

Saikosaponin A Recovers Impaired Filaggrin Levels in Inflamed Skin by Downregulating the Expression of FRA1 and c-Jun

 , , and
, , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
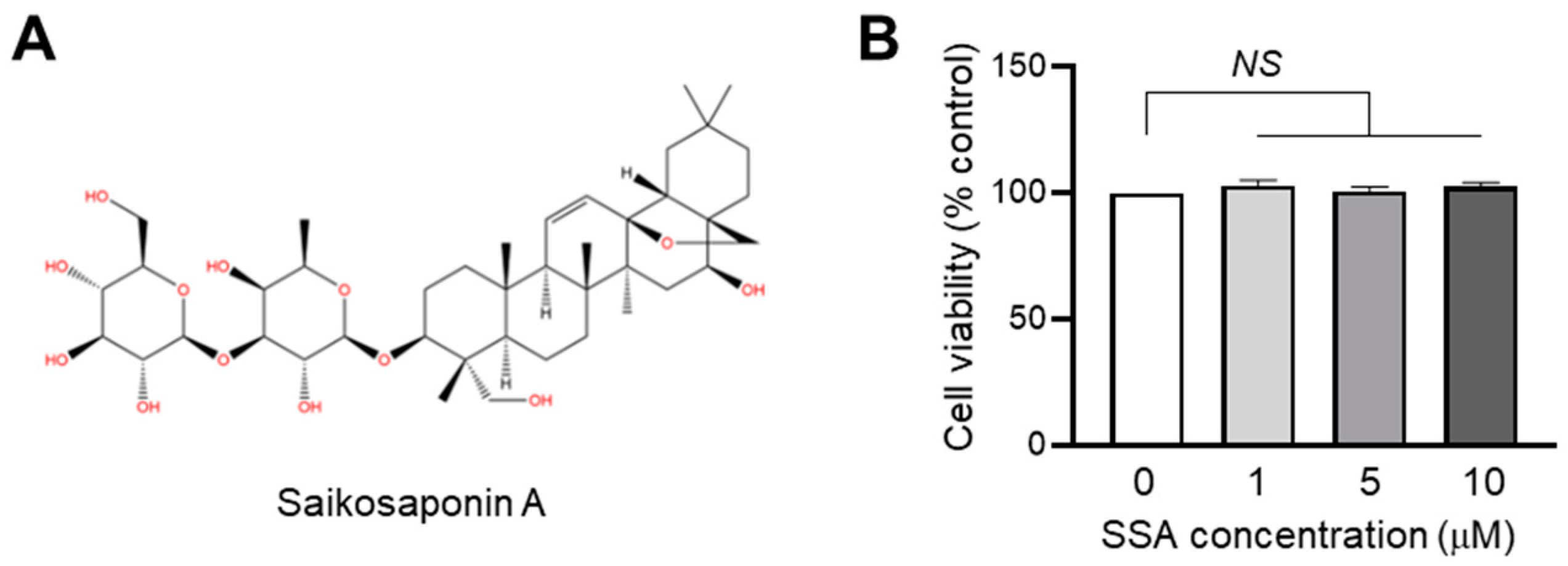
2.1. Effect of SSA on Cytotoxicity in HaCaT Keratinocytes
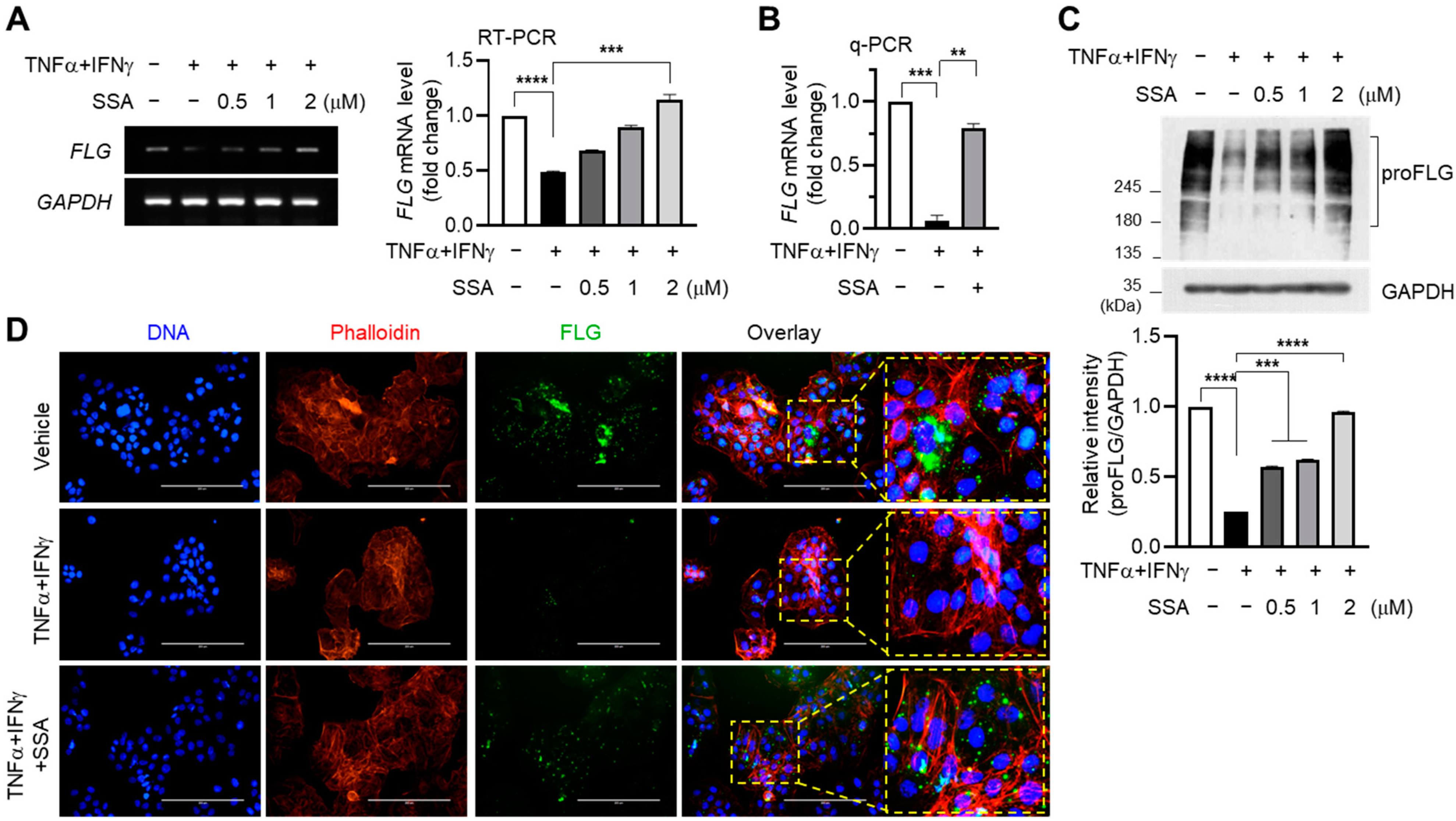
2.2. Inhibitory Effect of SSA on TNFα + IFNγ-Induced Suppression of FLG Expression in HaCaT Keratinocytes
2.3. SSA Inhibits TNFα + IFNγ-Induced Repression of FLG Transcription
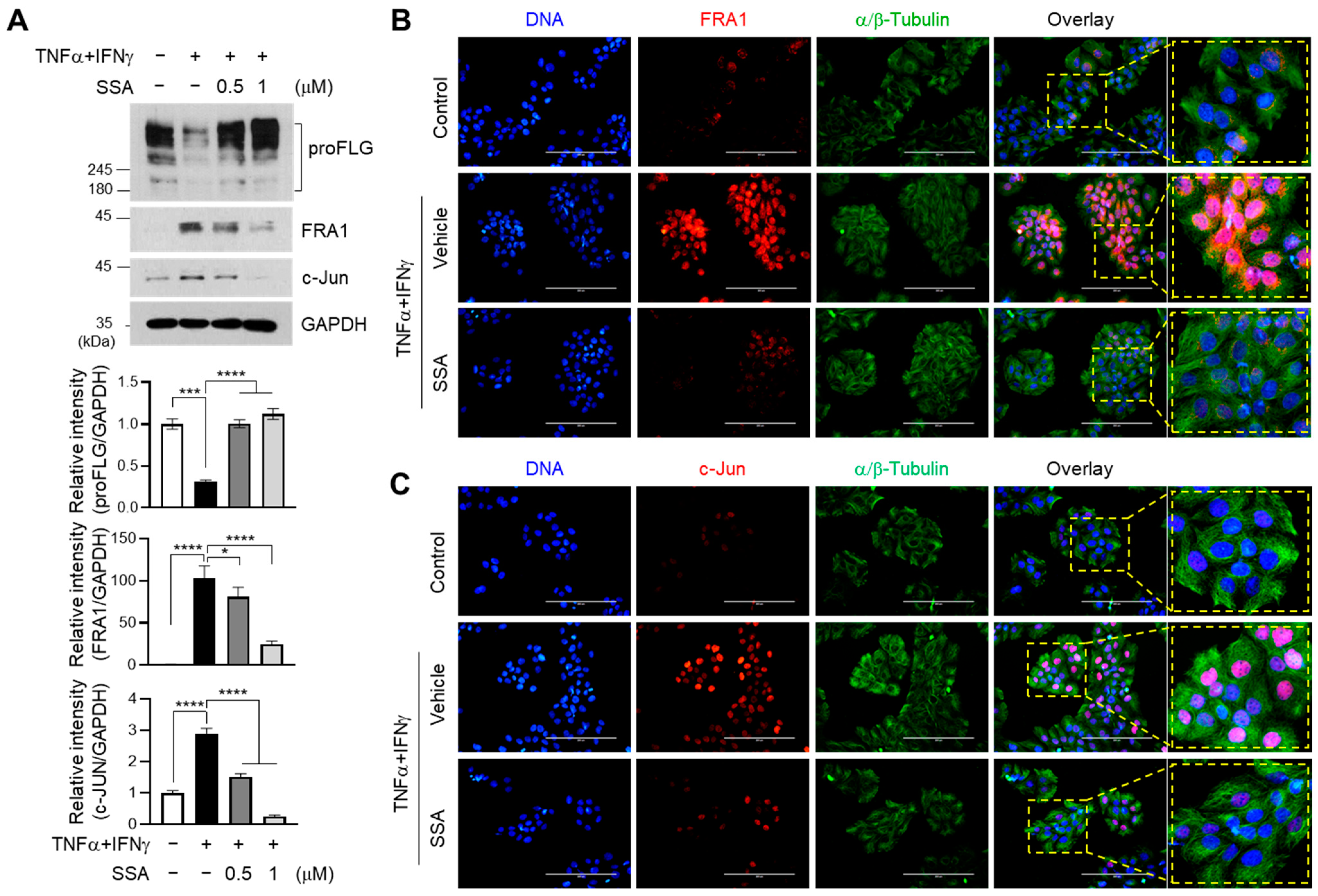
2.4. SSA Inhibits TNFα + IFNγ-Induced Expression of c-Jun and FRA1
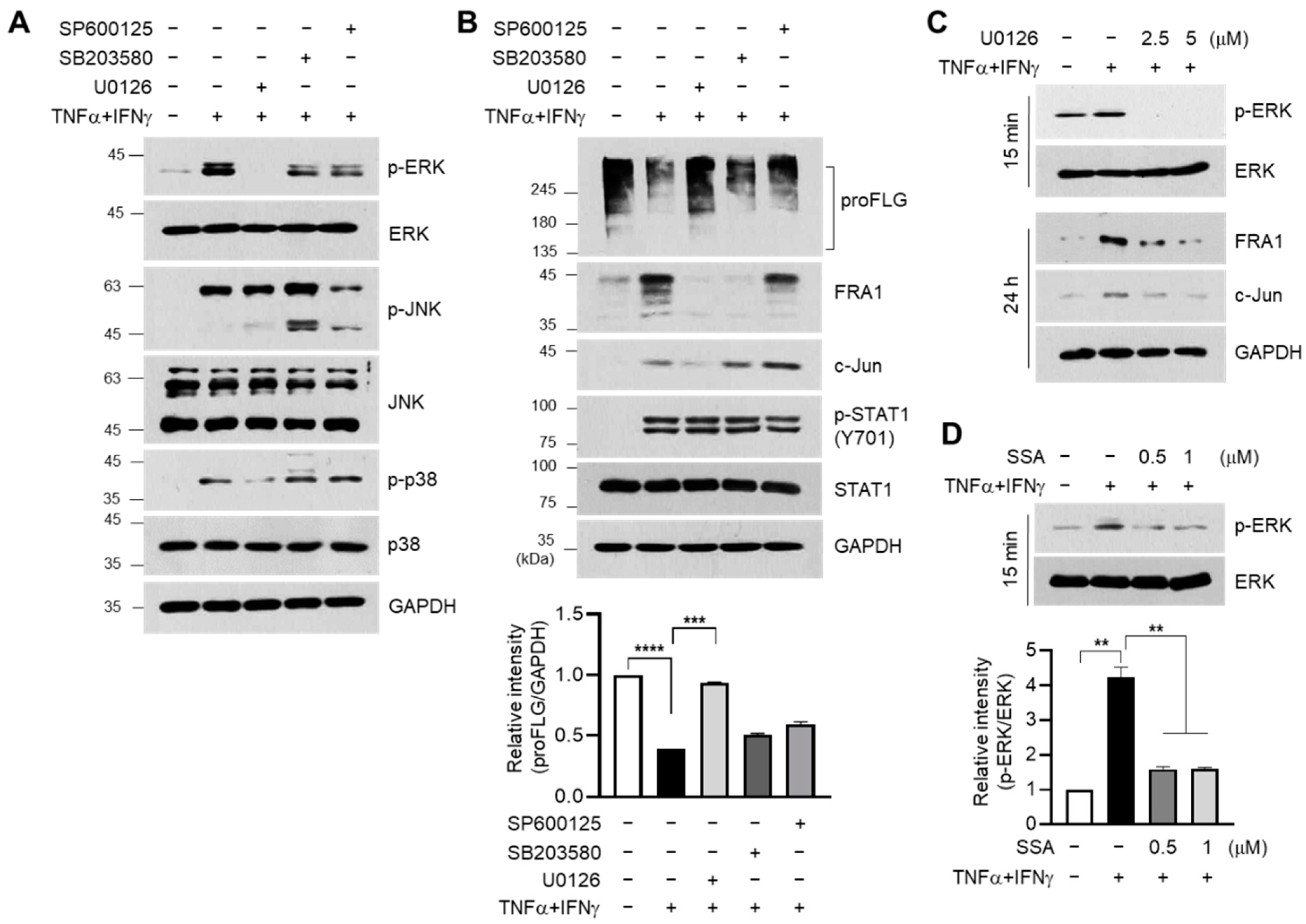
2.5. SSA Abrogates TNFα + IFNγ-Induced Activation of ERK Signaling
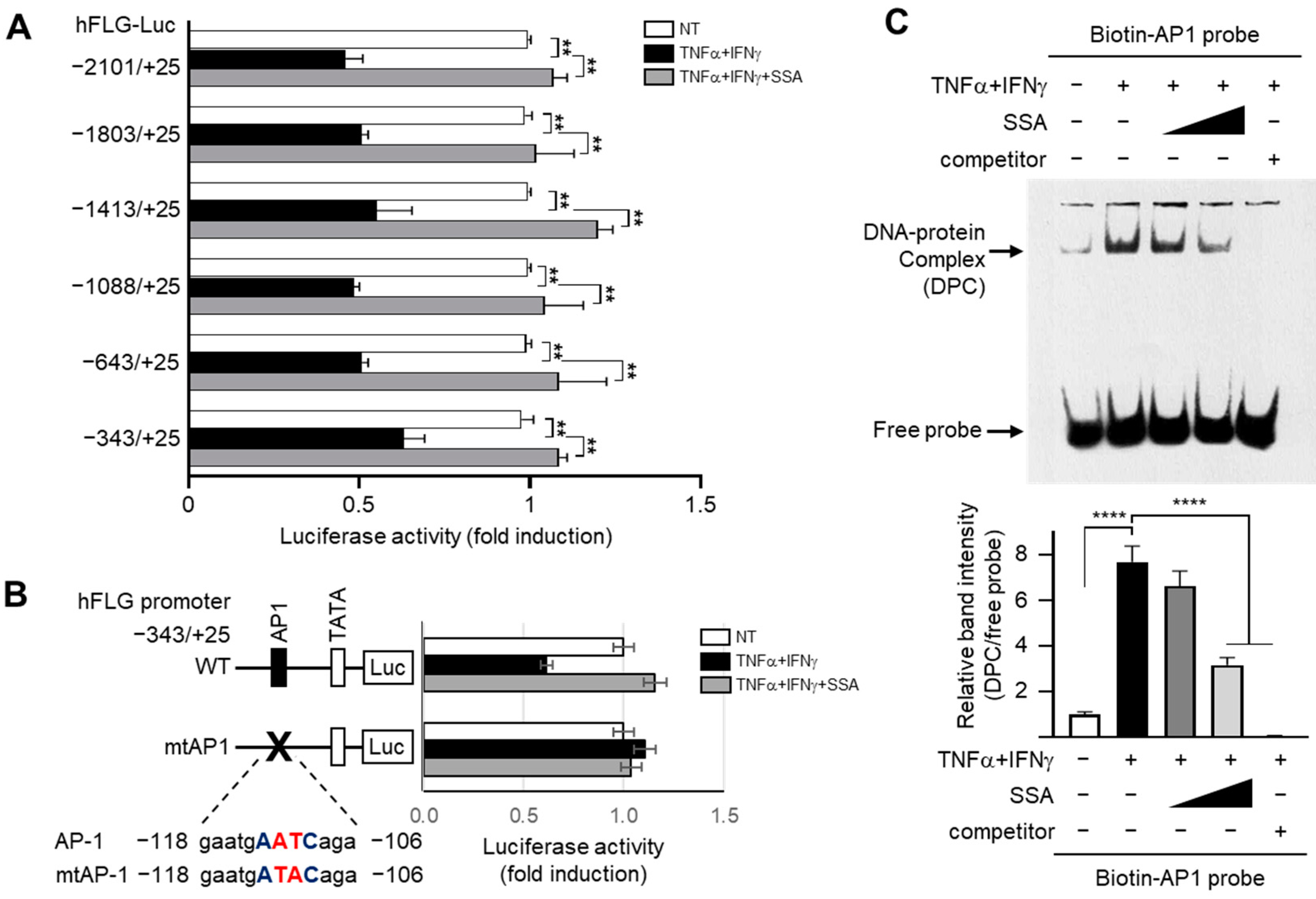
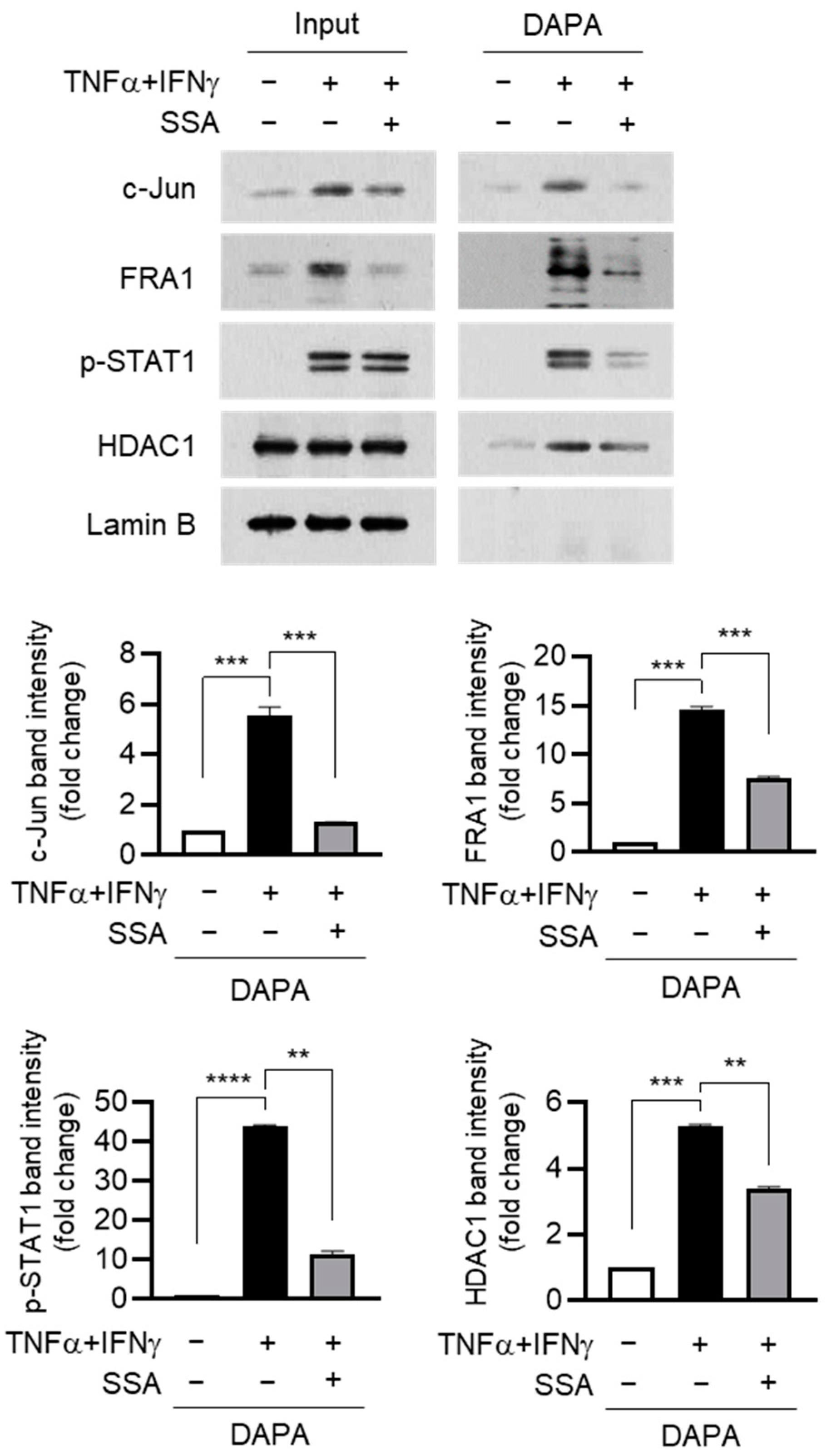
2.6. SSA Inhibits the Binding of c-Jun and FRA1 to the AP1 Motif of the FLG Promoter under TNFα + IFNγ Stimulation
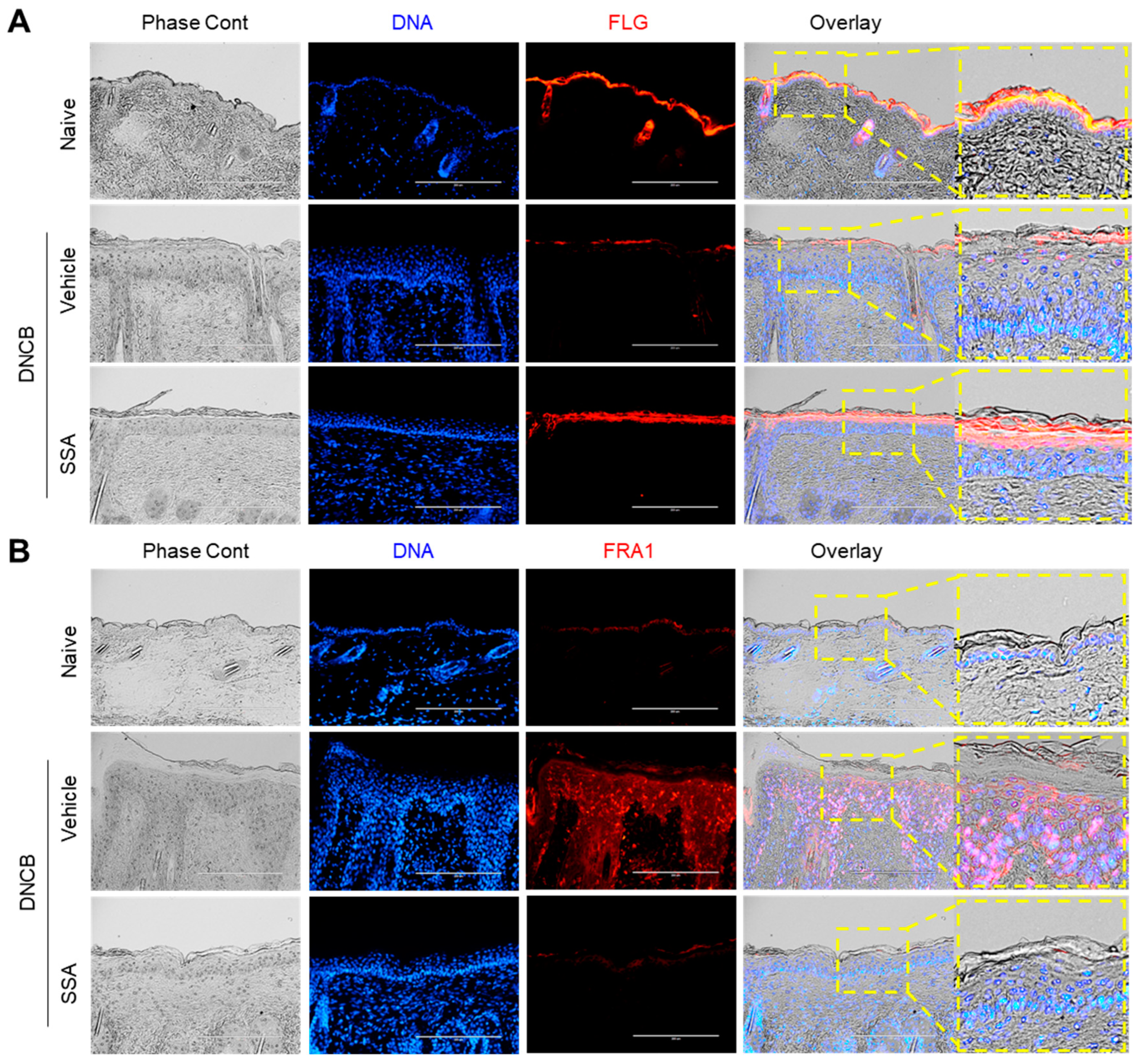
2.7. Effect of Topical Application of SSA on the Amelioration of DNCB-Induced AD-Like Skin Lesions in BALB/c Mice
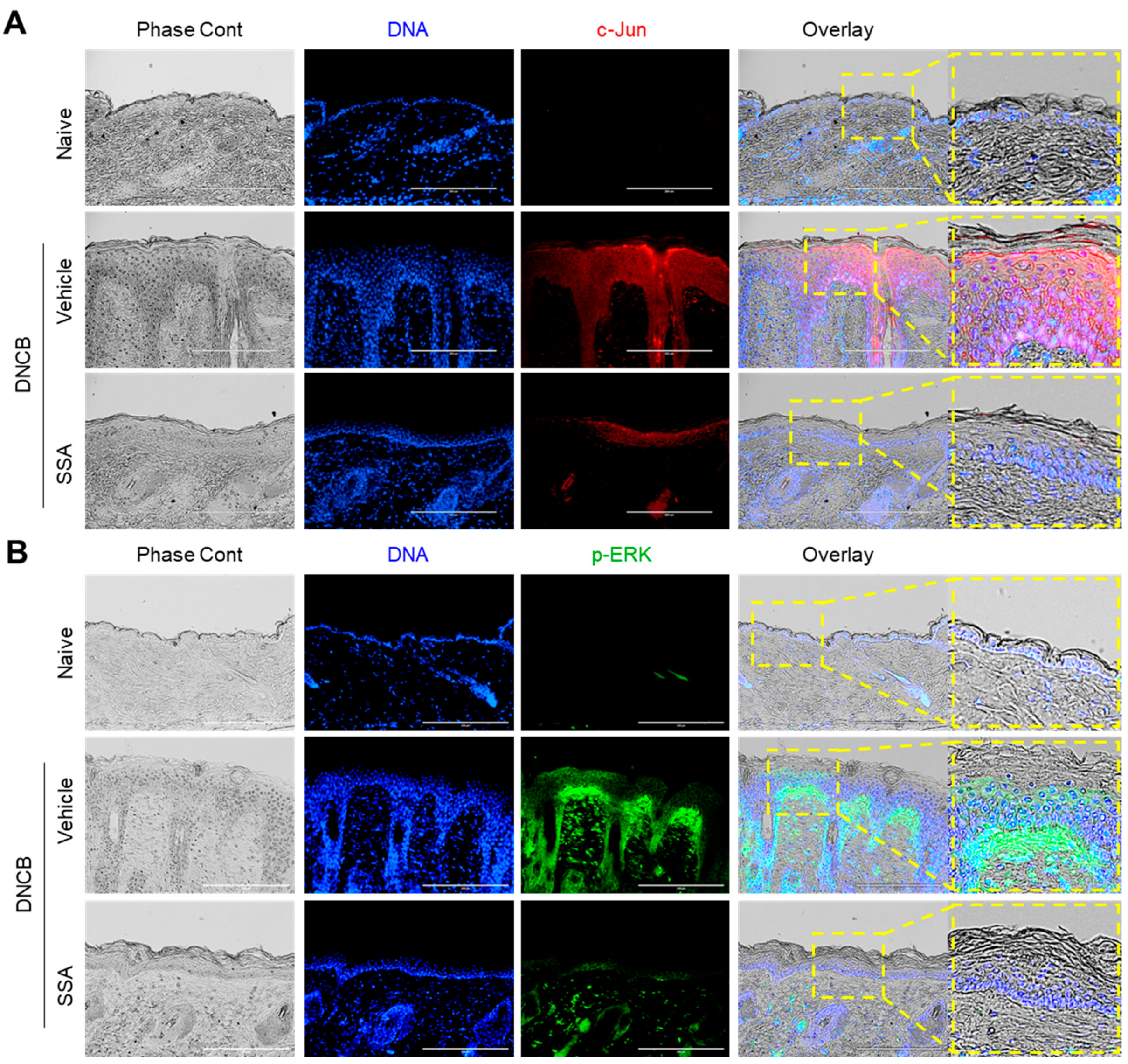
2.8. SSA Modulates the Expression of FLG, FRA1, c-Jun, and p-ERK in DNCB-Treated BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cells and Culture Conditions
4.3. Cytotoxicity Assay
4.4. Reverse Transcription Polymerase Chain Reaction and Quantitative Real-Time PCR
4.5. Immunoblot Analysis
4.6. Immunofluorescence
4.7. Construction and Internal Deletion of Human FLG Promoter–Reporter Constructs
4.8. Luciferase Promoter–Reporter Assay
4.9. Electrophoretic Mobility Shift Assay
4.10. DNA Affinity Precipitation Assay
4.11. DNCB-Challenged Atopic-Dermatitis-like Skin Lesions in BALB/c Mice
4.12. Fluorescent Immunohistochemical Staining
4.13. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eckert, R.L.; Rorke, E.A. Molecular biology of keratinocyte differentiation. Environ. Health Perspect. 1989, 80, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, G.M.; Sandilands, A.; McLean, W.I.; Irvine, A.D. Filaggrin in atopic dermatitis. J. Allergy Clin. Immunol. 2008, 122, 689–693. [Google Scholar] [CrossRef]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Chun, S.-Y.; Lee, M.-G.; Kim, S.; Jang, T.-J.; Nam, K.-S. The prevention of TNF-α/IFN-γ mixture-induced inflammation in human keratinocyte and atopic dermatitis-like skin lesions in Nc/Nga mice by mineral-balanced deep sea water. Biomed. Pharmacother. 2018, 97, 1331–1340. [Google Scholar] [CrossRef]
- Zaniboni, M.C.; Samorano, L.P.; Orfali, R.L.; Aoki, V. Skin barrier in atopic dermatitis: Beyond filaggrin. An. Bras. Dermatol. 2016, 91, 472–478. [Google Scholar] [CrossRef]
- Thyssen, J.; Godoy-Gijon, E.; Elias, P. Ichthyosis vulgaris: The filaggrin mutation disease. Br. J. Dermatol. 2013, 168, 1155–1166. [Google Scholar] [CrossRef]
- Brown, S.J.; Relton, C.L.; Liao, H.; Zhao, Y.; Sandilands, A.; Wilson, I.J.; Burn, J.; Reynolds, N.J.; McLean, W.I.; Cordell, H.J. Filaggrin null mutations and childhood atopic eczema: A population-based case-control study. J. Allergy Clin. Immunol. 2008, 121, 940–946.e3. [Google Scholar] [CrossRef]
- Gruber, R.; Elias, P.M.; Crumrine, D.; Lin, T.-K.; Brandner, J.M.; Hachem, J.-P.; Presland, R.B.; Fleckman, P.; Janecke, A.R.; Sandilands, A. Filaggrin genotype in ichthyosis vulgaris predicts abnormalities in epidermal structure and function. Am. J. Pathol. 2011, 178, 2252–2263. [Google Scholar] [CrossRef]
- Carretero, M.; Guerrero-Aspizua, S.; Illera, N.; Galvez, V.; Navarro, M.; Garcia-Garcia, F.; Dopazo, J.; Jorcano, J.L.; Larcher, F.; Del Rio, M. Differential features between chronic skin inflammatory diseases revealed in skin-humanized psoriasis and atopic dermatitis mouse models. J. Investig. Dermatol. 2016, 136, 136–145. [Google Scholar] [CrossRef]
- Danso, M.O.; van Drongelen, V.; Mulder, A.; van Esch, J.; Scott, H.; van Smeden, J.; El Ghalbzouri, A.; Bouwstra, J.A. TNF-alpha and Th2 cytokines induce atopic dermatitis-like features on epidermal differentiation proteins and stratum corneum lipids in human skin equivalents. J. Investig. Dermatol. 2014, 134, 1941–1950. [Google Scholar] [CrossRef]
- Ahn, S.S.; Yeo, H.; Jung, E.; Lim, Y.; Lee, Y.H.; Shin, S.Y. FRA1: C-JUN: HDAC1 complex down-regulates filaggrin expression upon TNFα and IFNγ stimulation in keratinocytes. Proc. Natl. Acad. Sci. USA 2022, 119, e2123451119. [Google Scholar] [CrossRef]
- Ahn, S.S.; Lee, Y.H.; Yeo, H.; Jung, E.; Lim, Y.; Shin, S.Y.; Saikosaponin, A.; Saikosaponin, C. Reduce TNF-alpha-Induced TSLP Expression through Inhibition of MAPK-Mediated EGR1 Expression in HaCaT Keratinocytes. Int. J. Mol. Sci. 2022, 23, 4857. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.S.; Lee, Y.H.; Yeo, H.; Lee, Y.; Min, D.S.; Lim, Y.; Shin, S.Y. Effect of 6, 7-dimethoxy-2, 2-dimethyl-2H-chromene (agerarin) on the recovery of filaggrin expression through targeting of Janus kinases in the inflammatory skin. J. Food Drug Anal. 2020, 28, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.L.; Wink, M. Genus Bupleurum: A review of its phytochemistry, pharmacology and modes of action. J. Pharm. Pharmacol. 2011, 63, 305–321. [Google Scholar] [CrossRef]
- Yang, F.; Dong, X.; Yin, X.; Wang, W.; You, L.; Ni, J. Radix Bupleuri: A review of traditional uses, botany, phytochemistry, pharmacology, and toxicology. BioMed Res. Int. 2017, 2017, 7597596. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Huang, N.; Liu, R.; Sun, R. A comprehensive review and perspectives on pharmacology and toxicology of saikosaponins. Phytomedicine 2018, 50, 73–87. [Google Scholar] [CrossRef]
- Ahmad, S.; Azid, N.A.; Boer, J.C.; Lim, J.; Chen, X.; Plebanski, M.; Mohamud, R. The key role of TNF-TNFR2 interactions in the modulation of allergic inflammation: A review. Front. Immunol. 2018, 9, 2572. [Google Scholar] [CrossRef]
- Talotta, F.; Casalino, L.; Verde, P. The nuclear oncoprotein Fra-1: A transcription factor knocking on therapeutic applications’ door. Oncogene 2020, 39, 4491–4506. [Google Scholar] [CrossRef]
- He, Y.-Y.; Zhou, H.-F.; Chen, L.; Wang, Y.-T.; Xie, W.-L.; Xu, Z.-Z.; Xiong, Y.; Feng, Y.-Q.; Liu, G.-Y.; Li, X. The Fra-1: Novel role in regulating extensive immune cell states and affecting inflammatory diseases. Front. Immunol. 2022, 13, 954744. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhu, G.; Li, Y.; Padia, R.N.; Dong, Z.; Pan, Z.K.; Liu, K.; Huang, S. Extracellular signal–regulated kinase signaling pathway regulates breast cancer cell migration by maintaining slug expression. Cancer Res. 2009, 69, 9228–9235. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.J.; Aziz, N.; McMahon, M. The repertoire of fos and jun proteins expressed during the G1 phase of the cell cycle is determined by the duration of mitogen-activated protein kinase activation. Mol. Cell. Biol. 1999, 19, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Casalino, L.; De Cesare, D.; Verde, P. Accumulation of Fra-1 in ras-transformed cells depends on both transcriptional autoregulation and MEK-dependent posttranslational stabilization. Mol. Cell. Biol. 2003, 23, 4401–4415. [Google Scholar] [CrossRef]
- Tulchinsky, E.; Pringle, J.H.; Caramel, J.; Ansieau, S. Plasticity of melanoma and EMT-TF reprogramming. Oncotarget 2014, 5, 1. [Google Scholar] [CrossRef]
- Yan, F.; Li, F.; Liu, J.; Ye, S.; Zhang, Y.; Jia, J.; Li, H.; Chen, D.; Mo, X. The formulae and biologically active ingredients of Chinese herbal medicines for the treatment of atopic dermatitis. Biomed. Pharmacother. 2020, 127, 110142. [Google Scholar] [CrossRef]
- Lee, H.; Shin, J.J.; Bae, H.C.; Ryu, W.-I.; Son, S.W. Toluene downregulates filaggrin expression via the extracellular signal-regulated kinase and signal transducer and activator of transcription–dependent pathways. J. Allergy Clin. Immunol. 2017, 139, 355–358.e5. [Google Scholar] [CrossRef]
- Song, Y.; Qian, L.; Song, S.; Chen, L.; Zhang, Y.; Yuan, G.; Zhang, H.; Xia, Q.; Hu, M.; Yu, M. Fra-1 and Stat3 synergistically regulate activation of human MMP-9 gene. Mol. Immunol. 2008, 45, 137–143. [Google Scholar] [CrossRef]
- Ray, S.; Lee, C.; Hou, T.; Boldogh, I.; Brasier, A.R. Requirement of histone deacetylase1 (HDAC1) in signal transducer and activator of transcription 3 (STAT3) nucleocytoplasmic distribution. Nucleic Acids Res. 2008, 36, 4510–4520. [Google Scholar] [CrossRef]
- Nusinzon, I.; Horvath, C.M. Interferon-stimulated transcription and innate antiviral immunity require deacetylase activity and histone deacetylase 1. Proc. Natl. Acad. Sci. USA 2003, 100, 14742–14747. [Google Scholar] [CrossRef]
- Brunmeir, R.; Lagger, S.; Seiser, C. Histone deacetylase 1 and 2-controlled embryonic development and cell differentiation. Int. J. Dev. Biol. 2009, 53, 275–289. [Google Scholar] [CrossRef]
- Hald, A.; Andrés, R.; Salskov-Iversen, M.; Kjellerup, R.; Iversen, L.; Johansen, C. STAT1 expression and activation is increased in lesional psoriatic skin. Br. J. Dermatol. 2013, 168, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lai, Y. Pharmacological properties and derivatives of saikosaponins—A review of recent studies. J. Pharm. Pharmacol. 2023, 75, 898–909. [Google Scholar] [CrossRef]
- Sellers, R.S. Translating mouse models: Immune variation and efficacy testing. Toxicol. Pathol. 2017, 45, 134–145. [Google Scholar] [CrossRef]
- Wen, H.; Jung, H.; Li, X. Drug delivery approaches in addressing clinical pharmacology-related issues: Opportunities and challenges. AAPS J. 2015, 17, 1327–1340. [Google Scholar] [CrossRef]
- Moniz, T.; Costa Lima, S.A.; Reis, S. Human skin models: From healthy to disease-mimetic systems; characteristics and applications. Br. J. Pharmacol. 2020, 177, 4314–4329. [Google Scholar] [CrossRef]
- Gallinari, P.; Marco, S.D.; Jones, P.; Pallaoro, M.; Steinkühler, C. HDACs, histone deacetylation and gene transcription: From molecular biology to cancer therapeutics. Cell Res. 2007, 17, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; O’Neil, K.; Capell, B.C. Histone modifiers: Dynamic regulators of the cutaneous transcriptome. J. Dermatol. Sci. 2018, 89, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Jung Jung, Y.; Koh, D.; Lim, Y.; Lee, Y.H.; Shin, S.Y. A synthetic chalcone, 2′-hydroxy-2,3,5′-trimethoxychalcone triggers unfolded protein response-mediated apoptosis in breast cancer cells. Cancer Lett. 2016, 372, 1–9. [Google Scholar] [CrossRef]
- Yeo, H.; Ahn, S.S.; Lee, J.Y.; Jung, E.; Jeong, M.; Kang, G.S.; Ahn, S.; Lee, Y.; Koh, D.; Lee, Y.H. Disrupting the DNA binding of EGR-1 with a small-molecule inhibitor ameliorates 2, 4-dinitrochlorobenzene-induced skin inflammation. J. Investig. Dermatol. 2021, 141, 1851–1855. [Google Scholar] [CrossRef]
- Jung, E.; Ou, S.; Ahn, S.S.; Yeo, H.; Lee, Y.H.; Shin, S.Y. The JNK-EGR1 signaling axis promotes TNF-alpha-induced endothelial differentiation of human mesenchymal stem cells via VEGFR2 expression. Cell Death Differ. 2023, 30, 356–368. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, S.S.; Yeo, H.; Jung, E.; Kim, T.Y.; Han, J.; Lee, Y.H.; Shin, S.Y. Saikosaponin A Recovers Impaired Filaggrin Levels in Inflamed Skin by Downregulating the Expression of FRA1 and c-Jun. Molecules 2024, 29, 4064. https://doi.org/10.3390/molecules29174064
Ahn SS, Yeo H, Jung E, Kim TY, Han J, Lee YH, Shin SY. Saikosaponin A Recovers Impaired Filaggrin Levels in Inflamed Skin by Downregulating the Expression of FRA1 and c-Jun. Molecules. 2024; 29(17):4064. https://doi.org/10.3390/molecules29174064
Chicago/Turabian StyleAhn, Sung Shin, Hyunjin Yeo, Euitaek Jung, Tae Yoon Kim, Junekyu Han, Young Han Lee, and Soon Young Shin. 2024. "Saikosaponin A Recovers Impaired Filaggrin Levels in Inflamed Skin by Downregulating the Expression of FRA1 and c-Jun" Molecules 29, no. 17: 4064. https://doi.org/10.3390/molecules29174064
APA StyleAhn, S. S., Yeo, H., Jung, E., Kim, T. Y., Han, J., Lee, Y. H., & Shin, S. Y. (2024). Saikosaponin A Recovers Impaired Filaggrin Levels in Inflamed Skin by Downregulating the Expression of FRA1 and c-Jun. Molecules, 29(17), 4064. https://doi.org/10.3390/molecules29174064