

Research Progress Regarding the Effect and Mechanism of Dietary Polyphenols in Liver Fibrosis


Abstract
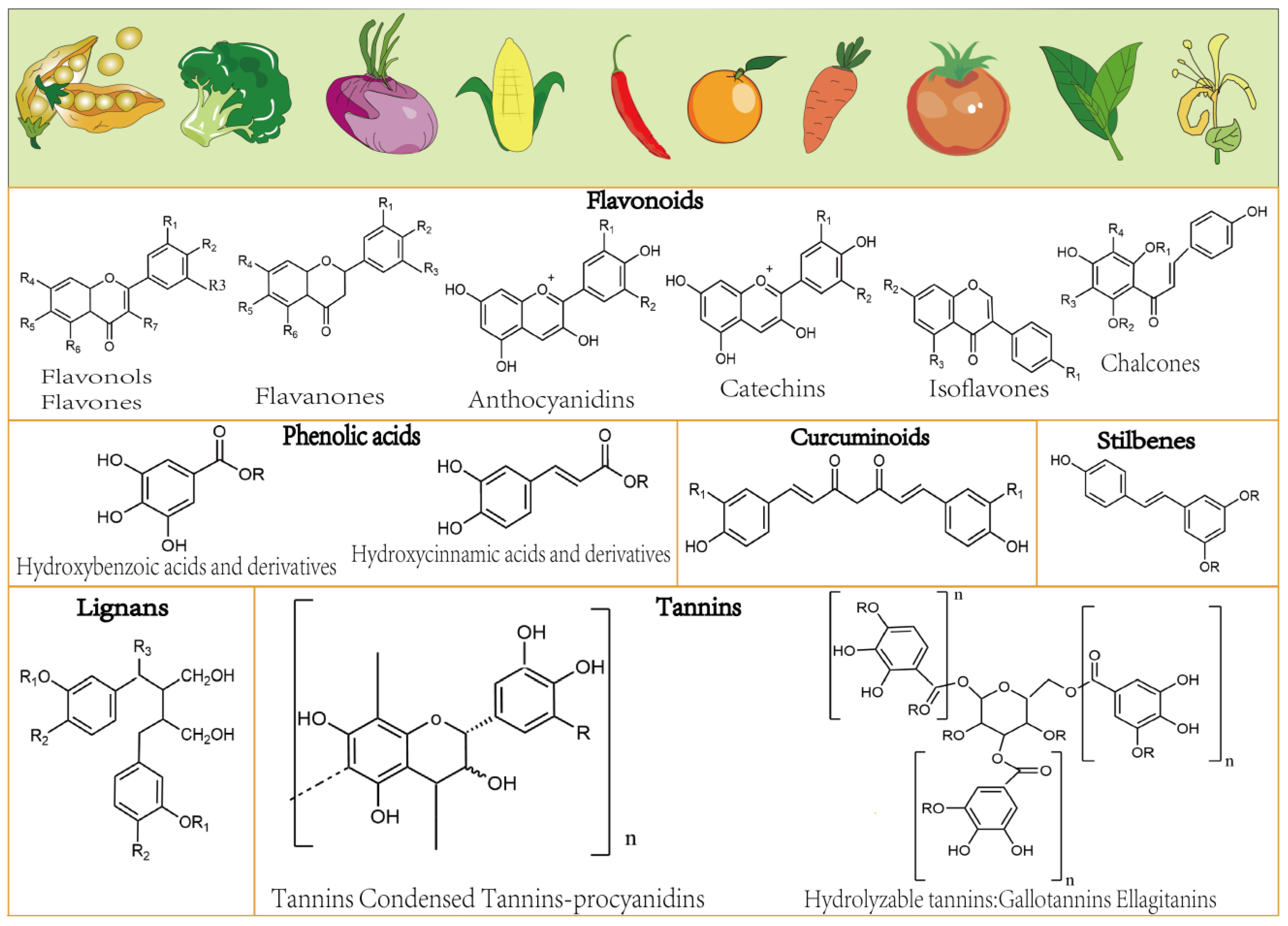
:1. Introduction
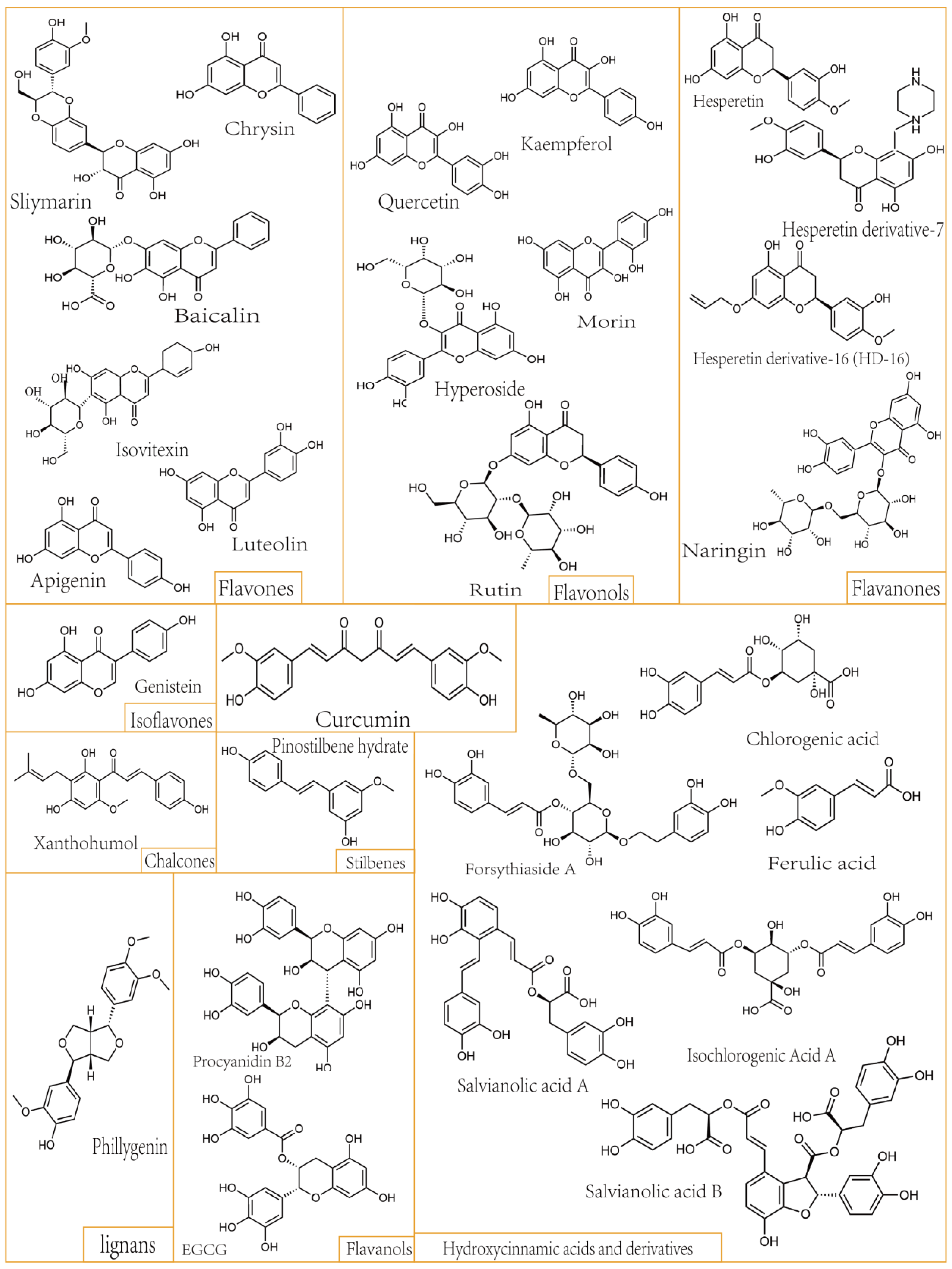
2. Dietary Polyphenols Affect Hepatic Fibrosis through Multiple Signaling Pathways
2.1. Dietary Polyphenols Reverse Hepatic Fibrosis via TGF-β/Smad Pathway
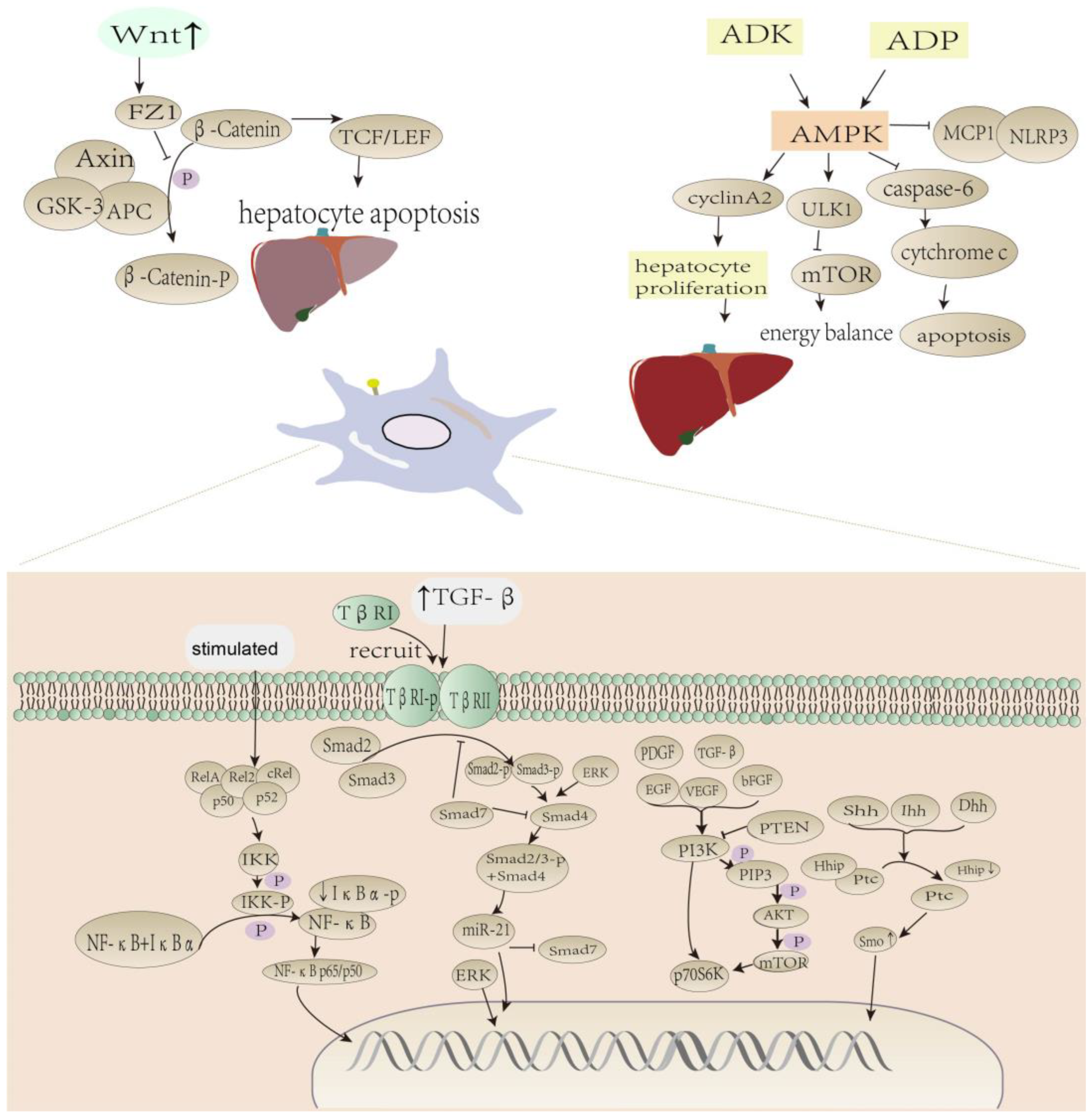
2.2. Dietary Polyphenols Activate AMPK/mTOR-Mediated Autophagic Pathway to Alleviate Liver Fibrosis
2.3. Dietary Polyphenols Reverse Wnt/β-Catenin Pathway in Hepatic Fibrosis
2.4. Polyphenols Inhibit the NF-κB Pathway in Liver Fibrosis
2.5. PI3K/AKT/mTOR Pathway Effects of Dietary Polyphenols
2.6. The Effect of Dietary Polyphenols on the Hedgehog Signaling Pathway
2.7. Role of Dietary Polyphenols on Liver Fibrosis-Related Factors
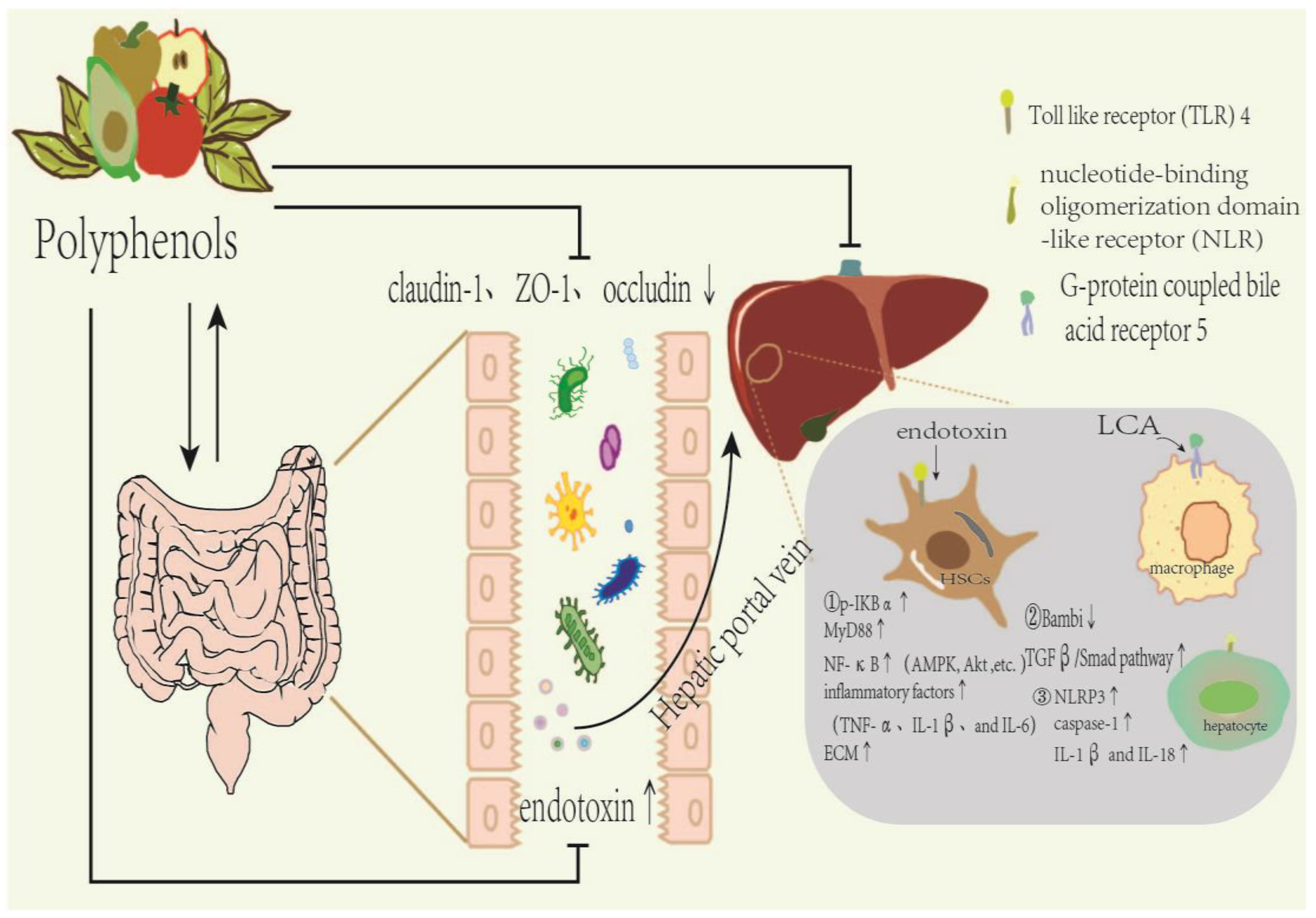
3. Dietary Polyphenols Affect Gut Microbiota Composition in Liver Fibrosis
3.1. Effect of Dietary Polyphenols on Intestinal Microbial Composition
3.2. Polyphenols Regulate Liver Fibrosis by Influencing Microbial Metabolites
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheemerla, S.; Balakrishnan, M. Global Epidemiology of Chronic Liver Disease. Clin. Liver Dis. 2021, 17, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Pinzani, M. Hepatic fibrosis 2022: Unmet needs and a blueprint for the future. Hepatology 2022, 75, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Wang, F.; Zhai, D.; Meng, X.; Liu, J.; Lv, X. New Drugs for Hepatic Fibrosis. Front. Pharmacol. 2022, 13, 874408. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Sun, H.; Xue, T.; Gan, C.; Liu, H.; Xie, Y.; Yao, Y.; Ye, T. Liver Fibrosis: Therapeutic Targets and Advances in Drug Therapy. Front. Cell Dev. Biol. 2021, 9, 730176. [Google Scholar] [CrossRef]
- Altamirano-Barrera, A.; Barranco-Fragoso, B.; Méndez-Sánchez, N. Management Strategies for Liver Fibrosis. Ann. Hepatol. 2017, 16, 48–56. [Google Scholar] [CrossRef]
- Pinheiro, D.; Dias, I.; Ribeiro Silva, K.; Stumbo, A.C.; Thole, A.; Cortez, E.; de Carvalho, L.; Weiskirchen, R.; Carvalho, S. Mechanisms Underlying Cell Therapy in Liver Fibrosis: An Overview. Cells 2019, 8, 1339. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Elfeky, M.G.; Mantawy, E.M.; Gad, A.M.; Fawzy, H.M.; El-Demerdash, E. Mechanistic aspects of antifibrotic effects of honokiol in Con A-induced liver fibrosis in rats: Emphasis on TGF-beta/SMAD/MAPK signaling pathways. Life Sci. 2020, 240, 117096. [Google Scholar] [CrossRef]
- Lee, I.H.; Im, E.; Lee, H.J.; Sim, D.Y.; Lee, J.H.; Jung, J.H.; Park, J.E.; Shim, B.S.; Kim, S.H. Apoptotic and antihepatofibrotic effect of honokiol via activation of GSK3beta and suppression of Wnt/beta-catenin pathway in hepatic stellate cells. Phytother. Res. 2021, 35, 452–462. [Google Scholar] [CrossRef]
- Gore, E.; Bigaeva, E.; Oldenburger, A.; Kim, Y.O.; Rippmann, J.F.; Schuppan, D.; Boersema, M.; Olinga, P. PI3K inhibition reduces murine and human liver fibrogenesis in precision-cut liver slices. Biochem. Pharmacol. 2019, 169, 113633. [Google Scholar] [CrossRef]
- Wang, J.N.; Li, L.; Li, L.Y.; Yan, Q.; Li, J.; Xu, T. Emerging role and therapeutic implication of Wnt signaling pathways in liver fibrosis. Gene 2018, 674, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Sara Lovisa, G.G.; Danese, S. Role of Epithelial-to-Mesenchymal Transition in Inflammatory Bowel Disease. J. Crohn’s Colitis 2018, 13, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.M.S.; Zaafan, M.A. The Hepatoprotective Effect of Piperine Against Thioacetamide-Induced Liver Fibrosis in Mice: The Involvement of miR-17 and TGF-beta/Smads Pathways. Front. Mol. Biosci. 2021, 8, 754098. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-T.; Wang, N.; Tan, H.Y.; Li, S.; Feng, Y. Targeting Hepatic Stellate Cells for the Treatment of Liver Fibrosis by Natural Products: Is It the Dawning of a New Era? Front. Pharmacol. 2020, 11, 548. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, M.; Zhao, Q.; Yu, Y. Prospective therapeutics for intestinal and hepatic fibrosis. Bioeng. Transl. Med. 2023, 8, e10579. [Google Scholar] [CrossRef] [PubMed]
- Tina Bormann, R.M.; Stolper, J.; Tarré, M.T.; Brandenberger, C.; Wedekind, D.; Jonigk, D.; Welte, T.; Gauldie, J.; Kolb, M.; Maus, U.A. Role of matrix metalloprotease-2 and MMP-9 in experimental lung fibrosis in mice. Respir. Res. 2022, 23, 180. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; D’Argenio, G.; Lembo, V.; Palumbo, A.; Castellano, I. Antifibrotic Effect of Marine Ovothiol in an In Vivo Model of Liver Fibrosis. Oxid. Med. Cell Longev. 2018, 2018, 5045734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Yuan, W.G.; He, P.; Lei, J.H.; Wang, C.X. Liver fibrosis and hepatic stellate cells: Etiology, pathological hallmarks and therapeutic targets. World J. Gastroenterol. 2016, 22, 10512–10522. [Google Scholar] [CrossRef]
- Ferro, D.B.F.; Pastori, D.; Cocomello, N.; Colantoni, A.; Angelico, F.; Del Ben, M. New Insights into the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Gut-Derived Lipopolysaccharides and Oxidative Stress. Nutrients 2020, 12, 2762. [Google Scholar] [CrossRef]
- Chenxu Ge, J.T.; Lou, D.; Zhu, L.; Zhong, Z.; Dai, X.; Sun, Y.; Kuang, Q.; Zhao, J.; Wang, L.; Liu, J.; et al. Mulberrin confers protection against hepatic fibrosis by Trim31/Nrf2 signaling. Redox Biol. 2022, 51, 102274. [Google Scholar]
- Schuppan, D.A.-K.; Yang, A.T.M.; Kim, Y.O. Liver fibrosis: Direct antifibrotic agents and targeted therapies. Matrix Biol. 2018, 68–69, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.M.; Wang, C.; Zhou, H.; Gong, L.; Zhang, Y.; Li, Y. Forsythiaside A alleviated carbon tetrachloride-induced liver fibrosis by modulating gut microbiota composition to increase short-chain fatty acids and restoring bile acids metabolism disorder. Biomed. Pharmacother. 2022, 151, 113185. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.F.Z.; Zhang, R.; Zhao, X. The effects and underlying mechanisms of medicine and food homologous flowers on the prevention and treatment of related diseases. J. Food Biochem. 2022, 46, e14430. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.M.; Issaoui, M.; Chammem, N. Analysis of Main and Healthy Phenolic Compounds in Foods. J. AOAC Int. 2019, 102, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chen, M. Research progress on the prevention and treatment of cardiovascular diseases by the regulation of intestinal flora by phenolic acids of traditional Chinese medicine. Chin. Tradit. Pat. Med. 2022, 44, 3920–3926. [Google Scholar]
- Jiao, H.; Liu, H.; Xue, X.; Chen, X. Research Progress on Active Compounds and Biological Activities of Polyphenols from Blueberry Leaves. Guangzhou Chem. Ind. 2022, 50, 10–12+20. [Google Scholar]
- Soheila, J.; Maleki, J.F.C.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar]
- Ye, D. Preparation and Properties of Lignin-Based Polyphenolantioxidants; Guangdong University of Technology: Guangzhou, China, 2022. [Google Scholar]
- Li, Z.R.; Zhao, L.; Chen, L.; Yu, Y.; Wang, D.; Mao, X.; Cao, G.; Zhao, Z.; Yang, H. Unique roles in health promotion of dietary flavonoids through gut microbiota regulation: Current understanding and future perspectives. Food Chem. 2023, 399, 133959. [Google Scholar] [CrossRef]
- Singh, A.; Yau, Y.F.; Leung, K.S.; El-Nezami, H.; Lee, J.C. Interaction of Polyphenols as Antioxidant and Anti-Inflammatory Compounds in Brain-Liver-Gut Axis. Antioxidants 2020, 9, 669. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Cheung, F.; Hong, M.; Feng, Y. The Potential and Action Mechanism of Polyphenols in the Treatment of Liver Diseases. Oxid. Med. Cell Longev. 2018, 2018, 8394818. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Aquino, E.; Quezada-Ramirez, M.A.; Silva-Olivares, A.; Ramos-Tovar, E.; Flores-Beltran, R.E.; Segovia, J.; Shibayama, M.; Muriel, P. Curcumin downregulates Smad pathways and reduces hepatic stellate cells activation in experimental fibrosis. Ann. Hepatol. 2020, 19, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Lian, N.; Jin, H.; Zhang, F.; Wu, L.; Shao, J.; Lu, Y.; Zheng, S. Curcumin inhibits aerobic glycolysis in hepatic stellate cells associated with activation of adenosine monophosphate-activated protein kinase. IUBMB Life 2016, 68, 589–596. [Google Scholar] [CrossRef] [PubMed]
- MadanKumar, P.; NaveenKumar, P.; Manikandan, S.; Devaraj, H.; NiranjaliDevaraj, S. Morin ameliorates chemically induced liver fibrosis in vivo and inhibits stellate cell proliferation in vitro by suppressing Wnt/beta-catenin signaling. Toxicol. Appl. Pharmacol. 2014, 277, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Luo, L.; Zhu, Z.D.; Zhou, X.; Wang, Y.; Xue, J.; Zhang, J.; Cai, X.; Chen, Z.L.; Ma, Q.; et al. Chlorogenic Acid Inhibits Liver Fibrosis by Blocking the miR-21-Regulated TGF-beta1/Smad7 Signaling Pathway In Vitro and In Vivo. Front. Pharmacol. 2017, 8, 929. [Google Scholar] [CrossRef]
- Shi, H.; Dong, L.; Jiang, J.; Zhao, J.; Zhao, G.; Dang, X.; Lu, X.; Jia, M. Chlorogenic acid reduces liver inflammation and fibrosis through inhibition of toll-like receptor 4 signaling pathway. Toxicology 2013, 303, 107–114. [Google Scholar] [CrossRef]
- Qin Yang, T.T.; He, Q.; Guo, C.; Chen, D.; Tan, Y.; Feng, J.; Song, X.; Li, J. Combined Amphiphilic Silybin Meglumine Nanosuspension Effective Against Hepatic Fibrosis in Mice Model. Int. J. Nanomed. 2023, 18, 5197–5211. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, S.; Sun, Z.; Tong, Z.; Zhou, L.; Xu, Q.; Zhou, H. Preclinical validation of silibinin/albumin nanoparticles as an applicable system against acute liver injury. Acta Biomater. 2022, 146, 385–395. [Google Scholar] [CrossRef]
- Balta, C.; Herman, H.; Boldura, O.M.; Gasca, I.; Rosu, M.; Ardelean, A.; Hermenean, A. Chrysin attenuates liver fibrosis and hepatic stellate cell activation through TGF-beta/Smad signaling pathway. Chem. Biol. Interact. 2015, 240, 94–101. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Xu, W.; Wang, S.; Hu, Z.; Zhang, Q.; Deng, X.; Wang, J.; Zhang, J.; Guo, C. Antifibrotic effects of luteolin on hepatic stellate cells and liver fibrosis by targeting AKT/mTOR/p70S6K and TGFbeta/Smad signalling pathways. Liver Int. 2015, 35, 1222–1233. [Google Scholar] [CrossRef]
- Zhao, P.X.S.; Chaggan, C.; Liao, Z.; Wong, K.; He, F.; Singh, S.; Loomba, R.; Karin, M.; Witztum, J.L.; Saltiel, A.R. An AMPK–caspase-6 axis controls liver damage in nonalcoholic steatohepatitis. Science 2020, 367, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Virender Kumar, G.M.; Dutta, R.; Mahato, R.I. Co-delivery of small molecule hedgehog inhibitor and miRNA for treating liver fibrosis. Biomaterials 2015, 76, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xue, X.; Fan, G.; Gu, Y.; Zhou, F.; Zheng, Q.; Liu, R.; Li, Y.; Ma, B.; Li, S.; et al. Ferulic Acid Ameliorates Hepatic Inflammation and Fibrotic Liver Injury by Inhibiting PTP1B Activity and Subsequent Promoting AMPK Phosphorylation. Front. Pharmacol. 2021, 12, 754976. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Li, Z.; Yang, Z.; Wang, M.; Zhang, W.; Ren, Y.; Li, L.; Hu, J.; Sun, Z.; Nie, S. Ferulic acid positively modulates the inflammatory response to septic liver injury through the GSK-3beta/NF-kappaB/CREB pathway. Life Sci. 2021, 277, 119584. [Google Scholar] [CrossRef]
- Li, J.J.; Jiang, H.C.; Wang, A.; Bu, F.T.; Jia, P.C.; Zhu, S.; Zhu, L.; Huang, C.; Li, J. Hesperetin derivative-16 attenuates CCl(4)-induced inflammation and liver fibrosis by activating AMPK/SIRT3 pathway. Eur. J. Pharmacol. 2022, 915, 174530. [Google Scholar] [CrossRef]
- Zhou, B.; Yang, C.; Yan, X.; Shi, Z.; Xiao, H.; Wei, X.; Jiang, N.; Wu, Z. LETM1 Knockdown Promotes Autophagy and Apoptosis Through AMP-Activated Protein Kinase Phosphorylation-Mediated Beclin-1/Bcl-2 Complex Dissociation in Hepatocellular Carcinoma. Front. Oncol. 2020, 10, 606790. [Google Scholar] [CrossRef]
- Zhou, G.; Li, C.; Zhan, Y.; Zhang, R.; Lv, B.; Geng, W.; Zheng, J. Pinostilbene hydrate suppresses hepatic stellate cell activation via inhibition of miR-17-5p-mediated Wnt/beta-catenin pathway. Phytomedicine 2020, 79, 153321. [Google Scholar] [CrossRef]
- Lin, X.; Kong, L.N.; Huang, C.; Ma, T.T.; Meng, X.M.; He, Y.; Wang, Q.Q.; Li, J. Hesperetin derivative-7 inhibits PDGF-BB-induced hepatic stellate cell activation and proliferation by targeting Wnt/beta-catenin pathway. Int. Immunopharmacol. 2015, 25, 311–320. [Google Scholar] [CrossRef]
- Park, M.Y.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Abusaliya, A.; Jeong, S.H.; Park, J.S.; Heo, J.D.; Kim, G.S. Scutellarein Inhibits LPS-Induced Inflammation through NF-kappaB/MAPKs Signaling Pathway in RAW264.7 Cells. Molecules 2022, 27, 3782. [Google Scholar] [CrossRef]
- Ray, K. Activation of NF-κB signaling in hepatocytes induces liver fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 224. [Google Scholar] [CrossRef]
- Li, S.; Wang, R.; Song, F.; Chen, P.; Gu, Y.; Chen, C.; Yuan, Y. Salvianolic acid A suppresses CCl(4)-induced liver fibrosis through regulating the Nrf2/HO-1, NF-kappaB/IkappaBalpha, p38 MAPK, and JAK1/STAT3 signaling pathways. Drug Chem. Toxicol. 2022, 46, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Song, F.; Li, S.; Wu, B.; Gu, Y.; Yuan, Y. Salvianolic acid A attenuates CCl(4)-induced liver fibrosis by regulating the PI3K/AKT/mTOR, Bcl-2/Bax and caspase-3/cleaved caspase-3 signaling pathways. Drug Des. Devel. Ther. 2019, 13, 1889–1900. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, K.; Zhang, R.J.; Mei, D.; Zhang, B. Isochlorogenic Acid A Attenuates the Progression of Liver Fibrosis Through Regulating HMGB1/TLR4/NF-kappaB Signaling Pathway. Front. Pharmacol. 2020, 11, 582. [Google Scholar] [CrossRef] [PubMed]
- Dorn, C.; Kraus, B.; Motyl, M.; Weiss, T.S.; Gehrig, M.; Scholmerich, J.; Heilmann, J.; Hellerbrand, C. Xanthohumol, a chalcon derived from hops, inhibits hepatic inflammation and fibrosis. Mol. Nutr. Food Res. 2010, 54 (Suppl. S2), S205–S213. [Google Scholar] [CrossRef]
- El-Mihi, K.A.; Kenawy, H.I.; El-Karef, A.; Elsherbiny, N.M.; Eissa, L.A. Naringin attenuates thioacetamide-induced liver fibrosis in rats through modulation of the PI3K/Akt pathway. Life Sci. 2017, 187, 50–57. [Google Scholar] [CrossRef]
- Lee, K.W.; Thiyagarajan, V.; Sie, H.W.; Cheng, M.F.; Tsai, M.J.; Chia, Y.C.; Weng, C.F. Synergistic effect of natural compounds on the fatty acid-induced autophagy of activated hepatic stellate cells. J. Nutr. Biochem. 2014, 25, 903–913. [Google Scholar] [CrossRef]
- He, Y.J.; Kuchta, K.; Deng, Y.M.; Cameron, S.; Lin, Y.; Liu, X.Y.; Ye, G.R.; Lv, X.; Kobayashi, Y.; Shu, J.C. Curcumin Promotes Apoptosis of Activated Hepatic Stellate Cells by Inhibiting Protein Expression of the MyD88 Pathway. Planta Med. 2017, 83, 1392–1396. [Google Scholar] [CrossRef]
- Feng, J.; Wang, C.; Liu, T.; Li, J.; Wu, L.; Yu, Q.; Li, S.; Zhou, Y.; Zhang, J.; Chen, J.; et al. Procyanidin B2 inhibits the activation of hepatic stellate cells and angiogenesis via the Hedgehog pathway during liver fibrosis. J. Cell Mol. Med. 2019, 23, 6479–6493. [Google Scholar] [CrossRef]
- Zhu, S.; Chen, X.; Chen, S.Y.; Wang, A.; Wu, S.; Wu, Y.Y.; Cheng, M.; Xu, J.J.; Li, X.F.; Huang, C.; et al. Hesperetin derivative decreases CCl(4)-induced hepatic fibrosis by Ptch1-dependent mechanisms. J. Biochem. Mol. Toxicol. 2022, 36, e23149. [Google Scholar] [CrossRef]
- Tao, S.; Duan, R.; Xu, T.; Hong, J.; Gu, W.; Lin, A.; Lian, L.; Huang, H.; Lu, J.; Li, T. Salvianolic acid B inhibits the progression of liver fibrosis in rats via modulation of the Hedgehog signaling pathway. Exp. Ther. Med. 2022, 23, 116. [Google Scholar] [CrossRef]
- Wang, C.; Ma, C.; Fu, K.; Gong, L.H.; Zhang, Y.F.; Zhou, H.L.; Li, Y.X. Phillygenin Attenuates Carbon Tetrachloride-Induced Liver Fibrosis via Modulating Inflammation and Gut Microbiota. Front. Pharmacol. 2021, 12, 756924. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Asumda, F.Z.; Moser, C.D.; Kang, Y.N.N.; Lai, J.P.; Roberts, L.R. Sulfatase-2 Regulates Liver Fibrosis through the TGF-beta Signaling Pathway. Cancers 2021, 13, 5279. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. The effects and mechanism of polyphenols from Callistephus Chinensis Flower on Hepatic Fibrosis. Liaoning Univ. 2017, 44, 5. [Google Scholar]
- Gao, J.; Yang, J.; Yu, W.; Hao, R.; Fan, J.; Wei, J. Gooseberry anthocyanins protect mice hepatic fibrosis by inhibiting TGF-β/Smad pathway. Food Biosci. 2020, 37, 100717. [Google Scholar] [CrossRef]
- Bi, X.; Liu, X.; Sun, Y.; Liu, Y. Plant Polyphenols Reduce the Mechanisms and Research Progress of Liver Fibrosis. J. Liaoning Univ. (Nat. Sci. Ed.) 2017, 44, 257–261. [Google Scholar]
- James, J.; Tomasek, G.G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar]
- Shrestha, N.; Chand, L.; Han, M.K.; Lee, S.O.; Kim, C.Y.; Jeong, Y.J. Glutamine inhibits CCl4 induced liver fibrosis in mice and TGF-beta1 mediated epithelial-mesenchymal transition in mouse hepatocytes. Food Chem. Toxicol. 2016, 93, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.D.; Zhou, J.; Chen, E.Q. Molecular Mechanisms and Potential New Therapeutic Drugs for Liver Fibrosis. Front. Pharmacol. 2022, 13, 787748. [Google Scholar] [CrossRef]
- Kim, J.Y.; An, H.J.; Kim, W.H.; Gwon, M.G.; Gu, H.; Park, Y.Y.; Park, K.K. Anti-fibrotic Effects of Synthetic Oligodeoxynucleotide for TGF-beta1 and Smad in an Animal Model of Liver Cirrhosis. Mol. Ther. Nucleic Acids 2017, 8, 250–263. [Google Scholar] [CrossRef]
- Shu, G.; Dai, C.; Yusuf, A.; Sun, H.; Deng, X. Limonin relieves TGF-beta-induced hepatocyte EMT and hepatic stellate cell activation in vitro and CCl(4)-induced liver fibrosis in mice via upregulating Smad7 and subsequent suppression of TGF-beta/Smad cascade. J. Nutr. Biochem. 2022, 107, 109039. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad Signaling Pathways of the TGF-beta Family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef] [PubMed]
- Gart, E.; van Duyvenvoorde, W.; Toet, K.; Caspers, M.P.M.; Verschuren, L.; Nielsen, M.J.; Leeming, D.J.; Souto Lima, E.; Menke, A.; Hanemaaijer, R.; et al. Butyrate Protects against Diet-Induced NASH and Liver Fibrosis and Suppresses Specific Non-Canonical TGF-beta Signaling Pathways in Human Hepatic Stellate Cells. Biomedicines 2021, 9, 1954. [Google Scholar] [CrossRef] [PubMed]
- Tee, J.K.; Peng, F.; Tan, Y.L.; Yu, B.; Ho, H.K. Magnesium Isoglycyrrhizinate Ameliorates Fibrosis and Disrupts TGF-beta-Mediated SMAD Pathway in Activated Hepatic Stellate Cell Line LX2. Front. Pharmacol. 2018, 9, 1018. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zhou, D.; Meng, X.; Wang, X.; Liu, C.; Huang, C.; Li, J.; Zhang, L. Smad2 increases the apoptosis of activated human hepatic stellate cells induced by TRAIL. Int. Immunopharmacol. 2016, 32, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, Y.O.I. Emerging insights into Transforming growth factor beta Smad signal in hepatic fibrogenesis. Gut 2007, 56, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kong, D.; Qiu, J.; Xie, Y.; Lu, Z.; Zhou, C.; Liu, X.; Zhang, R.; Wang, Y. Praziquantel ameliorates CCl(4) -induced liver fibrosis in mice by inhibiting TGF-beta/Smad signalling via up-regulating Smad7 in hepatic stellate cells. Br. J. Pharmacol. 2019, 176, 4666–4680. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Li, R.; Cai, M.; Huang, J.; Huang, W.; Guo, Y.; Yang, L.; Yang, G.; Lan, T.; Zhu, K. Andrographolide Ameliorates Liver Fibrosis in Mice: Involvement of TLR4/NF-kappaB and TGF-beta1/Smad2 Signaling Pathways. Oxid. Med. Cell Longev. 2018, 2018, 7808656. [Google Scholar] [CrossRef]
- Yang, P.C.; Bai, W.Z.; Wang, J.; Yan, C.H.; Huang, W.F.; Jiang, S.Z. Sedum sarmentosum Total Flavonoids Alleviate Schistosomiasis-Induced Liver Fibrosis by Altering TGF-beta1 and Smad7 Expression. Evid. Based Complement Altern. Med. 2020, 2020, 2083697. [Google Scholar] [CrossRef]
- Sascha Behne, H.F.; Schwarz, S.; Lachenmeier, D.W. Risk Assessment of Chlorogenic and Isochlorogenic Acids in Coffee By-Products. Molecules 2023, 28, 5540. [Google Scholar] [CrossRef]
- Marco Trappoliere, A.C.; Schmid, M.; Bertolani, C.; Failli, P.; Vizzutti, F.; Novo, E.; di Manzano, C.; Marra, F.; Loguercio, C.; Pinzani, M. Silybin, a component of sylimarin, exerts anti-inflammatory and anti-fibrogenic effects on human hepatic stellate cells. J. Hepatol. 2009, 50, 1102–1111. [Google Scholar] [CrossRef]
- Meltem Macit, G.D.; Cumbul, A.; Sumer, E.; Macit, C. Formulation development of Silybum marianum seed extracts and silymarin nanoparticles, and evaluation of hepatoprotective effect. J. Drug Deliv. Sci. Technol. 2023, 83, 104378. [Google Scholar] [CrossRef]
- Bonepally, C.R.; Gandey, S.J.; Bommineni, K.; Gottumukkala, K.M.; Aukunuru, J. Preparation, Characterisation and In Vivo Evaluation of Silybin Nanoparticles for the Treatment of Liver Fibrosis. Trop. J. Pharm. Res. 2013, 12, 1–6. [Google Scholar] [CrossRef]
- Luo, S.Y.Y.; Zhao, T.; Zhang, R.; Fang, C.; Li, Y.; Zhang, Z.; Gong, T. Albumin-Based Silibinin Nanocrystals Targeting Activated Hepatic Stellate Cells for Liver Fibrosis Therapy. ACS Appl. Mater. Interfaces 2023, 15, 7747–7758. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.L.; Wang, L.; Zhang, K.; Peng, J.; Fan, G. Therapeutic Effect of Curcumin on Metabolic Diseases: Evidence from Clinical Studies. Int. J. Mol. Sci. 2023, 24, 3323. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-N.; Hsu, S.L.; Liao, M.-Y.; Liu, Y.-T.; Lai, C.-H.; Chen, J.-F.; Nguyen, M.-H.; Su, Y.-H.; Chen, S.-T.; Wu, L.-C. Ameliorative Effect of Curcumin-Encapsulated Hyaluronic Acid–PLA Nanoparticles on Thioacetamide-Induced Murine Hepatic Fibrosis. Int. J. Environ. Res. Public Health 2016, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pan, W.; Wang, Y.; Lei, W.; Feng, B.; Du, C.; Wang, X. Enhanced efficacy of curcumin with phosphatidylserine-decorated nanoparticles in the treatment of hepatic fibrosis. Drug Deliv. 2017, 25, 1–11. [Google Scholar] [CrossRef]
- Elzoheiry, A.; Ayad, E.; Omar, N.; Elbakry, K.; Hyder, A. Anti-liver fbrosis activity of curcumin/chitosan-coated green silver nanoparticles. Sci. Rep. 2022, 12, 18403. [Google Scholar] [CrossRef]
- Faiyaz Shakeel, M.M.A.; Alam, P.; Alshetaili, A.; Haq, N.; Alanazi, F.K.; Alshehri, S.; Ghoneim, M.M.; Alsarra, I.A. Hepatoprotective Effects of Bioflavonoid Luteolin Using Self-Nanoemulsifying Drug Delivery System. Molecules 2021, 26, 7497. [Google Scholar] [CrossRef]
- Renuka Mani, V.N. Chrysin: Sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef]
- Liu, K.; Gong, B.; He, F.; Li, S.; Zheng, Y.; Lei, H. Effects of Luteolin on Inflammation via TLR4/NF-κB Pathway in Young Rats in an Asthma Model. Heilongjiang Med. J. 2023, 47, 133–136. [Google Scholar]
- Zheng, Y.; Zhang, Y.; Zhu, X.; Zhou, X.; Yi, L. Research progress on role of AMPK in pathogenesis of liver fibrosis. Occup. Health 2019, 35, 62780–62792. [Google Scholar]
- Kumar, P.; Smith, T.; Rahman, K.; Thorn, N.E.; Anania, F.A. Adiponectin agonist ADP355 attenuates CCl4-induced liver fibrosis in mice. PLoS ONE 2014, 9, e110405. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.T.; Jiang, S.; Xu, J.; Di, W.; Yang, Z.; Hu, W.; Yang, Y. AMPK: A novel target for treating hepatic fibrosis. Int. J. Mol. Sci. 2023, 24, 3323. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.P.-F.; Kim, K.; Zhu, C.; Yu, J.; Meroni, M.; Bartolomé, A.; Dapito, D.H.; Ferrante, A.W., Jr.; Dongiovanni, P.; Valenti, L.; et al. Notch-mediated hepatocyte MCP-1 secretion causes liver fibrosis. JCI Insight 2023, 8, e165369. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Guo, F.; Li, Y.; Liang, Y.; Li, G.; Fu, P.; Ma, L. Activation of AMPK by triptolide alleviates nonalcoholic fatty liver disease by improving hepatic lipid metabolism, inflammation and fibrosis. Phytomedicine 2021, 92, 153739. [Google Scholar] [CrossRef] [PubMed]
- Shui, L.; Wang, W.; Xie, M.; Ye, B.; Li, X.; Liu, Y.; Zheng, M. Isoquercitrin induces apoptosis and autophagy in hepatocellular carcinoma cells via AMPK/mTOR/p70S6K signaling pathway. Aging-US 2020, 12, 24318. [Google Scholar] [CrossRef] [PubMed]
- Strzyz, P. AMPK against NASH. Nat. Rev. Mol. Cell Biol. 2020, 21, 181. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Wang, D.; Xu, Y.; Dong, R.; Yang, Y.; Lv, Q.; Chen, X.; Zhang, Z. The Upstream Pathway of mTOR-Mediated Autophagy in Liver Diseases. Cells 2019, 8, 1597. [Google Scholar] [CrossRef]
- Shu, S.P. Combined Regulation of AMPK-mTOR Pathway by Taurine, EGCG and Genistein on the Inhibition of Hepatic Stellate Cell Autophagy against Hepatic Fibrosis in Rats; Guangxi Medical University: Nanning, China, 2019. [Google Scholar]
- Juhaer Anjum, S.M.; Das, R.; Alam, R.; Mojumder, A.; Emran, T.B.; Islam, F.; Rauf, A.; Hossain, M.J.; Aljohani, A.S.M.; Al Abdulmonem, W.; et al. A renewed concept on the MAPK signaling pathway in cancers: Polyphenols as a choice of therapeutics. Pharmacol. Res. 2022, 184, 106398. [Google Scholar] [CrossRef]
- de Oliveira Silva, E.; Batista, R. Ferulic Acid and Naturally Occurring Compounds Bearing a Feruloyl Moiety: A Review on Their Structures, Occurrence, and Potential Health Benefits. Compr. Rev. Food Sci. Food Saf. 2017, 16, 580–616. [Google Scholar] [CrossRef]
- Tabassum Ara, S.O.; Hasan, M.; Ozono, M.; Kogure, K. Protective effects of liposomes encapsulating ferulic acid against CCl4-induced oxidative liver damage in vivo rat model. J. Clin. Biochem. Nutr. 2022, 72, 46–53. [Google Scholar]
- Yang, K.N.W.; Münch, C.; Xu, Y.; Satterstrom, K.; Jeong, S.; Hayes, S.D.; Jedrychowski, M.P.; Vyas, F.S.; Zaganjor, E.; Guarani, V.; et al. Mitochondrial Sirtuin Network Reveals Dynamic SIRT3-Dependent Deacetylation in Response to Membrane Depolarization. Cell 2016, 167, 985–1000.e21. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Z.; Wang, Y.; Sun, R.; Zou, B.; Tian, X.; Liu, D.; Zhao, X.; Zhou, J.; Zhao, Y.; et al. SIRT3 regulates mitophagy in liver fibrosis through deacetylation of PINK1/NIPSNAP1. J. Cell. Physiol. 2023, 238, 2090–2102. [Google Scholar] [CrossRef] [PubMed]
- Mas-Bargues, C.B.; Vina, J. Genistein, a tool for geroscience. Mech. Ageing Dev. 2022, 204, 111665. [Google Scholar] [CrossRef]
- Xiang, T.; Zhang, S.; Cheng, N.; Ge, S.; Wen, J.; Xiao, J.; Wu, X. Oxidored-nitro domain-containing protein 1 promotes liver fibrosis by activating the Wnt/beta-catenin signaling pathway in vitro. Mol. Med. Rep. 2017, 16, 5050–5054. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Sun, Z.; Zhang, L.; Xu, H. SERP1 reduces inchoate acute hepatic injury through regulation of endoplasmic reticulum stress via the GSK3β/β-catenin/TCF/LEF signaling pathway. Mol. Med. Rep. 2022, 25, 193. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, T.; Su, Z.; Pi, C.; Wei, Y.; Zhao, L. Latest research progress on anticancer effect of baicalin and its aglycone baicalein. Arch. Pharm. Res. 2022, 45, 535–557. [Google Scholar] [CrossRef]
- Risha Ganguly, A.G.; Pandey, A.K. Role of baicalin as a potential therapeutic agent in hepatobiliary and gastrointestinal disorders: A review. World J. Gastroenterol. 2022, 28, 3047. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, Y.; Gong, X.; Zhang, L.; Zhou, Q.; Wu, S.; Hu, J.; Kuang, G.; Ying, X.; Wan, J.; et al. Baicalin and its nanoliposomes ameliorates nonalcoholic fatty liver disease via suppression of TLR4 signaling cascade in mice. Int. Immunopharmacol. 2020, 80, 106208. [Google Scholar] [CrossRef] [PubMed]
- Perugorria, M.J.; Olaizola, P.; Labiano, I.; Esparza-Baquer, A.; Marzioni, M.; Marin, J.J.G.; Bujanda, L.; Banales, J.M. Wnt-beta-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 121–136. [Google Scholar] [CrossRef]
- Zhang, X.; Zeng, Y.; Zhao, L.; Xu, Q.; Miao, D.; Yu, F. Targeting Hepatic Stellate Cell Death to Reverse Hepatic Fibrosis. Curr. Drug Targets 2023, 24, 568–583. [Google Scholar] [CrossRef]
- Eleanor Jayawant, A.P.; Clark, H.; Kennedy, E.; Ghodke, A.; Jones, J.; Pepper, C.; Pepper, A.; Mitchell, S. NF-κB fingerprinting reveals heterogeneous NF-κB composition in diffuse large B-cell lymphoma. Front. Oncol. 2023, 13, 1181660. [Google Scholar] [CrossRef]
- Matthew Biancalana, E.N.; Lenardo, M.J.; Fersht, A.R. NF-κB Rel subunit exchange on a physiological timescale. Protein Sci. 2021, 30, 1818–1832. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Wang, Y.; Peng, R.; Wu, Y.; Yuan, Y. Ginkgo biloba extract mitigates liver fibrosis and apoptosis by regulating p38 MAPK, NF-kappaB/IkappaBalpha, and Bcl-2/Bax signaling. Drug Des. Devel. Ther. 2015, 9, 6303–6317. [Google Scholar]
- Liu, M.; Wu, Q.; Chen, P.; Buchele, B.; Bian, M.; Dong, S.; Huang, D.; Ren, C.; Zhang, Y.; Hou, X.; et al. A boswellic acid-containing extract ameliorates schistosomiasis liver granuloma and fibrosis through regulating NF-kappaB signaling in mice. PLoS ONE 2014, 9, e100129. [Google Scholar]
- Lin, X.; Wei, Y.; Li, Y.; Xiong, Y.; Fang, B.; Li, C.; Huang, Q.; Huang, R.; Wei, J. Tormentic Acid Ameliorates Hepatic Fibrosis in vivo by Inhibiting Glycerophospholipids Metabolism and PI3K/Akt/mTOR and NF-kappaB Pathways: Based on Transcriptomics and Metabolomics. Front. Pharmacol. 2022, 13, 801982. [Google Scholar] [CrossRef]
- de Souza Basso, B.; Haute, G.V.; Ortega-Ribera, M.; Luft, C.; Antunes, G.L.; Bastos, M.S.; Carlessi, L.P.; Levorse, V.G.; Cassel, E.; Donadio, M.V.F.; et al. Methoxyeugenol deactivates hepatic stellate cells and attenuates liver fibrosis and inflammation through a PPAR-ɣ and NF-kB mechanism. J. Ethnopharmacol. 2021, 280, 114433. [Google Scholar] [CrossRef]
- Wang, B.; Yang, H.; Fan, Y.; Yang, Y.; Cao, W.; Jia, Y.; Cao, Y.; Sun, K.; Pang, Z.; Du, H. 3-Methyladenine ameliorates liver fibrosis through autophagy regulated by the NF-κB signaling pathways on hepatic stellate cell. Oncotarget 2017, 8, 107603. [Google Scholar] [CrossRef]
- Senad Medunjanin, L.S.; Fiegehenn, C.; SoenkeWeinert; Braun-Dullaeus, R. GSK-3β controls NF-kappaB activity via IKKγ/NEMO. Sci. Rep. 2016, 6, 38553. [Google Scholar] [CrossRef]
- Bai, F.; Huang, Q.; Nie, J.; Lu, S.; Lu, C.; Zhu, X.; Wang, Y.; Zhuo, L.; Lu, Z.; Lin, X. Trolline Ameliorates Liver Fibrosis by Inhibiting the NF-kappaB Pathway, Promoting HSC Apoptosis and Suppressing Autophagy. Cell Physiol. Biochem. 2017, 44, 436–446. [Google Scholar] [CrossRef]
- Tran, C.W.; Saibil, S.D.; Le Bihan, T.; Hamilton, S.R.; Lang, K.S.; You, H.; Lin, A.E.; Garza, K.M.; Elford, A.R.; Tai, K.; et al. Glycogen Synthase Kinase-3 Modulates Cbl-b and Constrains T Cell Activation. J. Immunol. 2017, 199, 4056–4065. [Google Scholar] [CrossRef]
- Francisca Echeverría, A.B.; Sambra, V.; Álvarez, D.; Videla, L.; Valenzuela, R. Beneficial Effects of Dietary Polyphenols in the Prevention and Treatment of NAFLD: Cell-Signaling Pathways Underlying Health Effects. Curr. Med. Chem. 2022, 29, 299–328. [Google Scholar] [CrossRef]
- Piotr Holowinski, A.L.D.; Typek, R. Chlorogenic acid-water complexes in chlorogenic acid containing food products. J. Food Compos. Anal. 2022, 109, 104509. [Google Scholar] [CrossRef]
- Éliás, A.J.; Balogh, L.; Brányik, T.; Mák, E.; Csobod, É.C.; Bálint, M.V.; Benedek, C. Xanthohumol-Enriched Beer Does Not Exert Antitumorigenic Effects on HeLa Cell Line In Vivo. Molecules 2023, 28, 1070. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Wei, L.; Chen, Q.; Guo, A.; Fan, J.; Wang, R.; Zhang, H. Asiatic acid attenuates CCl(4)-induced liver fibrosis in rats by regulating the PI3K/AKT/mTOR and Bcl-2/Bax signaling pathways. Int. Immunopharmacol. 2018, 60, 1–8. [Google Scholar] [CrossRef]
- Son, M.K.; Ryu, Y.L.; Jung, K.H.; Lee, H.; Lee, H.S.; Yan, H.H.; Park, H.J.; Ryu, J.K.; Suh, J.K.; Hong, S.; et al. HS-173, a novel PI3K inhibitor, attenuates the activation of hepatic stellate cells in liver fibrosis. Sci. Rep. 2013, 3, 3470. [Google Scholar] [CrossRef]
- Dong, Z.; Li, S.; Wang, X.; Si, L.; Ma, R.; Bao, L.; Bo, A. lncRNA GAS5 restrains CCl(4)-induced hepatic fibrosis by targeting miR-23a through the PTEN/PI3K/Akt signaling pathway. Am. J. Physiol. Gastrointest Liver Physiol. 2019, 316, G539–G550. [Google Scholar] [CrossRef]
- Erwin Gäbele, S.R.; Tsukada, S.; Bataller, R.; Yata, Y.; Morris, T.; Schrum, L.W.; Brenner, D.A.; Rippe, R.A. The role of p70S6K in hepatic stellate cell collagen gene expression and cell proliferation. J. Biol. Chem. 2005, 280, 13374–13382. [Google Scholar] [CrossRef]
- Huang, J.; Chen, L.; Wu, J.; Ai, D.; Zhang, J.-Q.; Chen, T.-G.; Wang, L. Targeting the PI3K/AKT/mTOR Signaling Pathway in the Treatment of Human Diseases: Current Status, Trends, and Solutions. J. Med. Chem. 2022, 65, 16033–16061. [Google Scholar] [CrossRef]
- Lin, X.; Li, Y.; Zhang, X.; Wei, Y.; Wen, S.; Lu, Z.; Huang, Q.; Wei, J. Tormentic acid inhibits hepatic stellate cells activation via blocking PI3K/Akt/mTOR and NF-kappaB signalling pathways. Cell Biochem. Funct. 2021, 39, 77–87. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.; Zhang, J.; Xiong, C.; Huang, G. The mechanism of delaying starch digestion by luteolin. Food Funct. 2021, 12, 11862–11871. [Google Scholar] [CrossRef]
- Huang, Y.; Luo, W.; Chen, S.; Su, H.; Zhu, W.; Wei, Y.; Qiu, Y.; Long, Y.; Shi, Y.; Wei, J. Isovitexin alleviates hepatic fibrosis by regulating miR-21-mediated PI3K/Akt signaling and glutathione metabolic pathway: Based on transcriptomics and metabolomics. Phytomedicine 2023, 121, 155117. [Google Scholar] [CrossRef]
- Gu, Y.; Liu, X.; Liao, L.; Gao, Y.; Shi, Y.; Ni, J.; He, G. Relationship between lipid metabolism and Hedgehog signaling pathway. J. Steroid. Biochem. Mol. Biol. 2021, 209, 105825. [Google Scholar] [CrossRef]
- Seki, E. HEDGEHOG Signal in hepatocytes mediates macrophage recruitment: A new mechanism and potential therapeutic target for fatty liver disease. Hepatology 2016, 63, 1071–1073. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Z.; Zhang, P.; Yu, M.; Yang, T. Role of canonical Hedgehog signaling pathway in liver. Int. J. Biol. Sci. 2018, 14, 1636–1644. [Google Scholar] [CrossRef]
- Yang, J.J.; Tao, H.; Li, J. Hedgehog signaling pathway as key player in liver fibrosis: New insights and perspectives. Expert Opin. Ther. Targets 2014, 18, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yan, J.; Wei, X.; Liu, Z.; Ni, J.; Hu, B.; Jin, L. Gant61 ameliorates CCl(4)-induced liver fibrosis by inhibition of Hedgehog signaling activity. Toxicol. Appl. Pharmacol. 2020, 387, 114853. [Google Scholar]
- Shen, X.; Peng, Y.; Li, H. The Injury-Related Activation of Hedgehog Signaling Pathway Modulates the Repair-Associated Inflammation in Liver Fibrosis. Front. Immunol. 2017, 8, 1450. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Omenetti, A.; Syn, W.K.; Diehl, A.M. The role of Hedgehog signaling in fibrogenic liver repair. Int. J. Biochem. Cell Biol. 2011, 43, 238–244. [Google Scholar] [CrossRef]
- Paola Infante, R.A.; Botta, B.; Mori, M.; Di Marcotullio, L. Targeting GLI factors to inhibit the Hedgehog pathway. Trends Pharmacol. Sci. 2015, 36, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Aslam, A.; Sheikh, N.; Shahzad, M.; Saeed, G.; Fatima, N.; Akhtar, T. Quercetin ameliorates thioacetamide-induced hepatic fibrosis and oxidative stress by antagonizing the Hedgehog signaling pathway. J. Cell Biochem. 2022, 123, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, D.; Guo, Q.; Shan, W.; Yang, J.; Lin, T.; Ye, S.; Zhou, X.; Ge, Y.; Bi, S.; et al. Theranostic Quercetin Nanoparticle for Treatment of Hepatic Fibrosis. Bioconjugate Chem. 2019, 29, 2939–2946. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Lu, Z.; Chen, B.; Wu, X.; Dong, P.; Zheng, J. Salvianolic acid B-induced microRNA-152 inhibits liver fibrosis by attenuating DNMT1-mediated Patched1 methylation. J. Cell Mol. Med. 2015, 19, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhang, J.; Chen, F.; Guo, L.; Zhu, Z.; Shi, J. An anti-ROS/hepatic fibrosis drug delivery system based on salvianolic acid B loaded mesoporous silica nanoparticles. Biomaterials 2010, 31, 7785–7796. [Google Scholar] [CrossRef] [PubMed]
- Guo, J. Protective Effects and Mechanisms of Phillygenin on Lipopolysaccharide-Induced Acute Lung Injury; Shanxi University: Taiyuan, China, 2022. [Google Scholar]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta -Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.S.; Harrison, D.J.; Kisielewski, D.; Cassidy, D.M.; McNeilly, A.D.; Gallagher, J.R.; Walsh, S.V.; Honda, T.; McCrimmon, R.J.; Dinkova-Kostova, A.T.; et al. Experimental Nonalcoholic Steatohepatitis and Liver Fibrosis Are Ameliorated by Pharmacologic Activation of Nrf2 (NF-E2 p45-Related Factor 2). Cell. Mol. Gastroenterol. Hepatol. 2017, 5, 367–398. [Google Scholar] [CrossRef]
- Sang, L.; Wang, X.-M.; Xu, D.-Y.; Sang, L.-X.; Han, Y.; Jiang, L.-Y. Morin enhances hepatic Nrf2 expression in a liver fibrosis rat model. World J. Gastroenterol. 2017, 23, 8334. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, F.; Thakur, K.; Wang, J.; Wang, H.; Hu, F.; Zhang, J.-G.; Wei, Z.-J. Molecular mechanism of anti-cancerous potential of Morin extracted from mulberry in Hela cells. Food Chem. Toxicol. 2018, 112, 466–475. [Google Scholar] [CrossRef]
- Tang, W.; Jiang, Y.F.; Ponnusamy, M.; Diallo, M. Murugavel Ponnusamy, Mamadou Diallo, Role of Nrf2 in chronic liver disease. World J. Gastroenterol. 2014, 20, 13079. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, N.; Kawada, N. Role of the Gut-Liver Axis in Liver Inflammation, Fibrosis, and Cancer: A Special Focus on the Gut Microbiota Relationship. Hepatol. Commun. 2019, 3, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Fan, X.; Schnabl, B. Role of the intestinal microbiome in liver fibrosis development and new treatment strategies. Transl. Res. 2019, 209, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Li, Y.; Zhou, F.; Guo, J.; Wang, T.; Shi, Y.; Yang, Y.; Lu, J.; Pei, G. Administration of a mixture of triterpenoids from yeyachun and phenolic acids from danshen ameliorates carbon tetrachloride-induced liver fibrosis in mice by the regulation of intestinal flora. J. Pharmacol. Sci. 2020, 143, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Song, H.; Zhang, X.; Song, G.; Wang, Y.; Ding, X.; Duan, X.; Li, L.; Sun, T.; Kan, Q. Metformin attenuated sepsis-related liver injury by modulating gut microbiota. Emerg. Microbes Infect. 2022, 11, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Li, Z.J.; Gou, H.Z.; Song, X.J.; Zhang, L. The gut microbiota-bile acid axis: A potential therapeutic target for liver fibrosis. Front. Cell Infect. Microbiol. 2022, 12, 945368. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, R.; Ahmed, W.; Afify, N.; Memon, M.; Yasin, M.; Memon, H.; Rustom, M.; Al Akeel, M.; Alhajri, N. Trust Your Gut: The Association of Gut Microbiota and Liver Disease. Microorganisms 2022, 10, 1045. [Google Scholar] [CrossRef]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The Severity of Nonalcoholic Fatty Liver Disease Is Associated With Gut Dysbiosis and Shift in the Metabolic Function of the Gut Microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef]
- Cheng, Z.; Wang, Y.; Li, B. Dietary Polyphenols Alleviate Autoimmune Liver Disease by Mediating the Intestinal Microenvironment: Challenges and Hopes. J. Agric. Food Chem. 2022, 70, 10708–10737. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, M.; Wang, Z.; Qin, F.; Chen, J.; He, Z. Dietary Polyphenols to Combat Nonalcoholic Fatty Liver Disease via the Gut-Brain-Liver Axis: A Review of Possible Mechanisms. J. Agric. Food Chem. 2021, 69, 3585–3600. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Santangelo, C.; Silenzi, A.; Scazzocchio, B.; Vari, R.; Masella, R. Dietary EVOO Polyphenols and Gut Microbiota Interaction: Are There Any Sex/Gender Influences? Antioxidants 2022, 11, 1744. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ma, C.; Fu, K.; Liu, Y.; Gong, L.; Peng, C.; Li, Y. Hepatoprotective effect of phillygenin on carbon tetrachloride-induced liver fibrosis and its effects on short chain fatty acid and bile acid metabolism. J. Ethnopharmacol. 2022, 296, 115478. [Google Scholar] [CrossRef] [PubMed]
- Baky, M.H.; Elshahed, M.; Wessjohann, L.; Farag, M.A. Interactions between dietary flavonoids and the gut microbiome: A comprehensive review. Br. J. Nutr. 2022, 128, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, R.; Wu, Y.; Wu, C.; Jia, X.; Dong, L.; Liu, L.; Chen, Y.; Bai, Y.; Zhang, M. Rice Bran Phenolic Extract Protects against Alcoholic Liver Injury in Mice by Alleviating Intestinal Microbiota Dysbiosis, Barrier Dysfunction, and Liver Inflammation Mediated by the Endotoxin-TLR4-NF-kappaB Pathway. J. Agric. Food Chem. 2020, 68, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A. I7 Interaction of polyphenols with gut microbiota; understanding the health effects of polyphenols. Biochem. Pharmacol. 2017, 139, 109. [Google Scholar] [CrossRef]
- Seki, E.; De Minicis, S.; Österreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. TLR4 enhances TGF-beta signaling and hepatic fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cavallaro, P.M.; Kim, B.M.; Liu, T.; Wang, H.; Kuhn, F.; Adiliaghdam, F.; Liu, E.; Vasan, R.; Samarbafzadeh, E.; et al. A role for intestinal alkaline phosphatase in preventing liver fibrosis. Theranostics 2021, 11, 14–26. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, M.; Jiang, X.; Qiao, Q.; Liu, T.; Zhao, C.; Wang, M. Comprehensive Analysis of Fecal Microbiome and Metabolomics in Hepatic Fibrosis Rats Reveal Hepatoprotective Effects of Yinchen Wuling Powder From the Host-Microbial Metabolic Axis. Front. Pharmacol. 2021, 12, 713197. [Google Scholar] [CrossRef]
- Guo, W.; Xiang, Q.; Mao, B.; Tang, X.; Cui, S.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Protective Effects of Microbiome-Derived Inosine on Lipopolysaccharide-Induced Acute Liver Damage and Inflammation in Mice via Mediating the TLR4/NF-kappaB Pathway. J. Agric. Food Chem. 2021, 69, 7619–7628. [Google Scholar] [CrossRef]
- Li, Z.; Ni, M.; Yu, H.; Wang, L.; Zhou, X.; Chen, T.; Liu, G.; Gao, Y. Gut Microbiota and Liver Fibrosis: One Potential Biomarker for Predicting Liver Fibrosis. Biomed. Res. Int. 2020, 2020, 3905130. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Ge, T.; Tang, C.; Wang, G.; Pang, L.; Chen, Z. Synergistic anti-inflammatory effect of gut microbiota and lithocholic acid on liver fibrosis. Inflamm. Res. 2022, 71, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, P.; Mohammed, S.A.D.; Li, Z.; Jiang, X.; Wu, J.; Liu, S. Exploration of the potential mechanism of Baicalin for hepatic fibrosis based on network pharmacology, gut microbiota, and experimental validation. Front. Microbiol. 2022, 13, 1051100. [Google Scholar] [CrossRef]
- Zhu, H.; Jiang, W.; Liu, C.; Wang, C.; Hu, B.; Guo, Y.; Cheng, Y.; Qian, H. Ameliorative effects of chlorogenic acid on alcoholic liver injury in mice via gut microbiota informatics. Eur. J. Pharmacol. 2022, 928, 175096. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenols/Polyphenol-Rich Plants | Categories/ Identified Polyphenols | Examples of Dietary Sources | Experimental Models | Dosage | Duration | Effects | References |
|---|---|---|---|---|---|---|---|
| Forsythiaside A | Phenolic acid | Forsythiae Fructus | CCl4-induced mouse | 15, 30, 60 mg/kg | 4 weeks | claudin-1↑, ZO-1↑, inclusion↑, LPS↓, MIP-1↓ | [22] |
| Callistephus Chinensis flower | Polyphenol | Rat model (CCl4-induced) | 50, 100 mg/L | 6 weeks | TGF-β1↓, Smad2↓, P-ERK1↓, P-NK1↓ | [26] | |
| Curcumin | Polyphenol | Roots of curcumin spp. | Rat model (CCl4-induced) | 100 mg/kg | 4 weeks | JNK↓, Smad3↓, Smad7↑ | [33] |
| Primary rat HSCs | 20 µM | 24 h | α-SMA↓, Col1α1↓ PPAR↑, AMPK↑ | [34] | |||
| HSCs | 25 µM | 24 h | TLRs↓, MyD88↓, NF-κB↓, TNF-α↓, IL-1β↓ | [35] | |||
| Chlorogenic acid | Phenolic acids | Coffee beans, honeysuckle, tobacco leaves and kiwi | hepatic stellate LX2 cell line | 20, 40, 80 µg/mL | 24 h | miR-21↓, α-SMA↓, TIMP-1↓, Smad7↑, MMP-9↑ | [36] |
| Rat model (CCl4-induced) | 15, 30, 60 mg/kg | 4 weeks | miR-21↓, α-SMA↓, TIMP-1↓, TGF-β1↓ Smad7↑, MMP-9↑ | ||||
| Rat model (CCl4-induced) | 60 mg/kg | 8 weeks | TLR4↓, MyD88↓, NF-κB↓, p-IκBα↓ Bambi↑, IκB α↑ | [37] | |||
| Silymarin | Flavonoid | Silybum marianum | CCl4- or BDL-induced fibrosis | TGF-β↓, α-SMA↓, collagen I↓ | [38] | ||
| LPS/D-GalN induced liver injury | (Nrf2)/antioxidant responsive element (ARE) pathway↑ caspase 9/3 related apoptosis pathway↓ | [39] | |||||
| Chrysin | Flavonoids | Propolis, blue passion flower (Passiflora caerulea), and honey | Rat model (CCl4-induced) | 50, 100 and 200 mg/kg | 2 weeks | α-SMA↓, TGF-β1↓, Smad 2/3↓ | [40] |
| Luteolin | Flavonoids | Pepper, chrysanthemum, Lonicerae japonicae flos | rat models CCl4, dimethylnitrosamine (DMN) and bile duct ligation (BDL) | p-AKT↓, p-Smad2↓ | [41] | ||
| Primary HSCs and HSC-T6 cells | TGF-β1 (2 ng/mL) | 2 h | a-SMA↓, collagen I/III↓, AKT↓, Smad2/3↓, TGF-β1↓ | ||||
| Baicalin | Flavonoids | Roots of Scutellaria baicalensis | BDL-induced | 67.5–270 μM | PPAR-γ↓, Wnt↓ | [42,43] | |
| 25–100 mg/kg | SCFA↑, regulating FXR and TGR5 receptor↑, PI3K↓, AKT↓, mTOR↓, IL-17↓ | ||||||
| Ferulic acid | Phenolic acid | Tomatoes, carrots, oranges, and corn | RAW 264.7 cells and LX-2 cells | 50, 100, 200 μM And 12.5, 25, 50 μM | 24 h | Acta2↓, Col1a1↓, p-Smad↓, p-Smad3↓, p-AMPK↑ | [44] |
| (mice) CCl4-induced | 25, 50 and 100 mg/kg | ALT↓, AST↓, TGF-1β↓, Acta2↓, NOX2↓, SOD↑, AMPK↑, ERK1/2↑ | |||||
| lipopolysaccharide (LPS)-induced cellular ALI models | 6, 12 mg/kg | 6 day | GSK-3β↑, CREB (Ser133)↑, IL-10↑, p-NF-κB↓, IL-1β↓, IL-6↓, IL-12↓, TNF-α↓ | [45] | |||
| Hesperetin derivative-16 (HD-16) | Flavonoids | Pericarp of citrus | LX-2 cells (human immortalized HSCs) | 4, 8, and 16 μM | α-SMA↓, Col1α1↓, Col3α1↓, TIMP-1↓, TNF-α↓, IL-1β↓, IL-10↑, IL-13↑, SIRT3↑ | [46] | |
| CCl4-induced mouse | 25 mg/kg, 50 mg/kg, 100 mg/kg | ALT↓, AST↓, ALP↓, α-SMA↓, Col1α1↓, TNF-α↓, IL-1β↓, IL-10↑, IL-13↑, SIRT3↑ | |||||
| Salvianolic acid A | Phenolic acid | Salvia miltiorrhiza | CCl4-induced rats | 5, 15 mg/kg | 6 weeks | p-AKT↓, p-mTOR↓, p-p70S6K1↓, caspase 3↓, Bax↓, α-SMA↓, PDGF-β↓, Desmin↓, Vimentin↓, TGF-β1↓, Bcl-2↑ | [47] |
| Pinostilbene hydrate | Phenolic acid | Primary HSCs | 20 μM, 80 μM | 48 h | Wnt/β-catenin↓, WIF1↑, GSK3β↑, APCP↑, β-catenin↑ | [48] | |
| Hesperetin derivative-7 | Flavonoids | CCl4-induced mouse | 50, 100, 200 mg/kg | 4 weeks | SMA↓, collagen I↓, β-catenin↓, c-myc↓, cyclind1↓, | [49] | |
| HSC-T6 cell line | 12.5, 25, 50, 100, 200 μM | 48 h | p-smad3↓, smad4↓ α-SMA↓, β-catenin↓, cyclind1↓, c-myc↓ | ||||
| Morin | Flavonoids | Mulberry leaves | diethylnitrosamine induced rat model of liver fibrosis | 50 mg/kg | 6 weeks | GSK-3β↓, β-catenin↓, cyclin D1↓, c-myc↓ | [35,50] |
| CCl4-induced mouse | 50 mg/kg | 8 weeks | Nrf2↑, NQO1↑, HO-1↑ | ||||
| LX-2 cells (culture-activated human hepatic stellate cells) | 50 μM | 24, 48 h | GFAP↓, Wnt5a/b↓, Wnt3↓, GSK-3β↓, β-catenin↓ | ||||
| Quercetin | Flavonoid | Fruits and vegetables | Thioacetamide induced | 50 mg/kg | 4 weeks | shh↓, Ihh↓, Ptch-1↓, Smo↓, Hhip↓, Gli-3↓, TNF-α↓ | [51] |
| Salvianolic acid A | Salvia miltiorrhiza | CCl4-induced | 20, 40 mg/kg | 6 weeks | NF-κB↓ (in the nucleus), p-NF-κBp65↓, IL-1β↓, IL-6↓, TNF-α↓, TGF-β↓, Cox-2↓, p-JAK1↓, p-STAT3↓ | [52] | |
| 5, 15 mg/kg | 6 weeks | p-AKT↓, p-mTOR↓, p-p70S6K1↓, caspase 3↓, Bax↓, α-SMA↓, PDGF-β↓, desmin↓, vimentin↓, TGF-β1↓, Bcl-2↑ | [53] | ||||
| Isochlorogenic acid A | Phenolic acid | Coffee beans, honeysuckle, tobacco leaves and kiwi | CCl4-induced | 10, 20, 40 mg/kg | 8 weeks | (NF-κB) p65↓, IκBα↓, HMGB1↓, TLR4↓, NF-κB↓ | [54] |
| Xanthohumol | Flavonoids | Hop derived | primary human hepatocytes (PHH) and HSC | 5 and 10 mM | 3 day | collagen type I↓, α-SMA↓, MCP-1↓, IL-8↓ | [55] |
| Naringin | Flavonoids | Grapefruit and oranges | thioacetamide (TAA)-induced | 40 mg/kg | 6 weeks | ROS↓, p-Akt↓, IL-6↓, caspase-3↓, IL-10↑ | [56] |
| Curcumin/Rutin | Flavonoids | Roots of curcumin spp/Flos Sophorae Immaturus | Hepatic stellate cells | PI3K-Class I↓, Akt↓, p-mTOR↓, TLRs↓, MyD88↓, NF-κB↓, TNF-α↓, IL-1β↓ | [57,58] | ||
| Procyanidin B2 | Flavonoid | Proanthocyanidin | CCl4-induced mouse | 50, 100 and 150 mg/kg | 4 weeks | VEGF↓, HIF-1α↓, α-SMA↓, Col-1↓, TGF-β1↓, CD31↓, Smo↓, GLI1↓ | [59] |
| human hepatic stellate cell (HSC) line (LX2 cells) | 60, 80 and 100 μM | 24 h | VEGF↓, HIF-1α↓, α-SMA↓, Col-1↓, TGF-β1↓ | ||||
| Hesperetin | Flavanone | Pericarp of citrus | CCl4-induced mouse | 25, 50, 100 mg/kg | 6 weeks | Ptch1↑, Col1α1↓, α-SMA↓, ALT↓, AST↓ | [60] |
| HSC-T6 cells | 2.50 μM | Ptch1↑, c-Myc↓, CyclinD1↓ | |||||
| Salvianolic acid B | Phenolic acid | Salvia miltiorrhiza | CCl4-induced mouse | 60 mg/kg | 6 weeks | ALB↓, ALT↓, AST↓, Shh↓, Ptch1↓, Smo ↓, Gli1↓, TGF-β1↓ | [61] |
| CCl4 | 100 mg/kg | 8 weeks | PTCH1↑, Smo↓, Gli2↓ | ||||
| Phillygenin | Phenolic acid | Weeping forsythia capsule | CCl4-induced mouse | 10, 20, 40 mg/kg | 4 weeks | LPS↓, IL-1 β↓, IL-6↓, TNF- α↓ | [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, J.; Huang, C.; Li, S.; Jiang, X.; Chang, H.; Li, M. Research Progress Regarding the Effect and Mechanism of Dietary Polyphenols in Liver Fibrosis. Molecules 2024, 29, 127. https://doi.org/10.3390/molecules29010127
Chang J, Huang C, Li S, Jiang X, Chang H, Li M. Research Progress Regarding the Effect and Mechanism of Dietary Polyphenols in Liver Fibrosis. Molecules. 2024; 29(1):127. https://doi.org/10.3390/molecules29010127
Chicago/Turabian StyleChang, Jiayin, Congying Huang, Siqi Li, Xiaolei Jiang, Hong Chang, and Minhui Li. 2024. "Research Progress Regarding the Effect and Mechanism of Dietary Polyphenols in Liver Fibrosis" Molecules 29, no. 1: 127. https://doi.org/10.3390/molecules29010127
APA StyleChang, J., Huang, C., Li, S., Jiang, X., Chang, H., & Li, M. (2024). Research Progress Regarding the Effect and Mechanism of Dietary Polyphenols in Liver Fibrosis. Molecules, 29(1), 127. https://doi.org/10.3390/molecules29010127