
Chemical Composition, Repellent, and Oviposition Deterrent Potential of Wild Plant Essential Oils against Three Mosquito Species

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Yield (%) of EOs
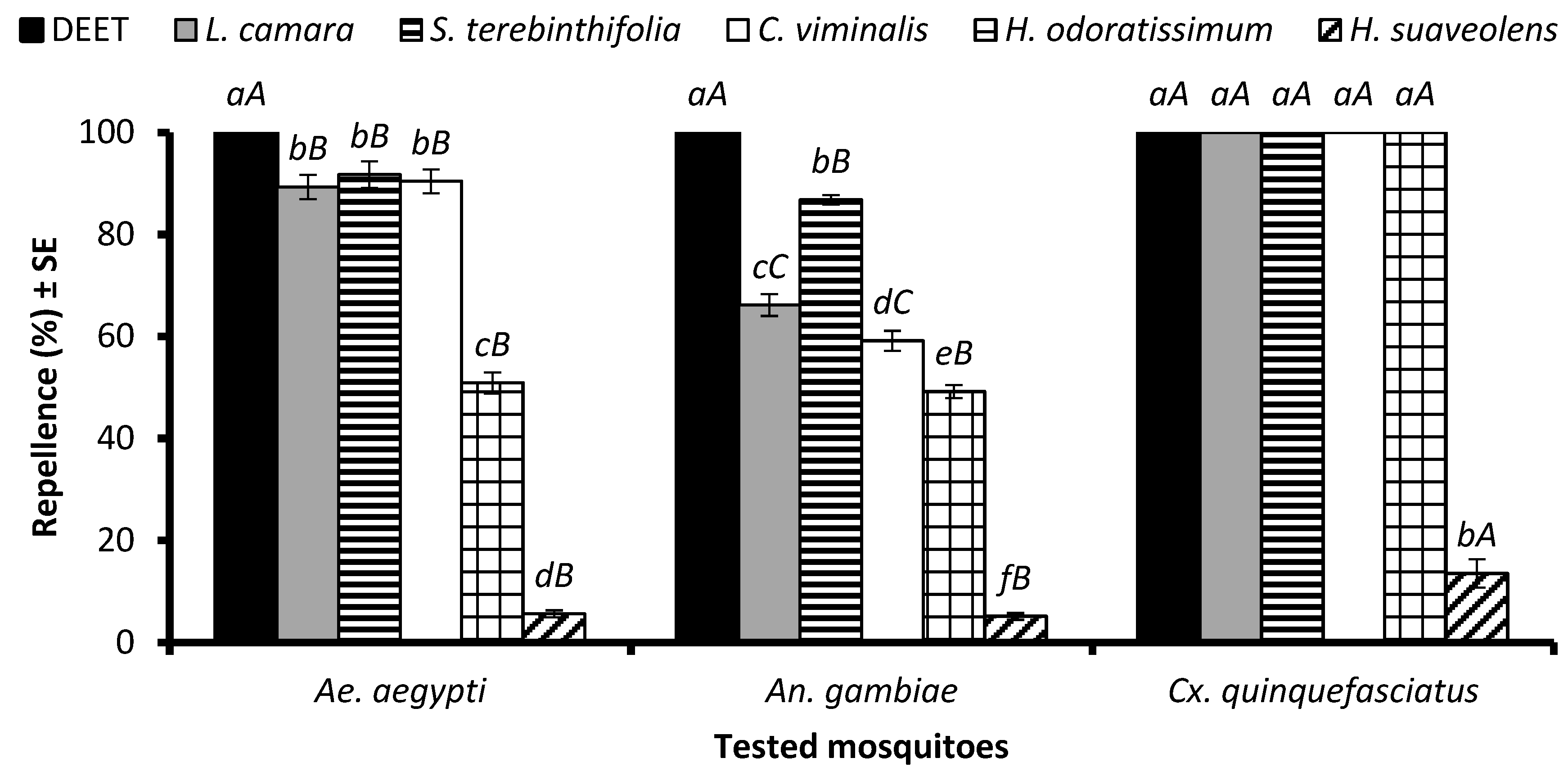
2.2. Repellency Results
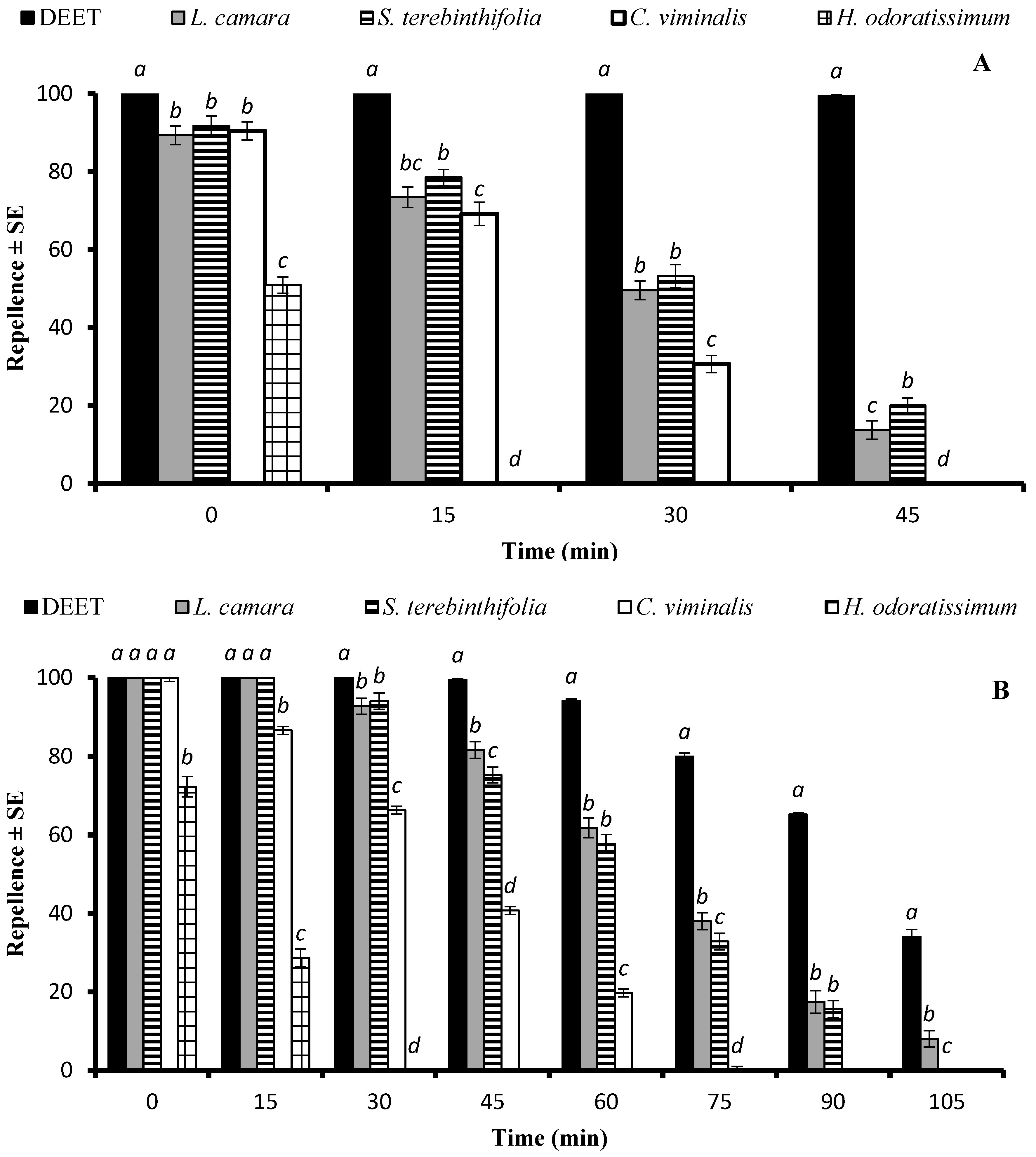
2.2.1. Time-Span Repellency against Ae. aegypti Mosquito Females
2.2.2. Time-Span Repellency against An. gambiae s. l. Mosquito Females
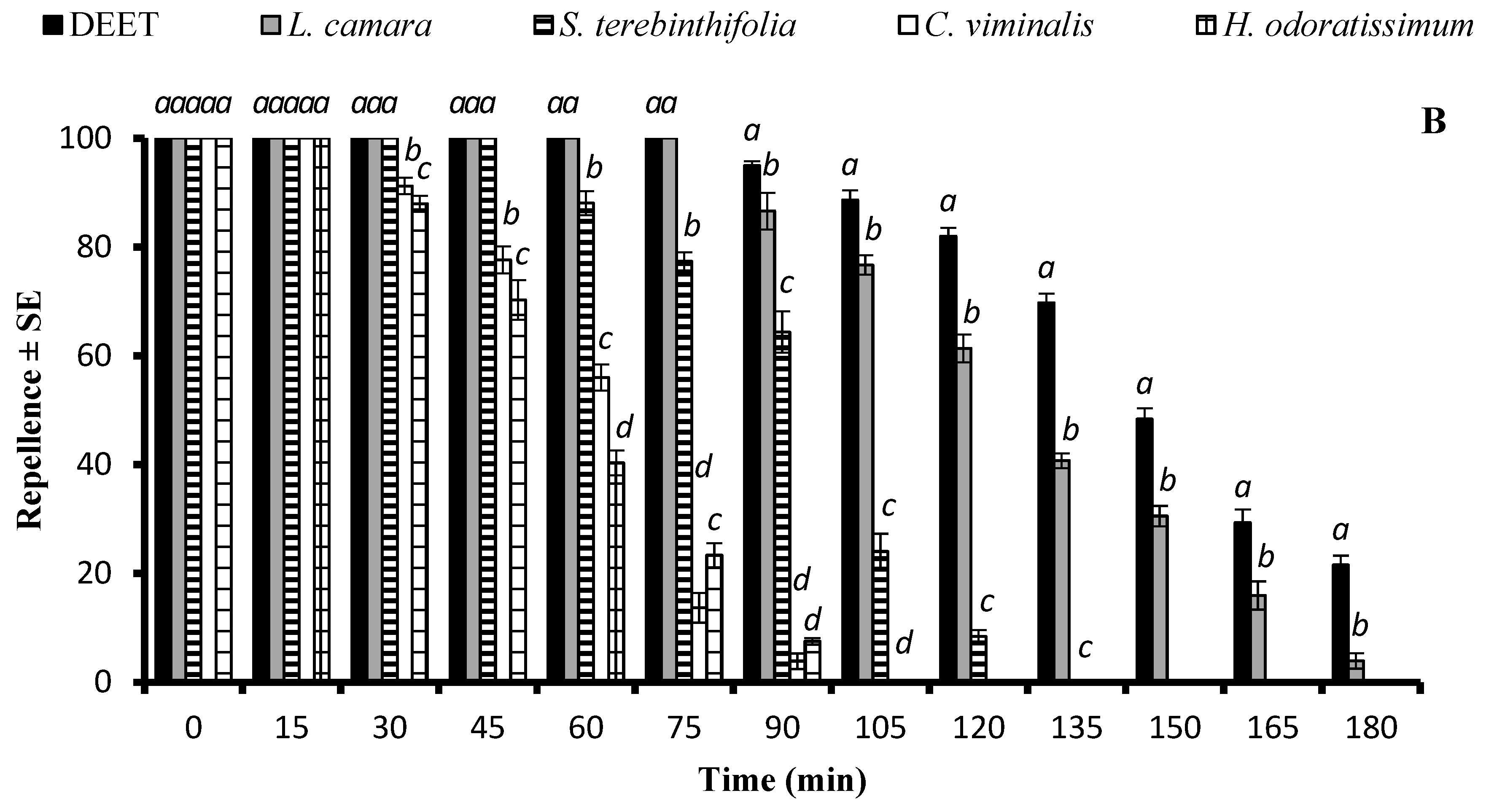
2.2.3. Time-Span Repellency against Cx. quinquefasciatus Mosquito Females
2.3. Oviposition Deterrence of EOs
2.4. Chemical Profile of EOs
3. Discussion
4. Materials and Methods
4.1. Collection of Plant Material
4.2. Extraction of Essential Oils
4.3. Rearing of Mosquitoes
4.4. Mosquito Repellency Bioassay
4.5. Time-Span Bioassays
4.6. Oviposition Deterrence
4.7. Chemical Analysis of EOs
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benelli, G.; Duggan, M.F. Management of arthropod vector data–Social and ecological dynamics facing the One Health perspective. Acta Trop. 2018, 182, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Gresh, L.; Vargas, M.J.; Ballesteros, G.; Tellez, Y.; Soda, K.J.; Sahoo, M.K.; Nuñez, A.; Balmaseda, A.; Harris, E. Viremia and clinical presentation in Nicaraguan patients infected with Zika virus, chikungunya virus, and dengue virus. Clin. Infect. Dis. 2016, 1, ciw589. [Google Scholar] [CrossRef] [PubMed]
- Kesete, Y.; Mhretab, S.; Tesfay, M. Prevalence of malaria from blood smears examination: A three-year retrospective study from Nakfa Hospital, Eritrea. medRxiv 2020, 2, 20–28. [Google Scholar]
- Sutthanont, N.; Attrapadung, S.; Nuchprayoon, S. Larvicidal activity of synthesized silver nanoparticles from Curcuma zedoaria essential oil against Culex quinquefasciatus. Insects 2019, 10, 27. [Google Scholar] [CrossRef]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Produc. 2021, 283, 124–133. [Google Scholar] [CrossRef]
- Andreazza, F.; Oliveira, E.E.; Martins, G.F. Implications of sublethal insecticide exposure and the development of resistance on mosquito physiology, behavior, and pathogen transmission. Insects 2021, 12, 917. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.; Potter, C. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar. J. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Qiu, H.; Jun, H.W.; Dzimianski, M.; McCall, J. Reduced transdermal absorption of N, N-diethyl-m-toluamide from a new topical insect repellent formulation. Pharm. Dev. Technol. 1997, 2, 33–42. [Google Scholar] [CrossRef]
- Calafat, A.M.; Baker, S.E.; Wong, L.-Y.; Bishop, A.M.; Morales-A, P.; Valentin-Blasini, L. Novel exposure biomarkers of N, N-diethyl-m-toluamide (DEET): Data from the 2007–2010 National Health and Nutrition Examination Survey. Environ. Int. 2016, 92, 398–404. [Google Scholar] [CrossRef]
- Oftadeh, M.; Sendi, J.J.; Ebadollahi, A.; Setzer, W.N.; Krutmuang, P. Mulberry protection through flowering-stage essential oil of Artemisia annua against the lesser mulberry pyralid, Glyphodes pyloalis Walker. Foods 2021, 10, 210. [Google Scholar] [CrossRef]
- Manh, H.D.; Tuyet, O.T. Larvicidal and repellent activity of Mentha arvensis L. essential oil against Aedes aegypti. Insects 2020, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.G.; Haris, A.; Binyameen, M.; Nazir, A.; Mozūratis, R.; Azeem, M. Chemical Composition, Larvicidal and Repellent Activities of Wild Plant Essential Oils against Aedes Aegypti. Biology 2023, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Rao, R.; Kumar, S.; Mahant, S.; Khatkar, S. Therapeutic potential of citronella essential oil: A review. Curr. Drug Discov. Technol. 2019, 16, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.P.; Loye, J. PMD, a registered botanical mosquito repellent with deet-like efficacy. J. Am. Mosq. Control Assoc. 2006, 22, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Amer, A.; Mehlhorn, H. Repellency effect of forty-one essential oils against Aedes, Anopheles, and Culex mosquitoes. Parasitol. Res. 2006, 99, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Sivakumar, R.; Rajeswari, M.; Yogalakshmi, K. Chemical composition and larvicidal activity of essential oil from Mentha spicata (Linn.) against three mosquito species. Parasitol. Res. 2012, 110, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Negahban, M.; Moharramipour, S.; Sefidkon, F. Fumigant toxicity of essential oil from Artemisia sieberi Besser against three stored-product insects. J. Stored Prod. Res. 2007, 43, 123–128. [Google Scholar] [CrossRef]
- Kweka, E.J.; Munga, S.; Mahande, A.M.; Msangi, S.; Mazigo, H.D.; Adrias, A.Q.; Matias, J.R. Protective efficacy of menthol propylene glycol carbonate compared to N, N-diethyl-methylbenzamide against mosquito bites in Northern Tanzania. Parasit. Vectors 2012, 5, 1–10. [Google Scholar] [CrossRef]
- Haris, A.; Azeem, M.; Abbas, M.G.; Mumtaz, M.; Mozūratis, R.; Binyameen, M. Prolonged repellent activity of plant essential oils against dengue vector, Aedes aegypti. Molecules 2023, 28, 1351. [Google Scholar] [CrossRef]
- Dua, V.; Gupta, N.; Pandey, A.; Sharma, V. Repellency of Lantana camara (Verbenaceae) flowers against Aedes mosquitoes. J. Am. Mosq. Control Assoc. 1996, 12, 406–408. [Google Scholar] [PubMed]
- Keziah, E.A.; Nukenine, E.N.; Danga, S.P.Y.; Younoussa, L.; Esimone, C.O. Creams formulated with Ocimum gratissimum L. and Lantana camara L. crude extracts and fractions as mosquito repellents against Aedes aegypti L.(Diptera: Culicidae). J. Insect Sci. 2015, 15, 123–135. [Google Scholar] [CrossRef]
- Bhargava, S.; Agrawal, D.; Agrawal, O. Repellent activity of essential oil and leaf extract of Lantana camara L. in laboratory condition. Int. J. Theor. Appl. Sci. 2013, 5, 170–174. [Google Scholar]
- Azeem, M.; Zaman, T.; Tahir, M.; Haris, A.; Iqbal, Z.; Binyameen, M.; Nazir, A.; Shad, S.A.; Majeed, S.; Mozūraitis, R. Chemical composition and repellent activity of native plants essential oils against dengue mosquito. Aedes Aegypti. Ind. Crops Prod. 2019, 140, 111–119. [Google Scholar] [CrossRef]
- Iqbal, S.; Khan, F.A.; Haris, A.; Mozūratis, R.; Binyameen, M.; Azeem, M. Essential oils of four wild plants inhibit the blood seeking behaviour of female Aedes aegytpi. Exp. Parasitol. 2023, 244, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Chen, B.; McClements, D.J. Improving the efficacy of essential oils as antimicrobials in foods: Mechanisms of action. Ann. Rev. Food Sci. Technol. 2019, 10, 365–387. [Google Scholar] [CrossRef] [PubMed]
- Laznik, Z.; Vidrih, M.; Trdan, S. Efficacy of four essential oils against Sitophilus granarius (L.) adults after short-term exposure. Afr. J. Agric. Res. 2012, 7, 3175–3181. [Google Scholar]
- Nenaah, G.E.; Almadiy, A.A.; Al-Assiuty, B.A.; Mahnashi, M.H. The essential oil of Schinus terebinthifolius and its nanoemulsion and isolated monoterpenes: Investigation of their activity against Culex pipiens with insights into the adverse effects on non-target organisms. Pest Manag. Sci. 2022, 78, 1035–1047. [Google Scholar] [CrossRef]
- Parveen, A.; Abbas, M.G.; Keefover-Ring, K.; Binyameen, M.; Mozūraitis, R.; Azeem, M. Chemical Composition of Essential Oils from Natural Populations of Artemisia scoparia Collected at Different Altitudes: Antibacterial, Mosquito Repellent, and Larvicidal Effects. Molecules 2024, 29, 1359. [Google Scholar] [CrossRef]
- Klocke, J.A.; Darlington, M.V.; Balandrin, M.F. 1, 8-Cineole (Eucalyptol), a mosquito feeding and ovipositional repellent from volatile oil of Hemizonia fitchii (Asteraceae). J. Chem. Ecol. 1987, 13, 2131–2141. [Google Scholar] [CrossRef]
- Okoli, B.J.; Ladan, Z.; Mtunzi, F.; Hosea, Y.C. Vitex negundo L. Essential oil: Odorant binding protein efficiency using molecular docking approach and studies of the mosquito repellent. Insects 2021, 12, 1061. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Papachristos, D.P.; Kimbaris, A.; Koliopoulos, G.; Polissiou, M.G.; Emmanouel, N.; Michaelakis, A. Evaluation of bioefficacy of three Citrus essential oils against the dengue vector Aedes albopictus (Diptera: Culicidae) in correlation to their components enantiomeric distribution. Parasitol. Res. 2012, 111, 2253–2263. [Google Scholar] [CrossRef]
- Yadav, R.; Tyagi, V.; Tikar, S.N.; Sharma, A.K.; Mendki, M.J.; Jain, A.K.; Sukumaran, D. Differential larval toxicity and oviposition altering activity of some indigenous plant extracts against dengue and chikungunya vector Aedes albopictus. Entomol. Res. 2014, 2, 212–220. [Google Scholar]
- Hamzavi, F.; Moharramipour, S. Chemical composition and antifeedant activity of essential oils from Eucalyptus camaldulensis and Callistemon viminalis on Tribolium confusum. Int. J. Agric. Technol. 2017, 13, 413–424. [Google Scholar]
- Sales, T.A.; Cardoso, M.D.G.; Guimarães, L.G.D.L.; Camargo, K.C.; Rezende, D.A.; Brandão, R.M.; Souza, R.V.; Ferreira, V.R.; Marques, A.E.; Magalhães, M.L. Essential oils from the leaves and flowers of Callistemon viminalis: Chemical characterization and evaluation of the insecticide and antifungal activities. Am. J. Plant Sci. 2017, 8, 2516. [Google Scholar] [CrossRef]
- Ndomo, A.; Tapondjou, L.A.; Ngamo, L.; Hance, T. Insecticidal activities of essential oil of Callistemon viminalis applied as fumigant and powder against two bruchids. J. Appl. Entomol. 2010, 134, 333–341. [Google Scholar] [CrossRef]
- Ocheng, F.; Bwanga, F.; Joloba, M.; Softrata, A.; Azeem, M.; Pütsep, K.; Borg-Karlson, A.-K.; Obua, C.; Gustafsson, A. Essential oils from ugandan aromatic medicinal plants: Chemical composition and growth inhibitory effects on oral pathogens. Evid. Based Complement. Altern. Med. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Lawal, O.A.; Ogunwande, I.A.; Kasali, A.A.; Opoku, A.R.; Oyedeji, A.O. Chemical composition, antibacterial and cytotoxic activities of essential oil from the leaves of Helichrysum odoratissimum grown in South Africa. J. Essent. Oil Bear. Plants 2015, 18, 236–241. [Google Scholar] [CrossRef]
- Zantanta, N.; Kambizi, L.; Etsassala, N.G.; Nchu, F. Comparing crop yield, secondary metabolite contents, and antifungal activity of extracts of Helichrysum odoratissimum cultivated in aquaponic, hydroponic, and field systems. Plants 2022, 11, 2696. [Google Scholar] [CrossRef]
- Ocheng, F.; Bwanga, F.; Joloba, M.; Borg-Karlson, A.-K.; Gustafsson, A.; Obua, C. Antibacterial activities of extracts from Ugandan medicinal plants used for oral care. J. Ethnopharmacol. 2014, 155, 852–855. [Google Scholar] [CrossRef]
- Benelli, G.; Flamini, G.; Fiore, G.; Cioni, P.L.; Conti, B. Larvicidal and repellent activity of the essential oil of Coriandrum sativum L. (Apiaceae) fruits against the filariasis vector Aedes albopictus Skuse (Diptera: Culicidae). Parasitol. Res. 2013, 112, 1155–1161. [Google Scholar] [CrossRef]
- Abagli, A.Z.; Alavo, T.B.C. Essential oil from bush mint, Hyptis suaveolens, is as effective as DEET for personal protection against mosquito bites. Open Entomol. J. 2011, 5, 232–243. [Google Scholar] [CrossRef]
- Dua, V.; Pandey, A.; Dash, A. Adulticidal activity of essential oil of Lantana camara leaves against mosquitoes. Indian J. Med. Res. 2010, 131, 434–439. [Google Scholar]
- Nea, F.; Kambiré, D.A.; Genva, M.; Tanoh, E.A.; Wognin, E.L.; Martin, H.; Brostaux, Y.; Tomi, F.; Lognay, G.C.; Tonzibo, Z.F. Composition, seasonal variation, and biological activities of Lantana camara essential oils from Côte d’Ivoire. Molecules 2020, 25, 2400. [Google Scholar] [CrossRef]
- El-Sabrout, A.M.; Zoghroban, A.A.; Abdelgaleil, S.A. Chemical composition and effects of four essential oils on mortality, development and physiology of the West Nile virus vector, Culex pipiens. Int. J. Trop. Insect Sci. 2020, 40, 789–799. [Google Scholar] [CrossRef]
- Srivastava, S.; Ahmad, A.; Syamsunder, K.; Aggarwal, K.; Khanuja, S. Essential oil composition of Callistemon viminalis leaves from India. Flavour Fragr. J. 2003, 18, 361–363. [Google Scholar] [CrossRef]
- Mubarak, E.E.; Mohajer, S.; Ahmed, I.; Taha, R.M. Essential oil compositions from leaves of Eucalyptus camaldulensis Dehn. and Callistemon viminalis Originated from Malaysia. IPCBEE 2014, 70, 137–141. [Google Scholar]
- Oliveira, J.A.D.; Garcia, I.P.; Corrêa, E.J.A.; de Lima, L.H.F.; Santos, H.D.; de Assis, R.M.A.; Pinto, J.; Bertolucci, S.K.V. Larvicidal susceptibility of essential oils from Cinnamodendron dinisii, Callistemon viminalis and Myrcia tomentosa against Culex quinquefasciatus (Say) (Diptera: Culicidae). South Afr. J. Bot. 2023, 163, 95–104. [Google Scholar] [CrossRef]
- Silva, A.G.; Almeida, D.L.; Ronchi, S.N.; Bento, A.C.; Scherer, R.; Ramos, A.C.; Cruz, Z.M. The essential oil of Brazilian pepper, Schinus terebinthifolia Raddi in larval control of Stegomyia aegypti (Linnaeus, 1762). Parasit. Vectors 2010, 3, 79. [Google Scholar] [CrossRef]
- Kweka, E.J.; Nyindo, M.; Mosha, F.; Silva, A.G. Insecticidal activity of the essential oil from fruits and seeds of Schinus terebinthifolia Raddi against African malaria vectors. Parasit. Vectors 2011, 4, 129. [Google Scholar] [CrossRef]
- Hussein, H.S.; Salem, M.Z.; Soliman, A.M. Repellent, attractive, and insecticidal effects of essential oils from Schinus terebinthifolius fruits and Corymbia citriodora leaves on two whitefly species, Bemisia tabaci, and Trialeurodes ricini. Sci. Hortic. 2017, 216, 111–119. [Google Scholar] [CrossRef]
- Belhoussaine, O.; El Kourchi, C.; Harhar, H.; Bouyahya, A.; El Yadini, A.; Fozia, F.; Alotaibi, A.; Ullah, R.; Tabyaoui, M. Chemical composition, antioxidant, insecticidal activity, and comparative analysis of essential oils of leaves and fruits of Schinus molle and Schinus terebinthifolius. Evid. Based Complement. Altern. Med. 2022, 2022, 4288890. [Google Scholar] [CrossRef]
- Wangrawa, D.W.; Badolo, A.; Guelbeogo, W.M.; Nebie, R.C.H.; Sagnon, N.F.; Borovsky, D.; Sanon, A. Larvicidal, oviposition-deterrence, and excito-repellency activities of four essential oils: An eco-friendly tool against malaria vectors Anopheles coluzzii and Anopheles gambiae (Diptera: Culicidae). Int. J. Trop. Insect Sci. 2021, 41, 1771–1781. [Google Scholar] [CrossRef]
- Koul, O.; Singh, R.; Kaur, B.; Kanda, D. Comparative study on the behavioral response and acute toxicity of some essential oil compounds and their binary mixtures to larvae of Helicoverpa armigera, Spodoptera litura and Chilo partellus. Ind. Crops Prod. 2013, 49, 428–436. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition deterrent and larvicidal and pupaecidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism–antagonism effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef]
- Soonwera, M.; Phasomkusolsil, S. Adulticidal, larvicidal, pupicidal and oviposition deterrent activities of essential oil from Zanthoxylum limonella Alston (Rutaceae) against Aedes aegypti (L.) and Culex quinquefasciatus (Say). Asian Pac. J. Trop. Biomed. 2017, 7, 967–978. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Kramer, W.L.; Mulla, M.S. Oviposition attractants and repellents of mosquitoes: Isolation and identification of oviposition repellents for Culex mosquitoes. J. Chem. Ecol. 1980, 6, 71–80. [Google Scholar] [CrossRef]
- Davis, E.E. A receptor sensitive to oviposition site attractants on the antennae of the mosquito, Aedes aegypti. J. Insect Physiol. 1976, 22, 1371–1376. [Google Scholar] [CrossRef]
- Bhujel, P.; Saha, D. Rearing Protocol for Culex quinquefasciatus. Sci. Vis. 2023, 2023, 61–65. [Google Scholar]
- Zheng, M.-L.; Zhang, D.-J.; Damiens, D.D.; Lees, R.S.; Gilles, J.R. Standard operating procedures for standardized mass rearing of the dengue and chikungunya vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)-II-Egg storage and hatching. Parasit. Vectors 2015, 8, 348. [Google Scholar] [CrossRef]
- Govindarajan, M.; Sivakumar, R.; Rajeswary, M.; Yogalakshmi, K. Chemical composition and larvicidal activity of essential oil from Ocimum basilicum (L.) against Culex tritaeniorhynchus, Aedes albopictus and Anopheles subpictus (Diptera: Culicidae). Exp. Parasitol. 2013, 134, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Soonwera, M. Larvicidal and oviposition deterrent activities of essential oils against house fly (Musca domestica L.; Diptera: Muscidae). J. Agric. Technol. 2015, 11, 657–667. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Family | Location Coordinates | Part Used | Yield (%) |
|---|---|---|---|---|
| Lantana camara L. | Verbenaceae | 30°20′3.6″ N 71°56′24″ E | Leaves | 0.19 |
| Schinus terebinthifolia Raddi. | Anacardiaceae | 30°14′24″ N 71°26′24″ E | Leaves | 0.09 |
| Callistemon viminalis (Sol. ex Gaertn.) G.Don | Myrtaceae | 30°20′24″ N 71°31′48″ E | Leaves, stems | 0.21 |
| Helichrysum odoratissimum L. | Asteraceae | 0°19′32.8″ S 31°47′01.0″ E | Aerial parts | 0.29 |
| Hyptis suaveolens (L.) Poit. | Lamiaceae | 30°19′12″ N 70°58′4.8″ E | Leaves | 0.27 |
| Retention Index | Compounds | L. camara | S. terebinthifolia | C. viminalis |
|---|---|---|---|---|
| 922 | α-Thujene | 1.1 | 0.6 | 2.8 |
| 928 | α-Pinene | 4.3 | 15.5 | 34.2 |
| 942 | Camphene | 1.9 | 0.3 | 0.1 |
| 969 | Sabinene | 16.5 | 0.3 | |
| 971 | β-Pinene | 1.9 | 1.2 | 1.2 |
| 988 | β-Myrcene | 1.9 | 1 | 0.1 |
| 1001 | α-Phellandrene | 0.2 | 12.4 | 2.9 |
| 1007 | 3-Carene | 2.4 | 0.2 | |
| 1013 | α-Terpinene | 0.6 | 0.7 | 0.1 |
| 1021 | p-Cymene | 1.8 | 11.5 | 9 |
| 1026 | Limonene | 3.2 | 14 | 2.3 |
| 1027 | 1,8-Cineole | 13.1 | 38.1 | |
| 1036 | cis-β-Ocimene | 0.2 | 0.1 | |
| 1056 | γ-Terpinene | 1 | 0.5 | 1.2 |
| 1064 | cis-Sabinenhydrate | 0.8 | ||
| 1086 | α-Terpinolene | 0.3 | 0.8 | 0.5 |
| 1095 | trans-Sabinene hydrate | 0.2 | ||
| 1099 | Linalool | 0.1 | 0.1 | 0.3 |
| 1103 | 2-Methylbutyl 2-methylbutanoate | 0.2 | 0.1 | |
| 1135 | trans-Pinocarveol | Tr | 0.2 | 0.1 |
| 1140 | Camphor | 1 | 0.1 | |
| 1163 | Borneol | 0.7 | ||
| 1175 | 4-Terpineol | 1.1 | 0.4 | 0.5 |
| 1183 | Cryptone | Tr | 0.6 | |
| 1188 | α-Terpineol | 0.6 | 4 | |
| 1199 | Sabinol | 1.1 | ||
| 1205 | Verbenone | 0.3 | ||
| 1218 | trans-Carveol | 0.2 | ||
| 1250 | cis-Ascaridol | 0.2 | ||
| 1272 | Phellandral | 0.2 | ||
| 1302 | Carvacrol | 0.6 | ||
| 1337 | δ-Elemene | 0.1 | 0.5 | |
| 1349 | α-Cubebene | 0.2 | 0.1 | |
| 1375 | α-Copaene | 0.4 | 0.2 | |
| 1384 | β-Bourbonene | 0 | 0.5 | |
| 1389 | β-Cubebene | 0.4 | ||
| 1391 | β-Elemene | 0.2 | 4.5 | |
| 1419 | trans-β-Caryophyllene | 16.7 | 2 | 0.3 |
| 1428 | β-Gurjunene | 0.4 | 0.2 | |
| 1433 | γ-Elemene | 0.2 | 2.8 | |
| 1438 | Aromadendrene | 0 | 0.2 | 0.1 |
| 1443 | γ-Gurjunene | 0 | 0.1 | |
| 1453 | α-Humulene | 8.6 | 0.3 | 0.1 |
| 1460 | Alloaromadendrene | 0.3 | 0.1 | |
| 1476 | γ-Muurolene | 0.3 | ||
| 1480 | Germacrene D | 0.4 | 3.4 | |
| 1485 | β-Selinene | 0 | 0.6 | |
| 1496 | Elixene | 1 | 5.1 | |
| 1500 | α-Muurolene | 0.2 | ||
| 1504 | α-Selinene | 0.2 | ||
| 1523 | δ-Cadinene | 0.2 | 0.6 | |
| 1557 | Davanone | 1.2 | 2.8 | |
| 1564 | trans-Nerolidol | 5.5 | ||
| 1577 | Spathulenol | 1.6 | 8.4 | |
| 1577 | Ledene alcohol | 0 | 0.7 | |
| 1586 | Caryophyllene oxide | 0.5 | ||
| 1629 | Alloaromadendrene oxide | 4.7 | 0.4 | |
| 1638 | β-Cedren-9-α-ol | 0.4 | ||
| 1645 | Nordracorhodin | 0.1 | ||
| 1655 | τ-Cadinol | 0.4 | ||
| 1658 | Longifolenaldehyde | 0.5 | ||
| 1672 | Aromadendrene oxide | 0.4 | ||
| 1675 | Ledene oxide | 0.2 | ||
| 1704 | Santalol | 0.1 | ||
| 1708 | Cedren-13-ol | 0.1 | ||
| 1719 | Isoaromadendrene epoxide | 0.3 | ||
| 1754 | Globulol | 0.1 | ||
| Total Identified % | 99.2 | 97.3 | 99.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, M.G.; Azeem, M.; Bashir, M.U.; Ali, F.; Mozūratis, R.; Binyameen, M. Chemical Composition, Repellent, and Oviposition Deterrent Potential of Wild Plant Essential Oils against Three Mosquito Species. Molecules 2024, 29, 2657. https://doi.org/10.3390/molecules29112657
Abbas MG, Azeem M, Bashir MU, Ali F, Mozūratis R, Binyameen M. Chemical Composition, Repellent, and Oviposition Deterrent Potential of Wild Plant Essential Oils against Three Mosquito Species. Molecules. 2024; 29(11):2657. https://doi.org/10.3390/molecules29112657
Chicago/Turabian StyleAbbas, Muhammad Ghazanfar, Muhammad Azeem, Muhammad Umar Bashir, Fawad Ali, Raimondas Mozūratis, and Muhammad Binyameen. 2024. "Chemical Composition, Repellent, and Oviposition Deterrent Potential of Wild Plant Essential Oils against Three Mosquito Species" Molecules 29, no. 11: 2657. https://doi.org/10.3390/molecules29112657
APA StyleAbbas, M. G., Azeem, M., Bashir, M. U., Ali, F., Mozūratis, R., & Binyameen, M. (2024). Chemical Composition, Repellent, and Oviposition Deterrent Potential of Wild Plant Essential Oils against Three Mosquito Species. Molecules, 29(11), 2657. https://doi.org/10.3390/molecules29112657