
Computational Screening and Experimental Validation of Inhibitor Targeting the Complex Formation of Grb14 and Insulin Receptor

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
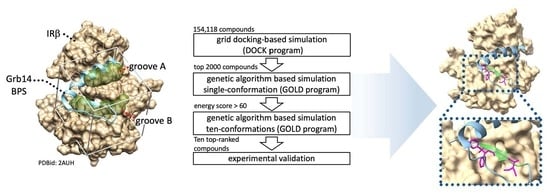
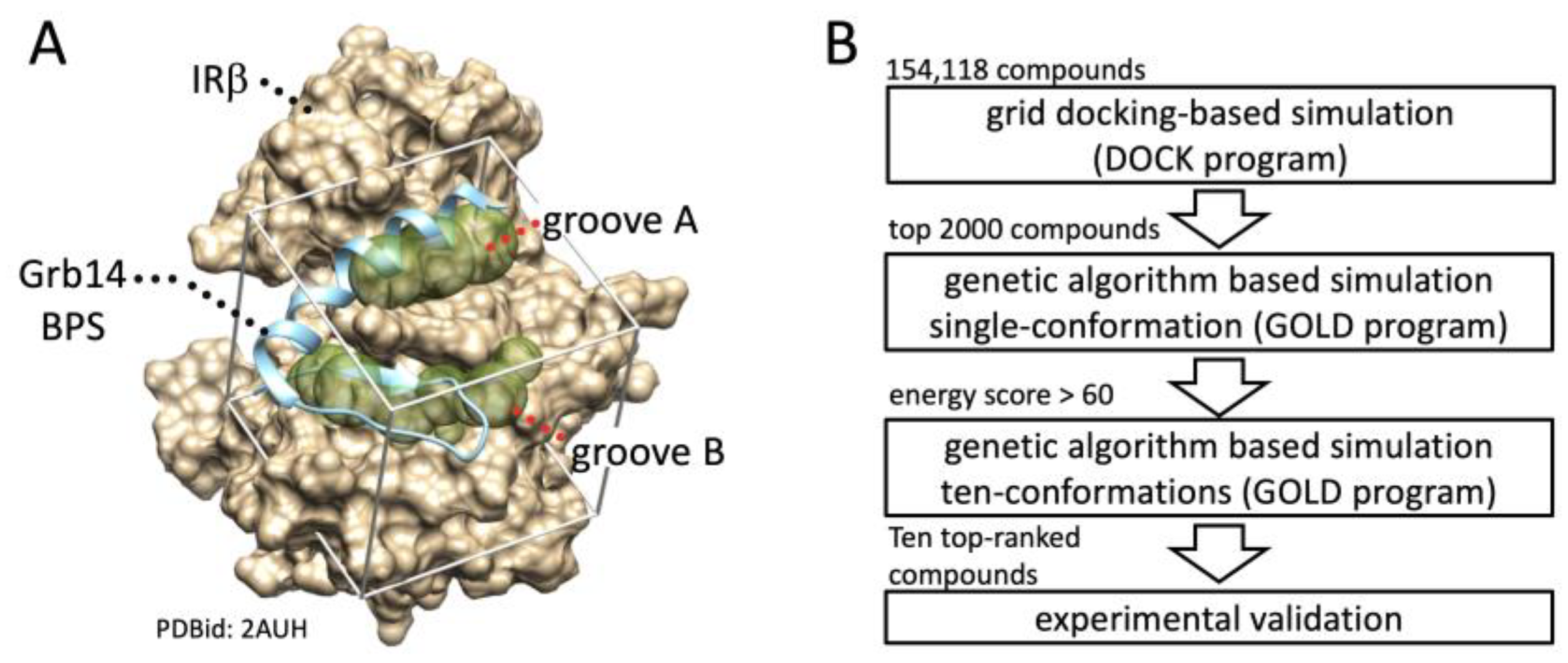
2.1. In Silico Identification of Small Compounds Inhibiting Grb14-IRβ Complex Formation
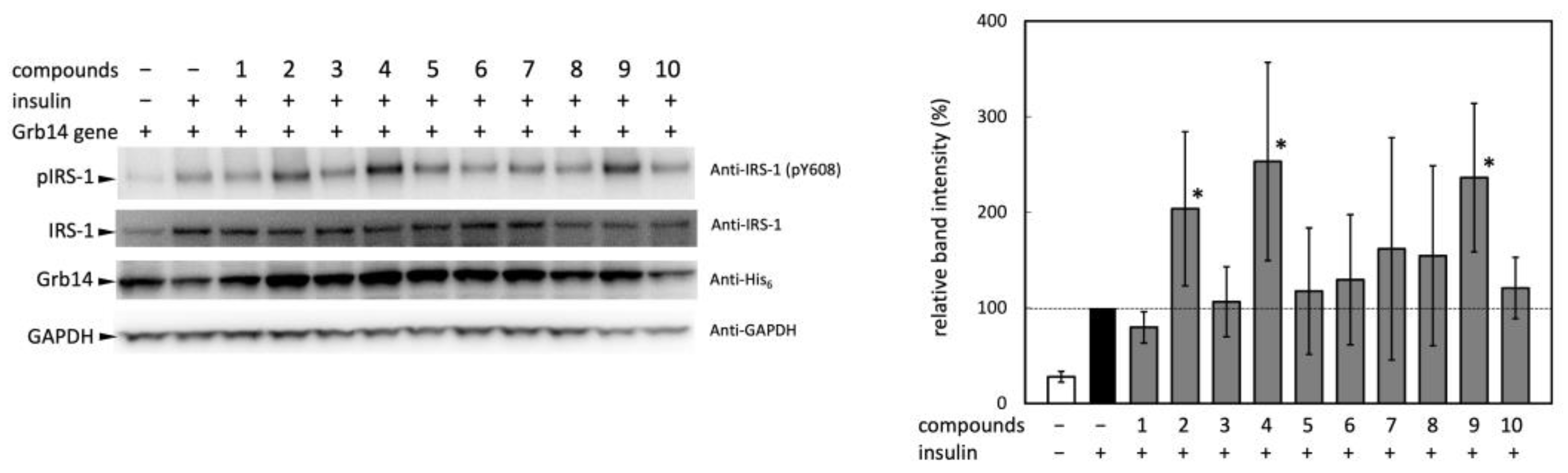
2.2. Experimental Validation of the Effect of Selected Compounds on IRS-1 Activity
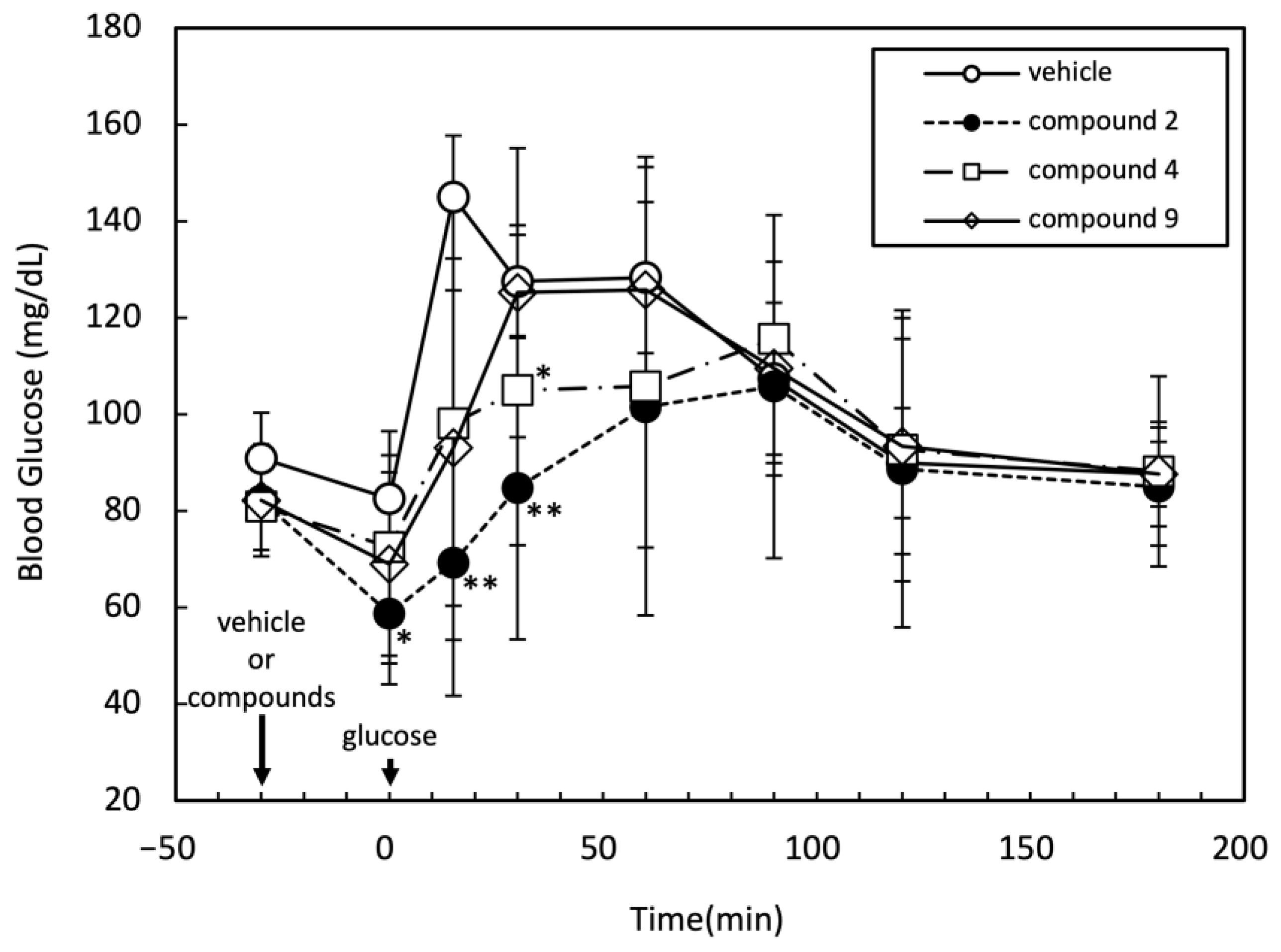
2.3. Effects of Three Candidate Compounds on Blood Glucose Levels
2.4. Effect of Compound 2 on IRβ–Grb14 Complex Formation
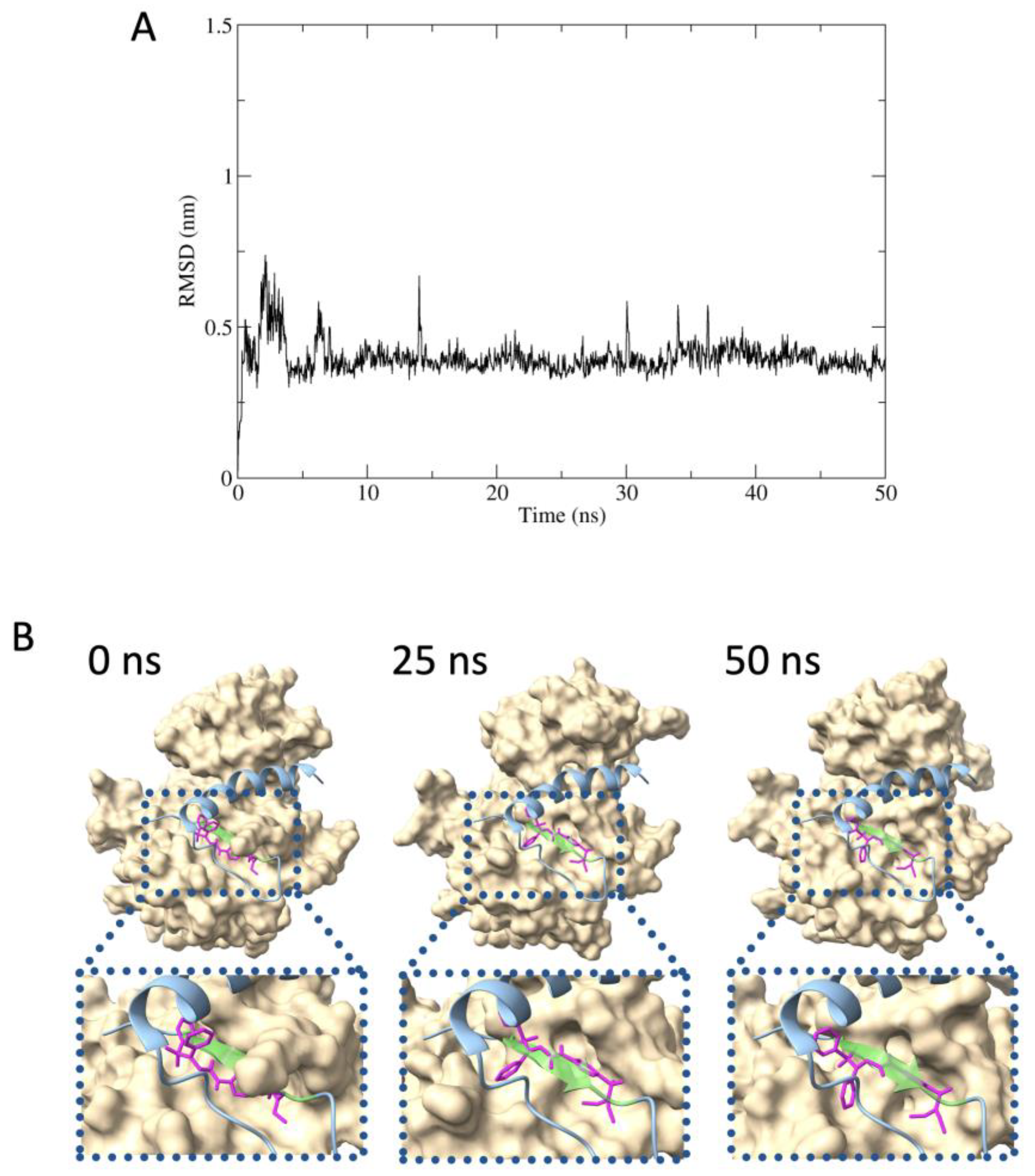
2.5. Predicted Protein–Inhibitor Interaction by MD Simulation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. In Silico Structure-Based Drug Screening
4.3. Phosphorylation of IRS-1
4.4. Oral Glucose Tolerance Test
4.5. Co-Immunoprecipitation (Co-IP)
4.6. Molecular Dynamics Simulation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Norris, J.M.; Rich, S.S. Genetics of glucose homeostasis: Implications for insulin resistance and metabolic syndrome. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2091–2096. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Walker, M. Genetics of insulin resistance and the metabolic syndrome. Curr. Cardiol. Rep. 2016, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, R.A.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2018, 19, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Artasensi, A.; Pedretti, A.; Vistoli, G.; Fumagalli, L. Type 2 diabetes mellitus: A review of multi-target drugs. Molecules 2020, 25, 1987. [Google Scholar] [CrossRef] [PubMed]
- Demir, S.; Nawroth, P.P.; Herzig, S.; Ekim Ustunel, B. Emerging targets in type 2 diabetes and diabetic complications. Adv. Sci. 2021, 8, e2100275. [Google Scholar] [CrossRef]
- Cheng, T.; Li, Q.; Zhou, Z.; Wang, Y.; Bryant, S.H. Structure-based virtual screening for drug discovery: A problem-centric review. AAPS J. 2012, 14, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.J.; Sanderson, G.M.; Janes, P.W.; Sutherland, R.L. Cloning and characterization of GRB14, a novel member of the GRB7 gene family. J. Biol. Chem. 1996, 271, 12502–12510. [Google Scholar] [CrossRef]
- Kasus-Jacobi, A.; Perdereau, D.; Auzan, C.; Clauser, E.; Van Obberghen, E.; Mauvais-Jarvis, F.; Girard, J.; Burnol, A.F. Identification of the rat adapter Grb14 as an inhibitor of insulin actions. J. Biol. Chem. 1998, 273, 26026–26035. [Google Scholar] [CrossRef]
- Nouaille, S.; Blanquart, C.; Zilberfarb, V.; Boute, N.; Perdereau, D.; Burnol, A.F.; Issad, T. Interaction between the insulin receptor and Grb14: A dynamic study in living cells using BRET. Biochem. Pharmacol. 2006, 72, 1355–1366. [Google Scholar] [CrossRef]
- Nouaille, S.; Blanquart, C.; Zilberfarb, V.; Boute, N.; Perdereau, D.; Roix, J.; Burnol, A.F.; Issad, T. Interaction with Grb14 results in site-specific regulation of tyrosine phosphorylation of the insulin receptor. EMBO Rep. 2006, 7, 512–518. [Google Scholar] [CrossRef]
- Cooney, G.J.; Lyons, R.J.; Crew, A.J.; Jensen, T.E.; Molero, J.C.; Mitchell, C.J.; Biden, T.J.; Ormandy, C.J.; James, D.E.; Daly, R.J. Improved glucose homeostasis and enhanced insulin signalling in Grb14-deficient mice. EMBO J. 2004, 23, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Cariou, B.; Capitaine, N.; Le Marcis, V.; Vega, N.; Bereziat, V.; Kergoat, M.; Laville, M.; Girard, J.; Vidal, H.; Burnol, A.F. Increased adipose tissue expression of Grb14 in several models of insulin resistance. FASEB J. 2004, 18, 965–967. [Google Scholar] [CrossRef]
- Kooner, J.S.; Saleheen, D.; Sim, X.; Sehmi, J.; Zhang, W.; Frossard, P.; Been, L.F.; Chia, K.S.; Dimas, A.S.; Hassanali, N.; et al. Genome-wide association study in individuals of South Asian ancestry identifies six new type 2 diabetes susceptibility loci. Nat. Genet. 2011, 43, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Rahmioglu, N.; Macgregor, S.; Drong, A.W.; Hedman, Å.; Harris, H.R.; Randall, J.C.; Prokopenko, I.; Nyholt, D.R.; Morris, A.P.; Montgomery, G.W.; et al. Genome-wide enrichment analysis between endometriosis and obesity-related traits reveals novel susceptibility loci. Hum. Mol. Genet. 2015, 24, 1185–1199. [Google Scholar] [CrossRef] [PubMed]
- Desbuquois, B.; Carre, N.; Burnol, A.F. Regulation of insulin and type 1 insulin-like growth factor signaling and action by the Grb10/14 and SH2B1/B2 adaptor proteins. FEBS J. 2013, 280, 794–816. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Rose, D.W.; Olefsky, J.M.; Gustafson, T.A. Grb10 interacts differentially with the insulin receptor, insulin-like growth factor I receptor, and epidermal growth factor receptor via the Grb10 Src homology 2 (SH2) domain and a second novel domain located between the pleckstrin homology and SH2 domains. J. Biol. Chem. 1998, 273, 6860–6867. [Google Scholar] [CrossRef] [PubMed]
- Depetris, R.S.; Hu, J.; Gimpelevich, I.; Holt, L.J.; Daly, R.J.; Hubbard, S.R. Structural basis for inhibition of the insulin receptor by the adaptor protein Grb14. Mol. Cell 2005, 20, 325–333. [Google Scholar] [CrossRef]
- Gondoin, A.; Hampe, C.; Eudes, R.; Fayolle, C.; Pierre-Eugene, C.; Miteva, M.; Villoutreix, B.O.; Charnay-Pouget, F.; Aitken, D.J.; Issad, T.; et al. Identification of insulin-sensitizing molecules acting by disrupting the interaction between the Insulin Receptor and Grb14. Sci. Rep. 2017, 7, 16901. [Google Scholar] [CrossRef]
- Taira, J.; Morita, K.; Kawashima, S.; Umei, T.; Baba, H.; Maruoka, T.; Komatsu, H.; Sakamoto, H.; Sacchettini, J.C.; Aoki, S. Identification of a novel class of small compounds with anti-tuberculosis activity by in silico structure-based drug screening. J. Antibiot. 2017, 70, 1057–1064. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, X.; Bi, X.; Huang, J.; Zhou, L. The insulin receptor: An important target for the development of novel medicines and pesticides. Int. J. Mol. Sci. 2022, 23, 7793. [Google Scholar] [CrossRef]
- Arkin, M.R.; Glicksman, M.A.; Fu, H.; Havel, J.J.; Du, Y. Inhibition of protein-protein interactions: Non-cellular assay formats. In Assay Guidance Manual; Markossian, S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C., Baell, J., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Shun, T.Y.; Lazo, J.S.; Sharlow, E.R.; Johnston, P. Identifying actives from HTS data sets: Practical approaches for the selection of an appropriate HTS data-processing method and quality control review. J. Biomol. Screen. 2010, 16, 1–14. [Google Scholar] [CrossRef]
- Dreiman, G.H.S.; Bictash, M.; Fish, P.V.; Griffin, L.; Svensson, F. Changing the HTS paradigm: AI-driven iterative screening for hit finding. SLAS Discov. 2021, 26, 257–262. [Google Scholar] [CrossRef]
- Taira, J.; Higashimoto, Y. Phosphorylation of Grb14 BPS domain by GSK-3 correlates with complex forming of Grb14 and insulin receptor. J. Biochem. 2014, 155, 353–360. [Google Scholar] [CrossRef]
- Bartsch, W.; Sponer, G.; Dietmann, K.; Fuchs, G. Acute toxicity of various solvents in the mouse and rat. LD50 of ethanol, diethylacetamide, dimethylformamide, dimethylsulfoxide, glycerine, N-methylpyrrolidone, polyethylene glycol 400, 1,2-propanediol and Tween 20. Arzneimittelforschung 1976, 26, 1581–1583. [Google Scholar]
- Taira, J.; Murakami, K.; Monobe, K.; Kuriki, K.; Fujita, M.; Ochi, Y.; Sakamoto, H.; Aoki, S. Identification of novel inhibitors for mycobacterial polyketide synthase 13 via in silico drug screening assisted by the parallel compound screening with genetic algorithm-based programs. J. Antibiot. 2022, 75, 552–558. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochi, Y.; Matsui, T.; Inoue, K.; Monobe, K.; Sakamoto, H.; Aoki, S.; Taira, J. Computational Screening and Experimental Validation of Inhibitor Targeting the Complex Formation of Grb14 and Insulin Receptor. Molecules 2024, 29, 198. https://doi.org/10.3390/molecules29010198
Ochi Y, Matsui T, Inoue K, Monobe K, Sakamoto H, Aoki S, Taira J. Computational Screening and Experimental Validation of Inhibitor Targeting the Complex Formation of Grb14 and Insulin Receptor. Molecules. 2024; 29(1):198. https://doi.org/10.3390/molecules29010198
Chicago/Turabian StyleOchi, Yosuke, Takanori Matsui, Keitaro Inoue, Kohei Monobe, Hiroshi Sakamoto, Shunsuke Aoki, and Junichi Taira. 2024. "Computational Screening and Experimental Validation of Inhibitor Targeting the Complex Formation of Grb14 and Insulin Receptor" Molecules 29, no. 1: 198. https://doi.org/10.3390/molecules29010198
APA StyleOchi, Y., Matsui, T., Inoue, K., Monobe, K., Sakamoto, H., Aoki, S., & Taira, J. (2024). Computational Screening and Experimental Validation of Inhibitor Targeting the Complex Formation of Grb14 and Insulin Receptor. Molecules, 29(1), 198. https://doi.org/10.3390/molecules29010198