
The Effect of Different Surfactants and Polyelectrolytes on Nano-Vesiculation of Artificial and Cellular Membranes

 , , ,
, , ,  and
and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. CMC of Surfactants
2.2. Size and Shape of Pure POPC Liposomes
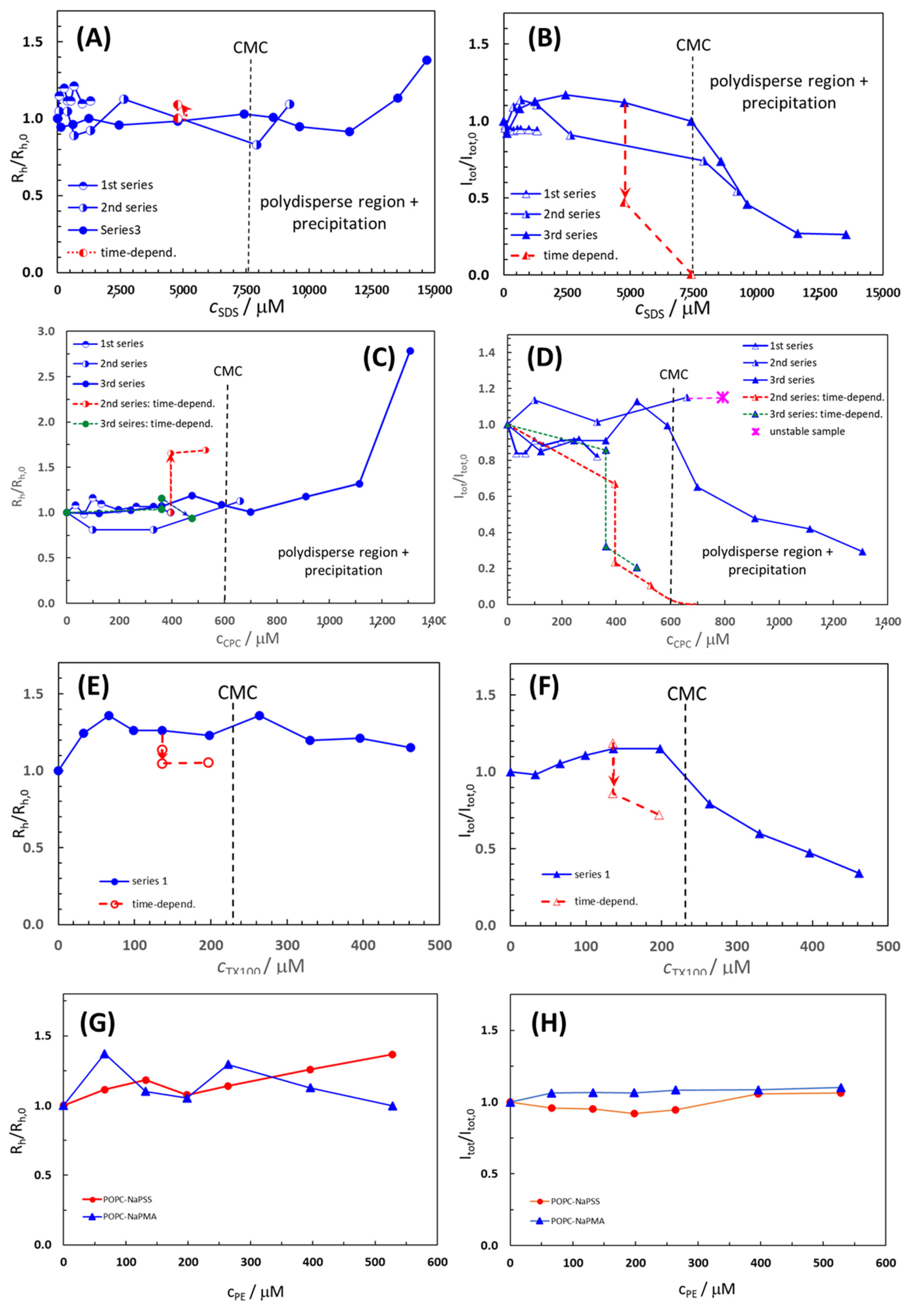
2.3. Effect of Surfactants and Polyelectrolytes on Liposome Size, Shape, and Stability
2.4. Effect of Temperature on Size and Stability of Liposomes with Added Surfactants and Polyelectrolytes
2.5. Calorimetric Measurements of POPC/Surfactant and POPC/Polyelectrolyte Colloids
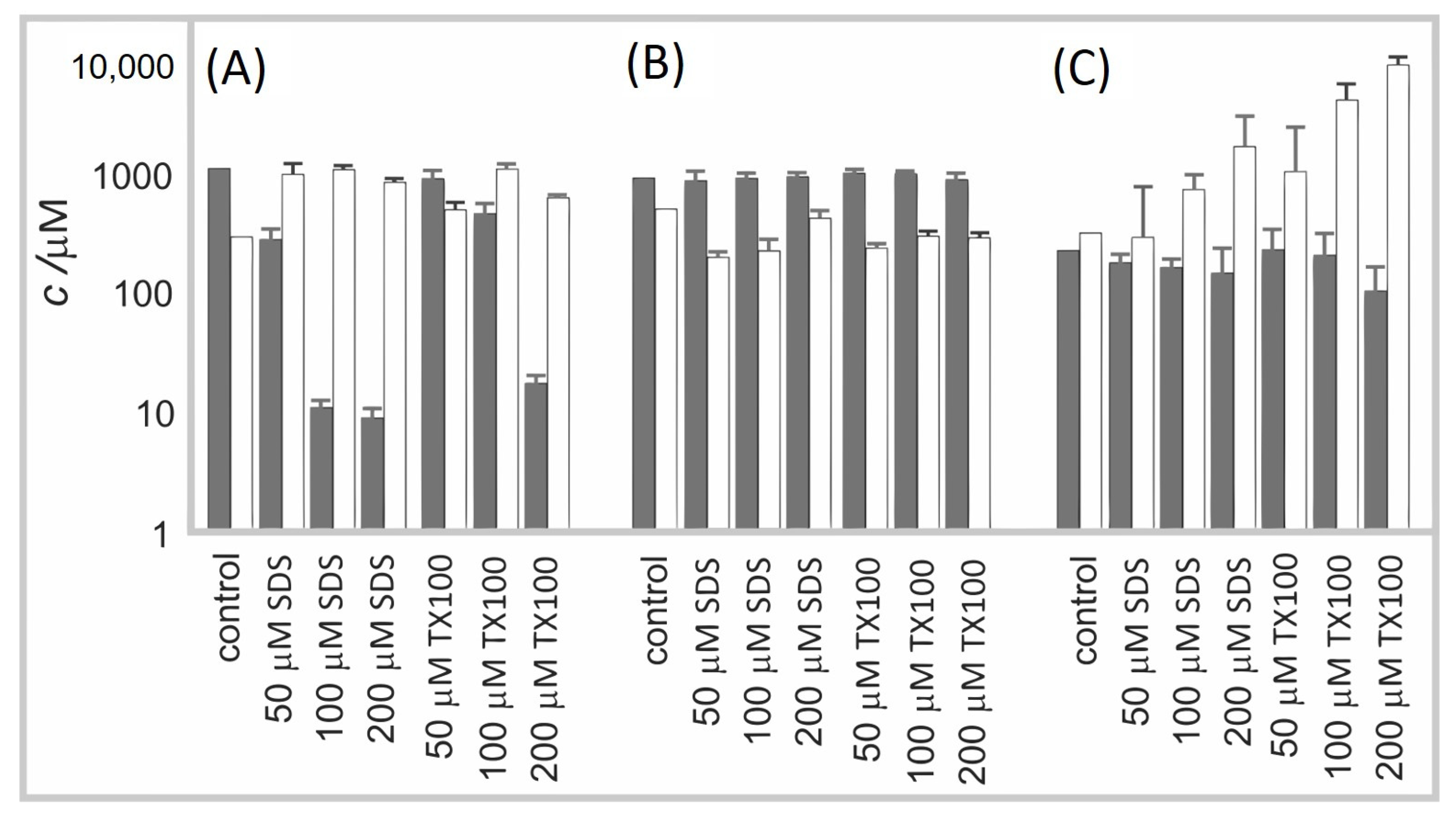
2.6. Effect of SDS and TX100 on Vesiculation of Membranes of Erythrocytes and Microalgae
3. Materials and Methods
3.1. Materials
3.1.1. Chemicals
3.1.2. Preparation of TRIS Buffer
3.1.3. Preparation of the Stock POPC Liposome Suspension
3.1.4. Preparation of Surfactant (S) and Polyelectrolyte (PE) Solutions
3.1.5. Preparation of POPC Suspensions with Added S or PE for Light Scattering Measurements
3.1.6. Cultures of Microalgae
3.1.7. Preparation of Cells for the Study of Effects of Surfactants
3.1.8. Preparation of Extracellular Vesicles from Erythrocytes
3.2. Methods
3.2.1. Conductivity Measurements
3.2.2. Isothermal Titration Calorimetry (ITC) Measurements
3.2.3. Light Scattering Measurements
3.2.4. Flow Cytometry (FCM)
3.2.5. Scanning Electron Microscopy (SEM)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welsh, A.J.; Goberdhan, C.D.; O′Driscoll, L.; Théry, C.; Witwer, W.K. MISEV 2023: An Updated Guide to EV Research and Applications. J. Extracell. Vesicles 2024, 13, e12416. [Google Scholar] [CrossRef]
- Gaurav, I.; Thakur, A.; Iyaswamy, A.; Wang, X.; Chen, X.; Yang, Z. Factors Affecting Extracellular Vesicles Based Drug Delivery Systems. Molecules 2021, 26, 1544. [Google Scholar] [CrossRef]
- Rajput, A.; Varshney, A.; Bajaj, R.; Pokharkar, V. Exosomes as New Generation Vehicles for Drug Delivery: Biomedical Applications and Future Perspectives. Molecules 2022, 27, 7289. [Google Scholar] [CrossRef]
- Sędzik, M.; Rakoczy, K.; Sleziak, J.; Kisiel, M.; Kraska, K.; Rubin, J.; Łuniewska, W.; Choromańska, A. Comparative Analysis of Exosomes and Extracellular Microvesicles in Healing Pathways: Insights for Advancing Regenerative Therapies. Molecules 2024, 29, 3681. [Google Scholar] [CrossRef]
- Pasarin, D.; Ghizdareanu, A.-I.; Enascuta, C.E.; Matei, C.B.; Bilbie, C.; Paraschiv-Palada, L.; Veres, P.-A. Coating Materials to Increase the Stability of Liposomes. Polymers 2023, 15, 782. [Google Scholar] [CrossRef]
- Isomaa, B.; Hägerstrand, H.; Paatero, G. Shape transformations induced by amphiphiles in erythrocytes. Biochim. Biophys. Acta. 1987, 899, 93–103. [Google Scholar] [CrossRef]
- Isomaa, B.; Hägerstrand, H. Effects of nonionic amphiphiles at sublytic concentrations on the erythrocyte membrane. Cell Biochem. Funct. 1988, 6, 183–190. [Google Scholar] [CrossRef]
- Pandur, Z.; Penic, S.; Iglic, A.; Kralj-Iglic, V.; Stopar, D.; Drab, M. Surfactin molecules with a cone-like structure promote the formation of membrane domains with negative spontaneous curvature and induce membrane invaginations. J. Coll. Int. Sci. 2023, 650B, 1193–1200. [Google Scholar] [CrossRef]
- Arrigler, V.; Kogej, K.; Majhenc, J.; Svetina, S. Interaction of cetylpyridinium chloride with giant lipid vesicles. Langmuir 2005, 21, 7653–7661. [Google Scholar] [CrossRef]
- Jicsinszky, L.; Martina, K.; Cravotto, G. Cyclodextrins in the antiviral therapy. J. Drug. Deliv. Sci. Technol. 2021, 64, 102589. [Google Scholar] [CrossRef]
- Fromm-Dornieden, C.; Rembe, J.D.; Schäfer, N.; Böhm, J.; Stuermer, E.K. Cetylpyridinium chloride and miramistin as antiseptic substances in chronic wound management—prospects and limitations. J. Med. Microbiol. 2015, 64, 407–414. [Google Scholar] [CrossRef]
- Alvarez, D.M.; Duarte, L.F.; Corrales, N.; Smith, P.C.; González, P.A. Cetylpyridinium chloride blocks herpes simplex virus replication in gingival fibroblasts. Antivir. Res. 2020, 179, 104818. [Google Scholar] [CrossRef]
- Pérez-Errázuriz, S.; Velasco-Ortega, E.; Jiménez-Guerra, A.; Aguilera-Navarro, E. Cetylpyridinium chloride as a tool against COVID-19. Int. J. Odontostomat. 2021, 15, 27–308. [Google Scholar] [CrossRef]
- Parker, W.; Song, P.S. Protein structures in SDS micelle-protein complexes. Biophys, J. 1992, 61, 1435–1439. [Google Scholar] [CrossRef]
- Koley, D.; Bard, A.J. Triton X-100 concentration effects on membrane permeability of a single HeLa cell by scanning electrochemical microscopy (SECM). Proc. Natl. Acad. Sci. USA 2010, 107, 16783–16787. [Google Scholar] [CrossRef]
- Borner, M.M.; Schneider, E.; Pirnia, F.; Sartor, O.; Trepel, J.B.; Myers, C.E. The detergent Triton X-100 induces a death pattern in human carcinoma cell lines that resembles cytotoxic lymphocyte-induced apoptosis. FEBS Lett. 1994, 353, 129–132. [Google Scholar] [CrossRef]
- Dayeh, V.R.; Chow, S.L.; Schirmer, K.; Lynn, D.H.; Bols, N.C. Evaluating the toxicity of Triton X-100 to protozoan, fish, and mammalian cells using fluorescent dyes as indicators of cell viability. Ecotoxicol Env. Saf. 2004, 57, 375–382. [Google Scholar] [CrossRef]
- Yasuhara, K.; Morigaki, K. Creation of supramolecular biomembrane by the bottom-up self-assembly: Where material science meets biophysics. Biophys. Physicobiol. 2022, 19, 190043. [Google Scholar] [CrossRef]
- Yasuhara, K.; Arakida, J.; Ravula, T.; Ramadugu, S.; Sahoo, B.; Kikuchi, J.; Ramamoorthy, A. Spontaneous lipid nanodisc fomation by amphiphilic polymethacrylate copolymers. J. Am. Chem. Soc. 2017, 139, 18657–18663. [Google Scholar] [CrossRef]
- Coleman, R.; Holdsworth, G. Effects of detergents on erythrocyte membranes: Different patterns of solubilization of the membrane proteins by dihydroxy and trihydroxy bile salts. Biochem. Soc. Trans. 1975, 3, 747–748. [Google Scholar] [CrossRef]
- Škerjanc, J.; Kogej, K.; Vesnaver, G. Polyelectrolyte-surfactant interactions, Enthalpy of binding of dodecyl- and cetylpyridinium cations to poly(styrenesulfonate) anion. J. Phys. Chem. 1988, 92, 6382–6385. [Google Scholar] [CrossRef]
- Kogej, K. Association and structure formation in oppositely charged polyelectrolyte-surfactant mixtures. Adv. Colloid Interface Sci. 2010, 158, 68–83. [Google Scholar] [CrossRef]
- Božič, D.; Sitar, S.; Junkar, I.; Štukelj, R.; Pajnič, M.; Žagar, E.; Kralj-Iglič, V.; Kogej, K. Viscosity of Plasma as a Key Factor in Assessment of Extracellular Vesicles by Light Scattering. Cell 2019, 8, 1046. [Google Scholar] [CrossRef]
- Kogej, K. Thermodynamic analysis of the conformational transition in aqueous solutions of isotactic and atactic poly(methacrylic acid) and the hydrophobic effect. Polymers 2016, 8, 168. [Google Scholar] [CrossRef]
- Medoš, Ž.; Plechkova, N.V.; Friesen, S.; Buchner, R.; Bešter-Rogač, M. Insight into the hydration of cationic surfactants: A thermodynamic and dielectric study of functionalized quaternary ammonium chlorides. Langmuir 2019, 35, 3759–3772. [Google Scholar] [CrossRef]
- Kralj-Iglič, V.; Pocsfalvi, G.; Mesarec, L.; Šuštar, V.; Hägerstrand, H.; Iglič, A. Minimizing isotropic and deviatoric membrane energy—An unifying formation mechanism of different cellular membrane nanovesicle types. PLoS ONE 2020, 15, e0244796. [Google Scholar] [CrossRef]
- Picciotto, S.; Barone, M.E.; Fierli, D.; Aranyos, A.; Adamo, G.; Božič, D.; Romancino, D.P.; Stanly, C.; Parkes, R.; Morsbach, S.; et al. Isolation of extracellular vesicles from microalgae: Towards the production of sustainable and natural nanocarriers of bioactive compounds. Biomater. Sci. 2021, 9, 2917–2930. [Google Scholar] [CrossRef]
- Adamo, G.; Fierli, D.; Romancino, D.P.; Picciotto, S.; Barone, M.E.; Aranyos, A.; Božič, D.; Morsbach, S.; Raccosta, S.; Stanly, C.; et al. Nanoalgosomes: Introducing extracellular vesicles produced by microalgae. J. Extracell. Vesicles 2021, 10, e12081. [Google Scholar] [CrossRef]
- Božič, D.; Hočevar, M.; Jeran, M.; Kisovec, M.; Bedina Zavec, A.; Romolo, A.; Škufca, D.; Podobnik, M.; Kogej, K.; Iglič, A.; et al. Ultrastructure and stability of cellular nanoparticles isolated from Phaeodactylum tricornutum and Dunaliella tertiolecta conditioned media. Open Res Eur. 2022, 2, 121. [Google Scholar] [CrossRef]
- Romolo, A.; Jan, Z.; Bedina Zavec, A.; Kisovec, M.; Arrigler, V.; Spasovski, V.; Podobnik, M.; Iglič, A.; Pocsfalvi, G.; Kogej, K.; et al. Assessment of Small Cellular Particles from Four Different Natural Sources and Liposomes by Interferometric Light Microscopy. Int. J. Mol. Sci. 2022, 23, 15801. [Google Scholar] [CrossRef]
- Štibler, U.; Božič, D.; Hočevar, M.; Jeran, M.; Touzet, N.; Manno, M.; Pocsfalvi, G.; Bongiovanni, A.; Iglič, A.; Kralj-Iglič, V. Toxicity of surfactants sodium dodecyl sulphate and TRITON X-100 to marine microorganisms. Proc. Socrat. Lect. 2020, 3, 55–66. [Google Scholar]
- Invally, K.; Ju, L.K. Biolytic effect of rhamnolipid biosurfactant and dodecyl sulfate against Phagotrophic alga Ochromonas danica. J. Surfactants Deterg. 2017, 20, 1161–1171. [Google Scholar] [CrossRef]
- Masakorala, K.; Turner, A.; Brown, M.T. Toxicity of synthetic surfactants to the marine macroalga, Ulva lactuca. Water Air Soil Pollut. 2011, 218, 283–291. [Google Scholar] [CrossRef]
- Cserháti, T.; Forgács, E.; Oros, G. Biological activity and environmental impact of anionic surfactants. Environ Int. 2002, 28, 337–348. [Google Scholar] [CrossRef]
- Motin, M.A.; Hafiz, M.K.M.S.; Reza, N.; Islam, M.A.; Yousuf, M.A. Effect of Sodium Dodecyl Sulfate on Volumetric Properties of Methanol Ethanol n-Propanol and iso-Propanol at (298.15–323.15) K. Salam Dhaka. Univ. J. Sci. 2012, 60, 129–136. [Google Scholar] [CrossRef]
- Tiller, G.; Mueller, T.; Dockter, M.; Struve, W. Hydrogenation of Triton X-100 eliminates its fluorescence and ultraviolet light absorption while preserving its detergent properties. Anal. Biochem. 1984, 141, 262–266. [Google Scholar] [CrossRef]
- Chen, M.; Zeng, Z.; Qu, X.; Tang, Y.; Long, Q.; Feng, X. Biocompatible anionic polyelectrolyte for improved liposome based gene transfection. Int. J. Pharm. 2015, 490, 173–179. [Google Scholar] [CrossRef]
- Tasi, L.M.; Liu, D.Z.; Chen, W.Y. Microcalorimetric investigation of the interaction of polysorbate surfactants with unilamellar phosphatidylcholines liposomes. Coll. Surf. A Physicochem. Eng. Asp. 2003, 213, 7–14. [Google Scholar] [CrossRef]
- Kawakami, K.; Nishihara, Y.; Hirano, K. Effect of Hydrophilic Polymers on Physical Stability of Liposome Dispersions. J. Phys. Chem. B 2001, 105, 2374–2385. [Google Scholar] [CrossRef]
- Chern, C.S.; Chiu, H.C.; Yang, Y.S. Interactions between nonionic Triton X surfactants and cholesterol-containing phosphatidylcholine liposomes. J. Coll. Int. Sci. 2006, 302, 335–340. [Google Scholar] [CrossRef]
- Tesson, B.; Gennet, M.J.; Fernandez, V.; Degand, S. Surface Chemical Composition of Diatoms. Chem. Bio. Chem. 2009, 10, 1915. [Google Scholar] [CrossRef]
- Tinaïg, L.C.; Carlos, U.; Lalia, M.; William, H. New structural insights into the cell-wall polysaccharide of the diatom Phaeodactylum tricornutum. Algal Res. 2017, 26, 172–179. [Google Scholar]
- Chen, H.; Jiang, J.-G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Zetsche, E.M.; Meysman, F.J.R. Dead or alive? Viability assessment of micro- and mesoplankton. J. Plankton Res. 2012, 34, 493–509. [Google Scholar] [CrossRef]
- Wright, D.A.; Gensemer, R.W.; Mitchelmore, C.L.; Stubblefield, W.A.; van Genderen, E.; Dawson, R.; Orano-Dawson, C.E.; Bearr, J.S.; Mueller, R.A.; Cooper, W.J. Shipboard trials of an ozone-based ballast water treatment system. Mar. Pollut. Bull. 2010, 60, 1571–1583. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Li, X.; Gong, J. Assays of cell viability: Discrimination of cells dying by apoptosis. Methods Cell Biol. 1994, 41, 15–38. [Google Scholar]
- Tellinghuisen, J. Calibration in isothermal titration calorimetry: Heat and cell volume from heat of dilution of NaCl(aq). Anal. Biochem. 2007, 360, 47–55. [Google Scholar] [CrossRef]
- Schärtl, W. Light Scattering from Polymer Solutions and Nanoparticle Dispersions; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Qazi, M.J.; Liefferink, R.W.; Schlegel, S.J.; Backus, E.H.G.; Bonn, D.; Shahidzadeh, N. Influence of Surfactants on Sodium Chloride Crystallization in Confinement. Langmuir ACS J. Surf. Colloids 2017, 33, 4260–4268. [Google Scholar] [CrossRef]
- Brown, W. Dynamic Light Scattering: The Method and Some Application; Clarendon Press: Oxford, UK, 1993. [Google Scholar]
- Hriberšek, P.; Kogej, K. Tacticity and counterion modulated temperature response of weak polyelectrolytes: The case of poly(methacrylic acid) stereoisomers in aqueous solutions. Macromolecules 2019, 52, 7028–7041. [Google Scholar] [CrossRef]
- Sitar, S.; Aseyev, V.; Kogej, K. Microgel-like aggregates of isotactic and atactic poly(methacrylic acid) chains in aqueous alkali chloride solutions as evidenced by ligh scattering. Soft Matter. 2014, 10, 7712–7722. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zagorc, U.; Božič, D.; Arrigler, V.; Medoš, Ž.; Hočevar, M.; Romolo, A.; Kralj-Iglič, V.; Kogej, K. The Effect of Different Surfactants and Polyelectrolytes on Nano-Vesiculation of Artificial and Cellular Membranes. Molecules 2024, 29, 4590. https://doi.org/10.3390/molecules29194590
Zagorc U, Božič D, Arrigler V, Medoš Ž, Hočevar M, Romolo A, Kralj-Iglič V, Kogej K. The Effect of Different Surfactants and Polyelectrolytes on Nano-Vesiculation of Artificial and Cellular Membranes. Molecules. 2024; 29(19):4590. https://doi.org/10.3390/molecules29194590
Chicago/Turabian StyleZagorc, Urška, Darja Božič, Vesna Arrigler, Žiga Medoš, Matej Hočevar, Anna Romolo, Veronika Kralj-Iglič, and Ksenija Kogej. 2024. "The Effect of Different Surfactants and Polyelectrolytes on Nano-Vesiculation of Artificial and Cellular Membranes" Molecules 29, no. 19: 4590. https://doi.org/10.3390/molecules29194590
APA StyleZagorc, U., Božič, D., Arrigler, V., Medoš, Ž., Hočevar, M., Romolo, A., Kralj-Iglič, V., & Kogej, K. (2024). The Effect of Different Surfactants and Polyelectrolytes on Nano-Vesiculation of Artificial and Cellular Membranes. Molecules, 29(19), 4590. https://doi.org/10.3390/molecules29194590