Maxim. | Regulates lipid metabolism | Activates AMPK-activated protein kinase, inhibits HMGCoA activity, enhances CPT activity | [30,33,34] |
| 16 | p-Coumaric acid | Prunella vulgaris L. | Regulates lipid metabolism | Dose-dependent increase in AMPK phosphorylation and ACC phosphorylation in differentiated L6 skeletal muscle cells | [35] |
| 17 | Vanillic acid | Crataegus pinnatifida Bge. | Regulates lipid metabolism | Activates AMPK phosphorylation and inhibits ACC activity | [36] |
| 18 | Ferulic acid | Dolichos lablab L.; Kaempferia galanga L. | Regulates lipid metabolism | Inhibits the activity of hydroxyglutaryl-5-pyrophosphate dehydrogenase in the liver, increases the activity of β-oxidation gene CPT1A and the expression of PPAR α in liver tissue | [15,27,37] |
| 19 | p-Coumaric acid | Prunella vulgaris L. | Regulates lipid metabolism | Enhances the expression of CPT-1 mRNA and PPAR α | [35] |
| 20 | Caffeic acid | Angelica sinensis (Oliv.) Diels. | Regulates lipid metabolism | Enhances the activity of fatty acid β oxidation and stimulates the expression of liver PPAR α | [30] |
| 21 | Chlorogenic acid | Prunella vulgaris L.; Angelica sinensis (Oliv.) Diels. | Regulates lipid metabolism | Enhances the activity of fatty acid β oxidation and stimulates the expression of liver PPAR α. Increases the expression level of genes involved in fatty acid metabolism | [30,38] |
| 22 | Protocatechuic acid | Lycium barbarum L. | Regulates lipid metabolism | Inhibits the expression of AR, SDH, GLI, PKC, PPAR-γ, and RAGE | [39] |
| 23 | Rosmarinic acid | Vigna umbellata Ohwi et Ohashi | Regulates lipid metabolism | Regulates the YAP1/TAZ-PPAR γ/PGC-1 α signaling pathway | [40] |
| 24 | Danshensu | Mentha haplocalyx Briq.; Prunella vulgaris L. | Regulates lipid metabolism | Increases the levels of LCAT and CYP7A1 genes and proteins in liver tissue, upregulates apolipoprotein apo AI and downregulates apo B | [41] |
| 25 | Caffeic acid | Phyllanthus emblica L.; Mentha haplocalyx Briq. | Regulates insulin signaling and improves insulin sensitivity | Improves the function and morphology of pancreatic β cells in type II diabetes rats | [14,42] |
| 26 | Chlorogenic acid | Lycium barbarum L.; Prunella vulgaris L. | Regulates insulin signaling and improves insulin sensitivity | Upregulates protein expression in the IR, IRS-1, PI3K, and Akt pathways, inhibits JNK pathway activation, and inhibits autophagy | [23,43] |
| 27 | Salvianolic acid B | Angelica sinensis (Oliv.) Diels.; Mentha haplocalyx Briq. | Regulates insulin signaling and improves insulin sensitivity | Inhibits the activation levels of JNK and NF-κ B in pancreatic tissue, downregulates the expression of pro-apoptotic proteins Bax and Bim, upregulates the level of anti-apoptotic protein Bcl-2, and significantly reduces the activity of Caspase-9 and Caspase-3 | [44,45] |
| 28 | Caffeic acid | Cirsium setosum (Willd.) MB.; Portulaca oleracea L.; Phyllanthus emblica L.; | Inhibits inflammatory responses | Reduces the expression of TLR4 in the liver and inhibits the activation of phosphorylated NF-κ B p65 in liver tissue | [46,47] |
| 29 | Danshensu | Mentha haplocalyx Briq.; Prunella vulgaris L. | Inhibits inflammatory responses | Downregulates TLR2 and TLR4, p-I-κ B, and NF-κ B p65 protein expression | [48] |
| 30 | Chlorogenic acid | Citrus medica L.; Prunus armeniaca L.; Hippophae rhamnoides L.; | Inhibits inflammatory responses | Reverses the TLR4 signaling pathway induced by HFD Inhibits autophagy through JNK pathway inactivation | [49,50,51] |
| 31 | Salicylic acid | Cichorium intybus L.; Hippophae rhamnoides L.; | Inhibits inflammatory responses | Downregulates the expression of liver inflammation genes TLR4, MYD88, NF-κ B, and upregulates the expression of fatty acid oxidation genes Ppar α, Acsl, Cpt1, and Cpt2 | [52] |
| 32 | Rosmarinic acid | Perilla frutescens (L.) Britt. (leaf) | Inhibits inflammatory responses | Inhibits NF-κ B and MAPK expression | [39] |
| 33 | Salvianolic acid B | Angelica sinensis (Oliv.) Diels.; Mentha haplocalyx Briq. | Inhibits inflammatory responses | Reduces the expression levels of NF-κ B p65, IL-6, and TNF-α in the liver | [28] |
| 34 | Protocatechuic acid | Hippophae rhamnoides L.; Hordeum vulgare L.; Ziziphus jujuba Mill. | Inhibits inflammatory responses | Reduces NF-κ B binding activity. By upregulating MERTK and MAPK 3/1, the activation of NF-κ B in macrophages is inhibited | [5,53,54] |
| 35 | Ferulic acid | Laminaria japonica Aresch. | Inhibits inflammatory responses | Significantly inhibits the expression of NF-κ B and significantly reduces the average concentration of MDA | [55] |
| 36 | Vanillic acid | Crataegus pinnatifida Bge. | Inhibits inflammatory responses | Downregulates NF-κ B and exerts anti-inflammatory effects | [36] |
| 37 | Salvianolic acid A | Angelica sinensis (Oliv.) Diels. | Inhibits inflammatory responses | Inhibits the expression of NLRP3 inflammasome and reduces inflammatory response Regulates the TXNIP-NLRP3 and TXNIP ChREBP pathways | [56,57] |
| 38 | Chlorogenic acid | Lonicera japonica Thunb. | Inhibition of oxidative stress | Increases the expression of Nrf2 and its downstream target proteins HO-1, NQO1, and GPx1 | [58] |
| 39 | p-Coumaric acid | Dimocarpus longan Lour. | Inhibition of oxidative stress | Upregulates the expression of Nrf2, SOD, HO-1, and NQO-1 | [59] |
| 40 | Caffeic acid | Nelumbo nucifera Gaertn. (leaf); Chrysanthemum morifolium Ramat. | Inhibition of oxidative stress | Significantly reduces the protein levels of ER stress markers BIP, ATF4, CHOP, GADD34, and XBP-1 Regulates the UPR pathways PERK, IRE1 α, and ATF6 α | [32,60,61] |
| 41 | Caffeic acid | Nelumbo nucifera Gaertn. (leaf); Chrysanthemum morifolium Ramat. | Regulates intestinal flora | Increases the ileum and colon OUT of HFD mice, restores the richness and diversity of the microbiota | [31] |
| 42 | Chlorogenic acid | Cirsium setosum (Willd.) MB.; Crataegus pinnatifida Bge.; Portulaca oleracea L. | Regulates intestinal flora | Increases the expression of tight junction proteins Occludin and ZO-1 in intestinal tissue, improves intestinal mucosal barrier, and reverses gut microbiota dysbiosis caused by HFD | [49,62,63] |
| 43 | Ferulic acid | Kaempferia galanga L. | Regulates intestinal flora | Specifically reduces the relative abundance of Bacteroides, Erysipella and Ileum, which are positively related to the blood lipid level of atherosclerosis mice | [26] |
| 44 | Protocatechuic acid | Phyllanthus emblica L.; | Regulates intestinal flora | Protects the integrity of the mucosal layer of the ileum, reduces the infiltration of Gram-negative and positive bacteria in the intestinal mucosal tissue, and significantly increases the expression of ZO-1 protein in the ileum tissue | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Zhang, Z.; Xie, J.; Yu, J.; Xiong, S.; Xiang, F.; Ma, X.; Yang, C.; Lin, L. Research Progress on the Mechanism for Improving Glucose and Lipid Metabolism Disorders Using Phenolic Acid Components from Medicinal and Edible Homologous Plants. Molecules 2024, 29, 4790. https://doi.org/10.3390/molecules29204790
Sun M, Zhang Z, Xie J, Yu J, Xiong S, Xiang F, Ma X, Yang C, Lin L. Research Progress on the Mechanism for Improving Glucose and Lipid Metabolism Disorders Using Phenolic Acid Components from Medicinal and Edible Homologous Plants. Molecules. 2024; 29(20):4790. https://doi.org/10.3390/molecules29204790
Chicago/Turabian StyleSun, Miao, Zhimin Zhang, Jingchen Xie, Jiahui Yu, Suhui Xiong, Feng Xiang, Xinyi Ma, Chen Yang, and Limei Lin. 2024. "Research Progress on the Mechanism for Improving Glucose and Lipid Metabolism Disorders Using Phenolic Acid Components from Medicinal and Edible Homologous Plants" Molecules 29, no. 20: 4790. https://doi.org/10.3390/molecules29204790
APA StyleSun, M., Zhang, Z., Xie, J., Yu, J., Xiong, S., Xiang, F., Ma, X., Yang, C., & Lin, L. (2024). Research Progress on the Mechanism for Improving Glucose and Lipid Metabolism Disorders Using Phenolic Acid Components from Medicinal and Edible Homologous Plants. Molecules, 29(20), 4790. https://doi.org/10.3390/molecules29204790