
Fucoxanthin Induces Ferroptosis in Cancer Cells via Downregulation of the Nrf2/HO−1/GPX4 Pathway

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Inhibitory Effects of Fucoxanthin on SCC−25 Tongue Cancer Cells
2.2. Effects of Fucoxanthin on ROS in SCC−25 Cells
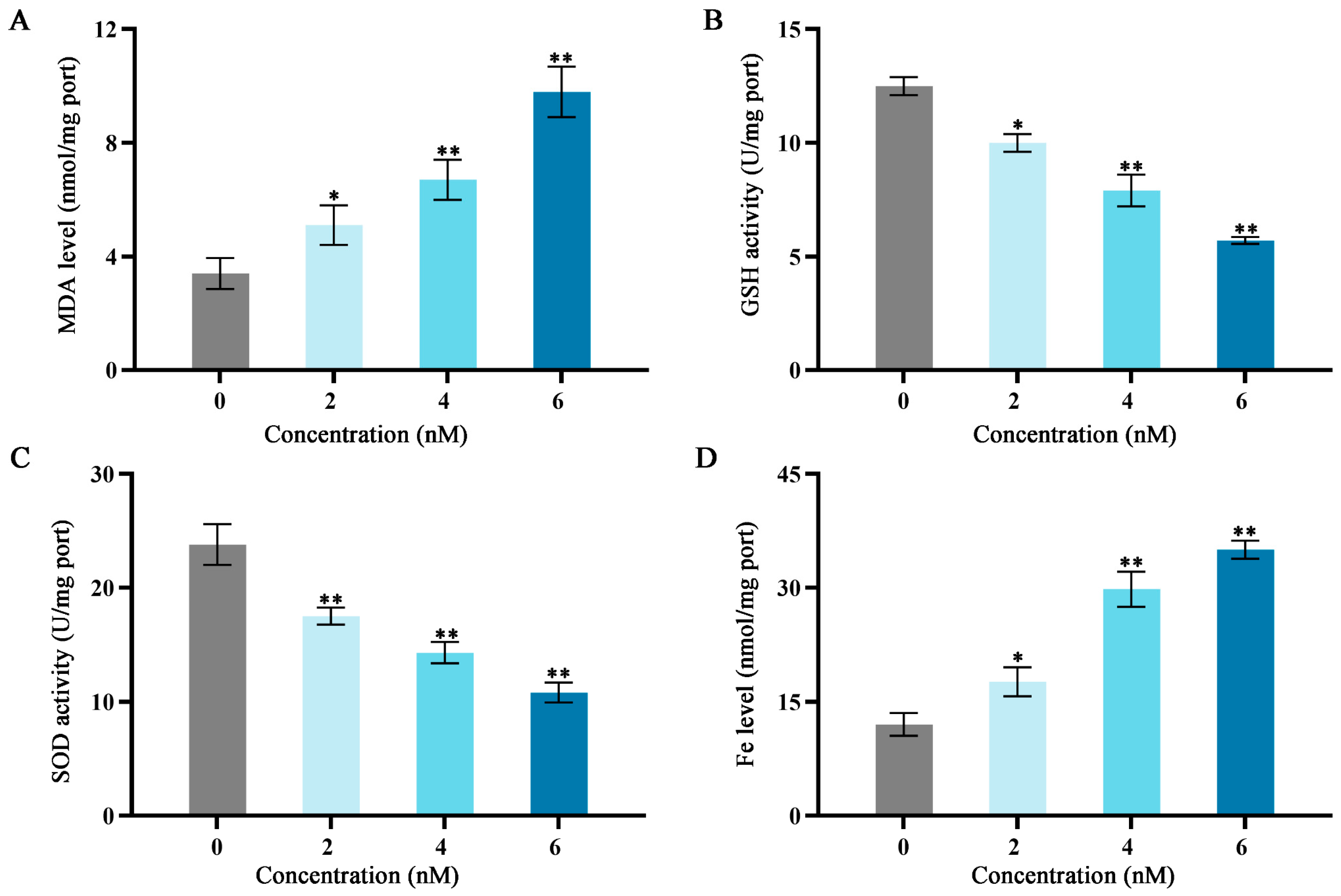
2.3. Effects of Fucoxanthin on Malondialdehyde (MDA), Superoxide Dismutase (SOD), Glutathione (GSH), and Iron (Fe) in SCC−25 Cells
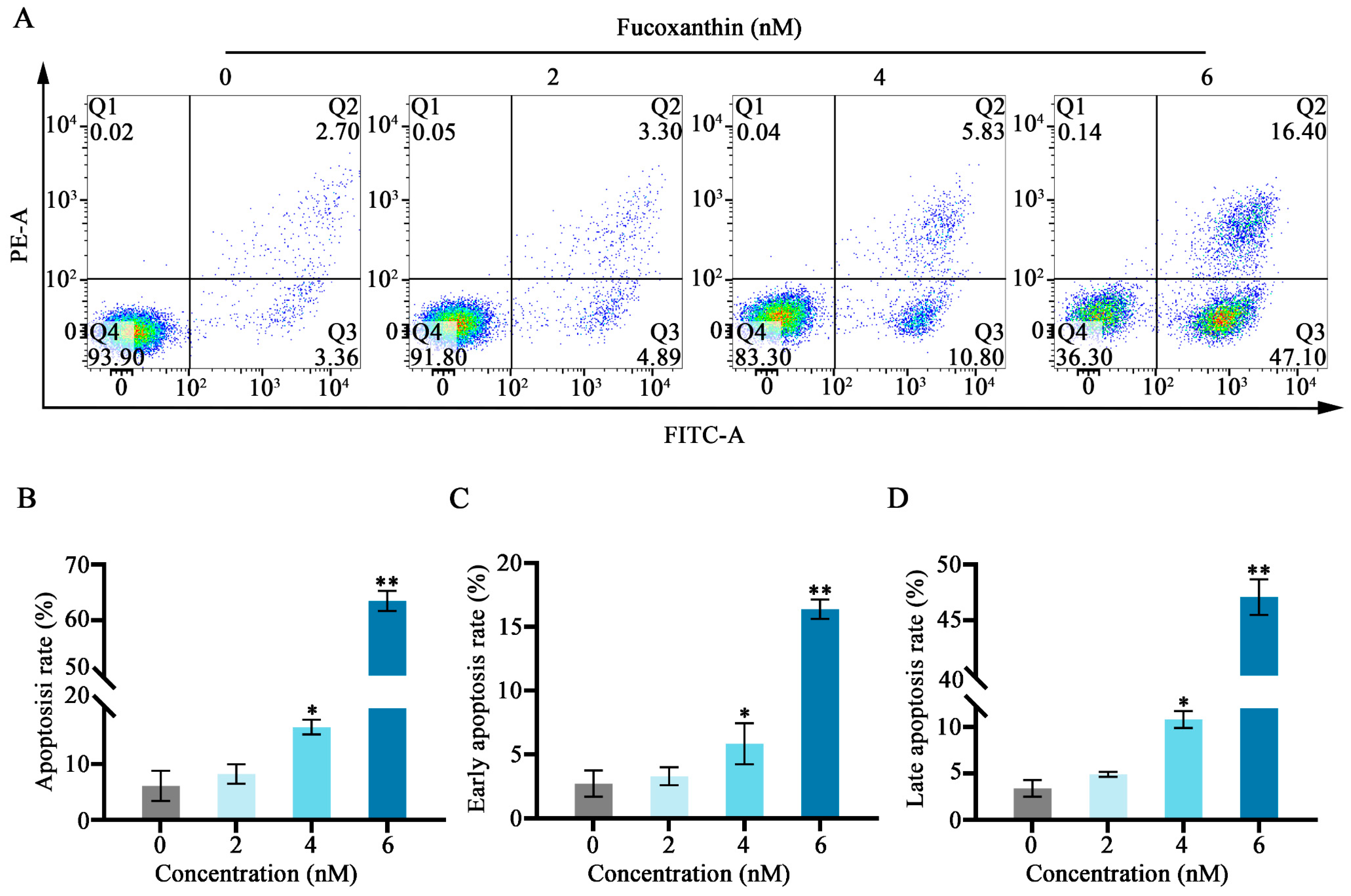
2.4. Effects of Fucoxanthin on Apoptosis in SCC−25 Cells
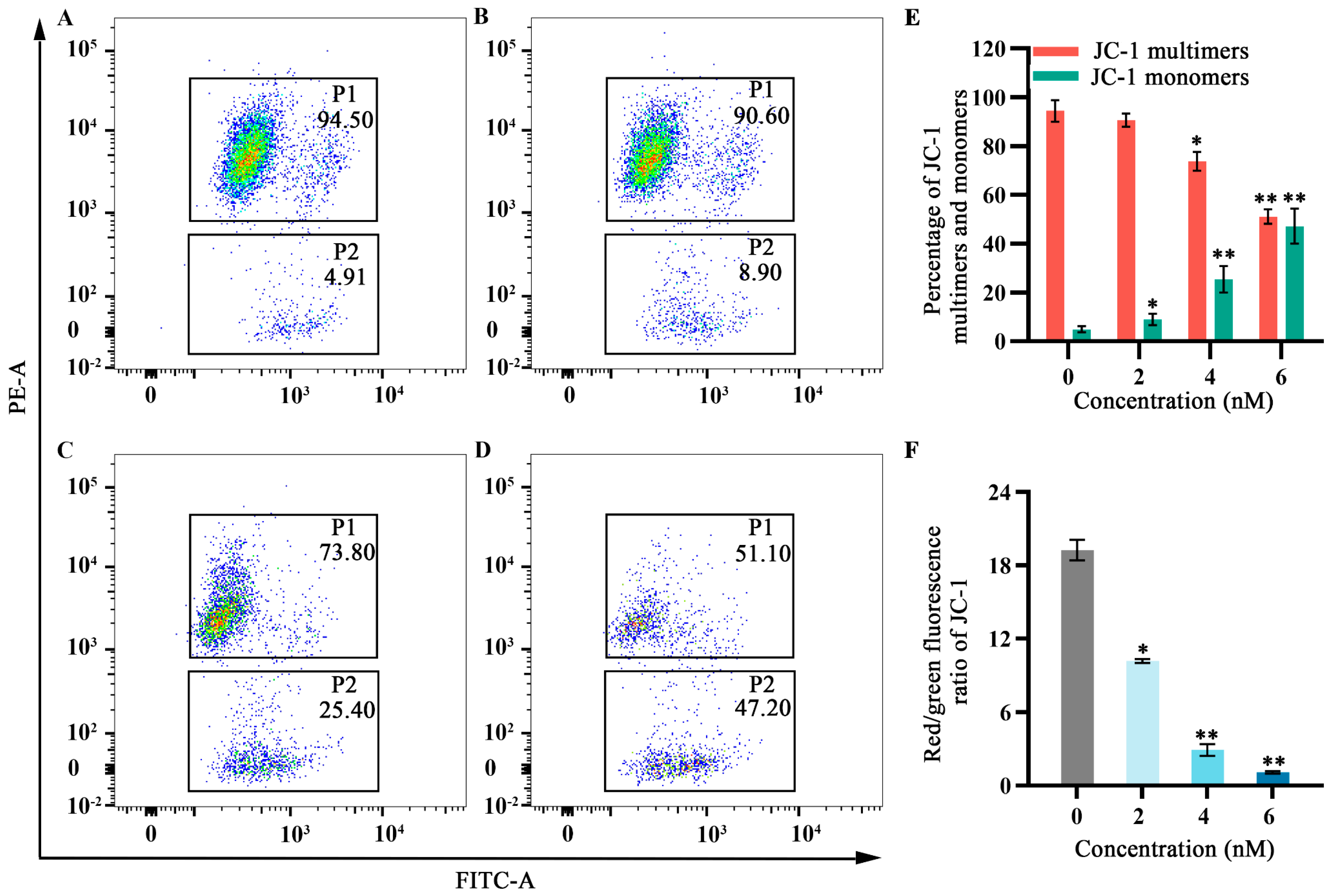
2.5. Effects of Fucoxanthin on Mitochondrial Membrane Potential in SCC−25 Tongue Cancer Cells
2.6. Effects of Fucoxanthin on mRNA Expression Levels of p53, SLC7A11, GPX4, Nuclear Factor Erythroid 2−Related Factor 2 (Nrf2), Keap1, Heme Oxygenase 1 (HO−1), and TF Receptor 1 (TFR1) in SCC−25 Tongue Cancer Cells
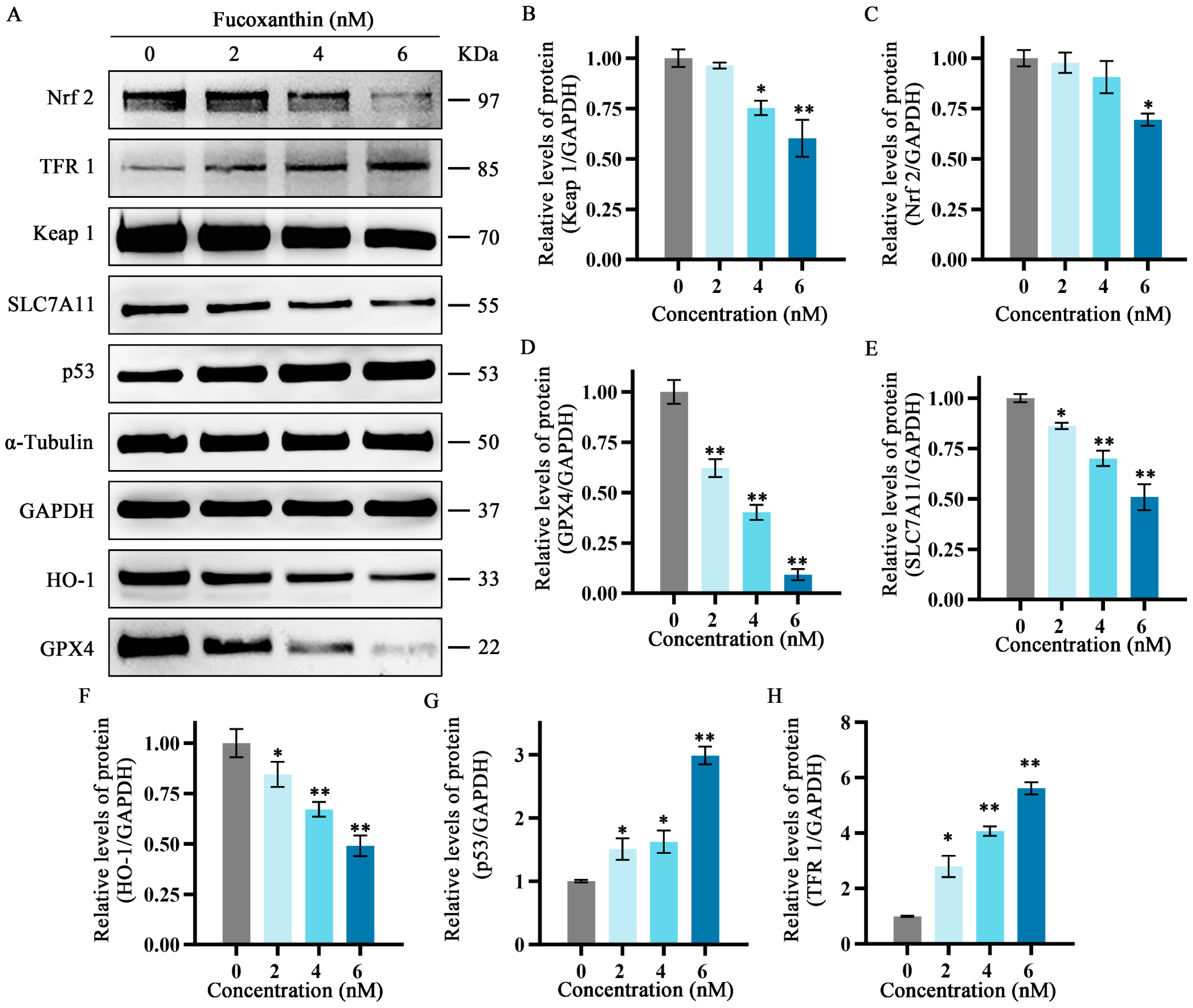
2.7. Effects of Fucoxanthin on Protein Expression Levels of p53, SLC7A11, GPX4, Nrf2, Keap1, HO−1, and TFR1 in SCC−25 Tongue Cancer Cells
2.8. Molecular Docking Validation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. MTT Assay to Detect Cell Viability
4.3. ROS Detection by Flow Cytometry
4.4. Measurement of MDA, GSH, SOD, and Fe Levels
4.5. Apoptosis Detection by Flow Cytometry
4.6. Mitochondrial Membrane Potential Measurement by Flow Cytometry
4.7. RT−qPCR Analysis of Ferroptosis−Related Gene Expression
4.8. Western Blotting Analysis of Ferroptosis−Related Protein Expression
4.9. Molecular Docking
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, T.; Yang, L.L.; Li, Z.; Sun, M.; Lv, N. F25, a novel synthetic quinoline derivative, inhibits tongue cancer cell invasion and survival by the PPAR pathway in vitro and vivo. Chem. Biol. Interact. 2024, 391, 110891. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, F.; Koizumi, K.; Ito, N.; Higaki, M.; Ishida, Y.; Hamada, A.; Yamasaki, S.; Tani, R.; Yanamoto, S. A study of the prognostic factors for late cervical lymph node metastasis and distant metastasis in patients with cT1−2N0 tongue cancer. J. Clin. Med. 2024, 13, 976. [Google Scholar] [CrossRef] [PubMed]
- da Silva, L.A.B.; da Costa, L.M.; Massetti, A.C.P.; de Lucena Pereira, L.; da Silveira, E.J.D.; Salo, T.A.; Coletta, R.D.; da Costa Miguel, M.C. Silencing of heat shock factor 1 (HSF1) inhibits proliferation, invasion, and epithelial−mesenchymal transition in oral squamous cell carcinoma. J. Oral Pathol. Med. 2023, 52, 961–970. [Google Scholar] [CrossRef]
- Wu, C.Y.; Xiang, S.Y.; Wang, H.T.; Zhang, X.M.; Tian, X.Y.; Tan, M.Q.; Su, W.T. Orally deliverable sequence−targeted fucoxanthin−loaded biomimetic extracellular vesicles for alleviation of nonalcoholic fatty liver disease. ACS Appl. Mater. Interfaces 2024, 16, 9854–9867. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Kim, S.C.; Oh, Y. Fucoxanthin abrogates ionizing radiation−induced inflammatory responses by modulating sirtuin 1 in macrophages. Mar. Drugs 2023, 21, 635. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kaur, A.; Singh, S. Fucoxanthin mitigates valproic acid−induced autistic behavior through modulation of the AKT/GSK−3beta signaling pathway. Eur. J. Pharmacol. 2024, 967, 176335. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Mendonca, P.; Messeha, S.S.; Soliman, K.F.A. Anticancer effects of fucoxanthin through cell cycle arrest, apoptosis induction, and angiogenesis inhibition in triple−negative breast cancer cells. Molecules 2023, 28, 6536. [Google Scholar] [CrossRef]
- Terasaki, M.; Tsuruoka, K.; Tanaka, T.; Maeda, H.; Shibata, M.; Miyashita, K.; Kanemitsu, Y.; Sekine, S.; Takahashi, M.; Yagishita, S.; et al. Fucoxanthin inhibits development of sigmoid colorectal cancer in a PDX model with alterations of growth, adhesion, and cell cycle signals. Cancer Genom. Proteom. 2023, 20, 686–705. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Liu, T.Y.; Zhang, L.T.; Hua, Z.H.; Jin, X.A.; Xu, F.; Ji, J.C.; Xu, B.H.; Ding, H.M. Effects and mechanisms of fucoxanthin from Hizikia fusiforme on inhibiting tongue squamous cell carcinoma proliferation via AKT/mTOR−mediated glycolysis. J. Food Biochem. 2023, 2023, 7944733. [Google Scholar] [CrossRef]
- Suwanmanee, G.; Tantrawatpan, C.; Kheolamai, P.; Paraoan, L.; Manochantr, S. Fucoxanthin diminishes oxidative stress damage in human placenta−derived mesenchymal stem cells through the PI3K/Akt/Nrf−2 pathway. Sci. Rep. 2023, 13, 22974. [Google Scholar] [CrossRef]
- Koshak, M.F.; El−Readi, M.Z.; Elzubier, M.E.; Refaat, B.; Almaimani, R.A.; Idris, S.; Althubiti, M.; Al−Amodi, H.S.; Eid, S.Y. Antioxidative and anti−inflammatory protective effects of fucoxanthin against paracetamol−induced hepatotoxicity in rats. Mar. Drugs 2023, 21, 592. [Google Scholar] [CrossRef]
- Chen, Y.; Dong, J.H.; Gong, L.Y.; Hong, Y.R.; Hu, C.W.; Bao, Y.J.; Chen, H.Y.; Liu, L.; Huang, L.; Zhao, Y.Z.; et al. Fucoxanthin, a marine derived carotenoid, attenuates surgery−induced cognitive impairments via activating Akt and ERK pathways in aged mice. Phytomedicine 2023, 120, 155043. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron−dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Lu, X.J.; Liang, Y.; Yang, L.; Yin, Y.H.; Chen, H. Naringenin alleviates liver fibrosis by triggering autophagy−dependent ferroptosis in hepatic stellate cells. Heliyon 2024, 10, e28865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.Z.; Chang, L.; Ren, Y.Q.; Sui, M.S.; Fu, Y.T.; Hao, L.R. Quercetin improves diabetic kidney disease by inhibiting ferroptosis and regulating the Nrf2 in streptozotocin−induced diabetic rats. Ren. Fail. 2024, 46, 2327495. [Google Scholar] [CrossRef]
- Wei, W.; Tian, L.Q.; Zheng, X.Y.; Zhong, L.; Chen, Y.; Dong, H.; Zhang, G.B.; Wang, S.B.; Tong, X.M. Expression of GPX4 by oncolytic vaccinia virus can significantly enhance CD8+T cell function and its impact against pancreatic ductal adenocarcinoma. Oncoimmunology 2024, 13, 2322173. [Google Scholar] [CrossRef]
- Chen, P.; Ye, Q.X.; Liang, S.; Zeng, L.H. Cephaeline promotes ferroptosis by targeting NRF2 to exert anti−lung cancer efficacy. Pharm. Biol. 2024, 62, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Nath, P.; Alfarsi, L.H.; El−Ansari, R.; Masisi, B.K.; Erkan, B.; Fakroun, A.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The amino acid transporter SLC7A11 expression in breast cancer. Cancer Biol. Ther. 2024, 25, 2291855. [Google Scholar] [CrossRef] [PubMed]
- Struckmeier, A.K.; Buchbender, M.; Agaimy, A.; Kesting, M. Prevalence and implications of bilateral and solely contralateral lymph node metastases in oral squamous cell carcinoma. Clin. Oral Investig. 2024, 28, 267. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, X.Y.; Zeng, Y.C.; Mo, X.L.; Hong, S.Q.; He, H.; Li, J.; Fatima, S.; Liu, Q.H. Oxidative stress induces mitochondrial iron overload and ferroptotic cell death. Sci. Rep. 2023, 13, 15515. [Google Scholar] [CrossRef]
- Chen, L.C.; Lo, Y.S.; Ho, H.Y.; Lin, C.C.; Chuang, Y.C.; Chang, W.C.; Hsieh, M.J. LDL receptor−related protein 1b polymorphisms associated with increased risk of lymph node metastasis in oral cancer group with diabetes mellitus. Int. J. Mol. Sci. 2024, 25, 3963. [Google Scholar] [CrossRef] [PubMed]
- Yap, H.Y.Y.; Kong, B.H.; Yap, C.S.A.; Ong, K.C.; Zain, R.B.; Tan, S.H.; Zaini, Z.M.; Ng, S.T.; Tan, C.S.; Fung, S.Y. Immunomodulatory effect and an intervention of TNF signalling leading to apoptotic and cell cycle arrest on orl−204 oral cancer cells by tiger milk mushroom, Lignosus rhinocerus. Food Technol. Biotechnol. 2022, 60, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Li, J.; Xiong, R.G.; Wu, S.X.; Xu, X.Y.; Tang, G.Y.; Huang, S.Y.; Zhou, D.D.; Li, H.B.; Feng, Y.; et al. Effects and mechanisms of anti−diabetic dietary natural products: An updated review. Food Funct. 2024, 15, 1758–1778. [Google Scholar] [CrossRef] [PubMed]
- Din, N.A.S.; Mohd Alayudin, S.; Sofian−Seng, N.S.; Rahman, H.A.; Mohd Razali, N.S.; Lim, S.J.; Wan Mustapha, W.A. Brown algae as functional food source of fucoxanthin: A review. Foods 2022, 11, 2235. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.T.; Jian, W.Y.; Zhang, Y.Y.; Li, Q.; Zhao, Y.; Liu, R.; Zeng, Y.; Zhang, F.W.; Duan, J.G. Puerarin attenuates iron overload−induced ferroptosis in retina through a Nrf2−mediated mechanism. Mol. Nutr. Food Res. 2024, 68, e2300123. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Song, S.J.; Fan, Z.W.; Li, W.T.; Jin, X.; Jiang, W.; Bai, J.; Shi, Z.Z. PKCiota inhibits the ferroptosis of esophageal cancer cells via suppressing USP14−mediated autophagic degradation of GPX4. Antioxidants 2024, 13, 114. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Chung, S.W. ROS−mediated autophagy increases intracellular iron levels and ferroptosis by ferritin and transferrin receptor regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef]
- Wang, C.Y.; Jiang, H.B.; Liu, H.H.; Chen, S.S.; Guo, H.Y.; Ma, S.S.; Han, W.W.; Li, Y.; Wang, D. Isoforsythiaside confers neuroprotection against Alzheimer’s disease by attenuating ferroptosis and neuroinflammation in vivo and in vitro. Food Sci. Hum. Well. 2023, 12, 1730–1742. [Google Scholar] [CrossRef]
- Cui, X.H.; Gong, Y.T.; Ge, J.; Feng, X.F.; Xiong, X.M.; Shi, Z.; Zheng, Q.S.; Li, D.F.; Bi, S.X. α−Solanine induces ferroptosis in nasopharyngeal carcinoma via targeting HSP90α/p53 axis. J. Funct. Foods 2023, 104, 105517. [Google Scholar] [CrossRef]
- Wang, G.Y.; Qin, S.L.; Zheng, Y.N.; Xia, C.; Zhang, P.; Zhang, L.X.; Yao, J.H.; Yi, Y.L.; Deng, L. T−2 toxin induces ferroptosis by increasing lipid reactive oxygen species (ROS) and downregulating solute carrier family 7 member 11 (SLC7A11). J. Agric. Food Chem. 2021, 69, 15716–15727. [Google Scholar] [CrossRef]
- Fu, F.; Lai, Q.; Hu, J.G.; Zhang, L.; Zhu, X.Z.; Kou, J.P.; Yu, B.Y.; Li, F. Ruscogenin alleviates myocardial ischemia−induced ferroptosis through the activation of BCAT1/BCAT2. Antioxidants 2022, 11, 583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, W.; Feng, X.; Yang, F.; Qin, H.; Wu, S.; Hou, D.X.; Chen, J. Nrf2−are signaling acts as master pathway for the cellular antioxidant activity of fisetin. Molecules 2019, 24, 708. [Google Scholar] [CrossRef]
- Chiang, S.K.; Chen, S.E.; Chang, L.C. A dual role of heme oxygenase−1 in cancer cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef]
- Adedoyin, O.; Boddu, R.; Traylor, A.; Lever, J.M.; Bolisetty, S.; George, J.F.; Agarwal, A. Heme oxygenase−1 mitigates ferroptosis in renal proximal tubule cells. Am. J. Physiol. Renal. Physiol. 2018, 314, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Chipurupalli, S.; Jiang, P.; Liu, X.; Santos, J.L.; Marcato, P.; Rosen, K.V. Three−dimensional growth sensitizes breast cancer cells to treatment with ferroptosis−promoting drugs. Cell Death Dis. 2023, 14, 580. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhu, Q.; Qi, X.; Yang, L.R.; Rong, Y.X.; Wei, Q.; Wu, S.Q.; Lu, Q.W.; Li, L.; Jiang, M.D.; et al. Dual role of Nrf2/HO−1 pathway in Z−ligustilide−induced ferroptosis against AML cells. Phytomedicine 2024, 124, 155288. [Google Scholar] [CrossRef]
- Chen, J.; Guo, P.; Han, M.M.; Chen, K.M.; Qin, J.; Yang, F.R. Cognitive protection of sinomenine in type 2 diabetes mellitus through regulating the EGF/Nrf2/HO−1 signaling, the microbiota−gut−brain axis, and hippocampal neuron ferroptosis. Phytother. Res. 2023, 37, 3323–3341. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense Primer (5′-3′) | Antisense Primer (3′-5′) |
|---|---|---|
| p53 | GCGTGTGGAGTATTTGGATGAC | AGTGTGATGATGGTGAGGATGG |
| TFR1 | AACTCAGCAAAGTCTGGCGT | GACCCCCAATACACCGCATA |
| SLC7A11 | TCCTGCTTTGGCTCCAT | ACAGGCGTTCGTGTGAGGAGA |
| GPX4 | ACAAGAACGGCTGCGTGGTGAA | AGATCGAGGTGTTCACACACCG |
| Nrf2 | TACTCCCAGGTTGCCCACA | AAGGGCAAACATCTAC |
| Keap1 | GTGTCCATTGAGGGTATCCACC | GCTCAGCGAAGTTGGCGAT |
| HO-1 | GGCCTCCCTGTACCACATCT | GGATGTGTGGTCGGTACGTC |
| β-actin | CCTGGCACCCAGCACAAT | GGGCCGGACTCGTCATAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.-F.; Wu, J.-W.; Zhu, Y.-S.; Hua, Z.-H.; Jin, S.-Z.; Ji, J.-C.; Wang, C.-S.; Qian, G.-Y.; Jin, X.-D.; Ding, H.-M. Fucoxanthin Induces Ferroptosis in Cancer Cells via Downregulation of the Nrf2/HO−1/GPX4 Pathway. Molecules 2024, 29, 2832. https://doi.org/10.3390/molecules29122832
Du H-F, Wu J-W, Zhu Y-S, Hua Z-H, Jin S-Z, Ji J-C, Wang C-S, Qian G-Y, Jin X-D, Ding H-M. Fucoxanthin Induces Ferroptosis in Cancer Cells via Downregulation of the Nrf2/HO−1/GPX4 Pathway. Molecules. 2024; 29(12):2832. https://doi.org/10.3390/molecules29122832
Chicago/Turabian StyleDu, Hao-Fei, Jia-Wei Wu, Yu-Shan Zhu, Zheng-Hao Hua, Si-Zhou Jin, Jin-Chao Ji, Cai-Sheng Wang, Guo-Ying Qian, Xu-Dong Jin, and Hao-Miao Ding. 2024. "Fucoxanthin Induces Ferroptosis in Cancer Cells via Downregulation of the Nrf2/HO−1/GPX4 Pathway" Molecules 29, no. 12: 2832. https://doi.org/10.3390/molecules29122832
APA StyleDu, H.-F., Wu, J.-W., Zhu, Y.-S., Hua, Z.-H., Jin, S.-Z., Ji, J.-C., Wang, C.-S., Qian, G.-Y., Jin, X.-D., & Ding, H.-M. (2024). Fucoxanthin Induces Ferroptosis in Cancer Cells via Downregulation of the Nrf2/HO−1/GPX4 Pathway. Molecules, 29(12), 2832. https://doi.org/10.3390/molecules29122832