


Acetogenins-Rich Fractions of Annona coriacea Suppress Human Glioblastoma Viability and Migration by Regulating Necroptosis and MMP-2 Activity In Vitro

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
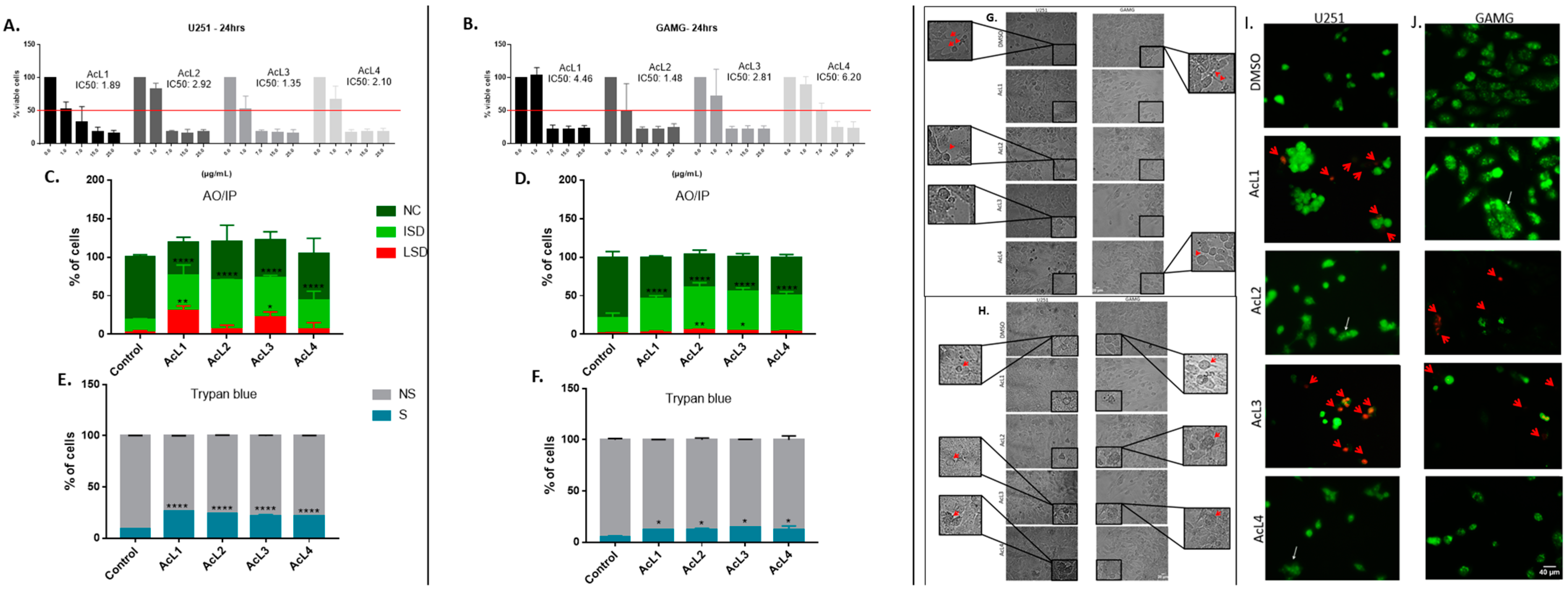
2.1. The Cell Viability of GBM Strains Decreased after Being Exposed to A. coriacea Fractions
2.2. A. coriacea Fractions Increased Morphological Alterations Associated with Necroptosis on GMB Cells Lines
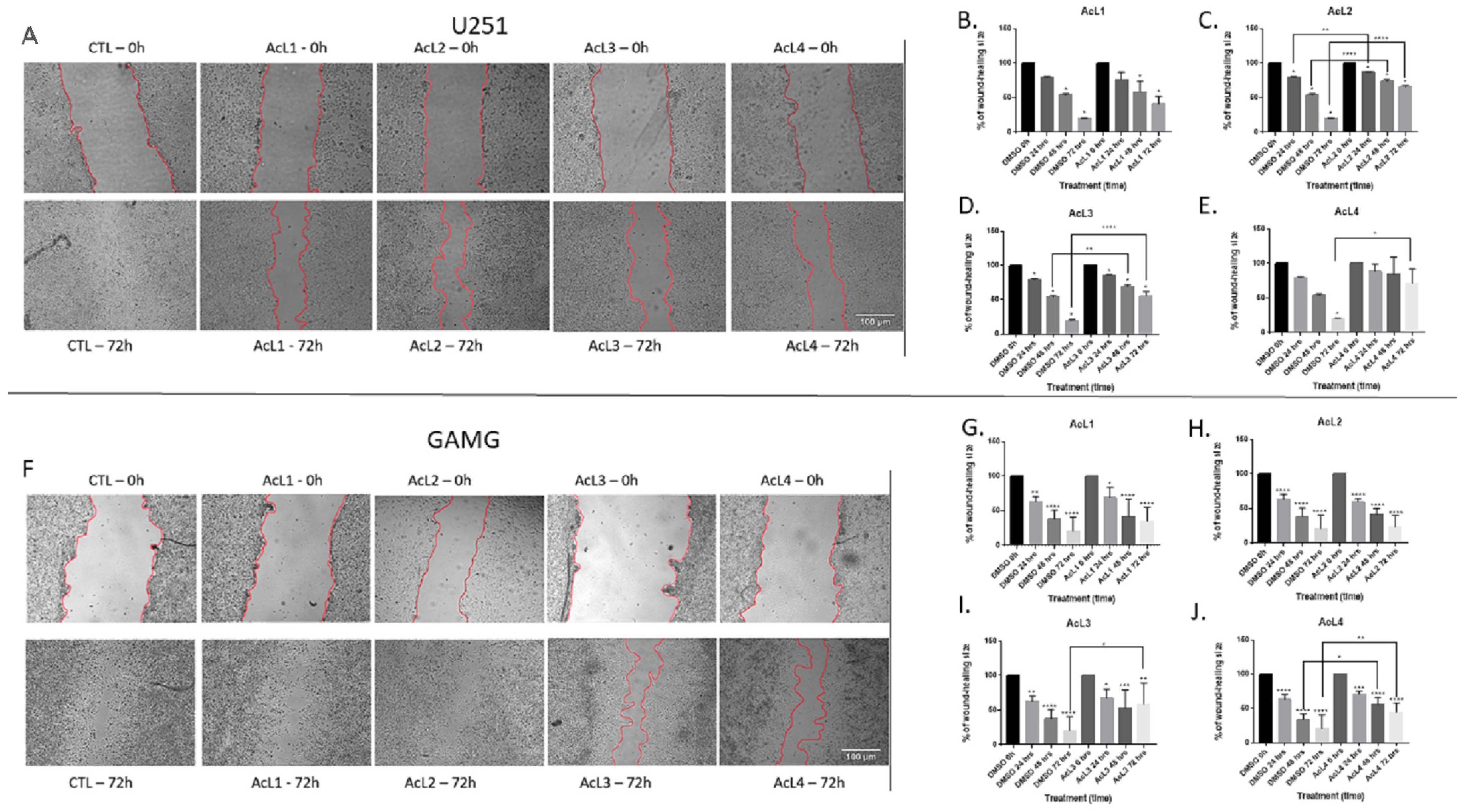
2.3. A. coriacea Fractions Inhibit Cells Migration of GBM Cell Lines
2.4. A. coriacea Fractions Is Selective for GBM Cell Lines
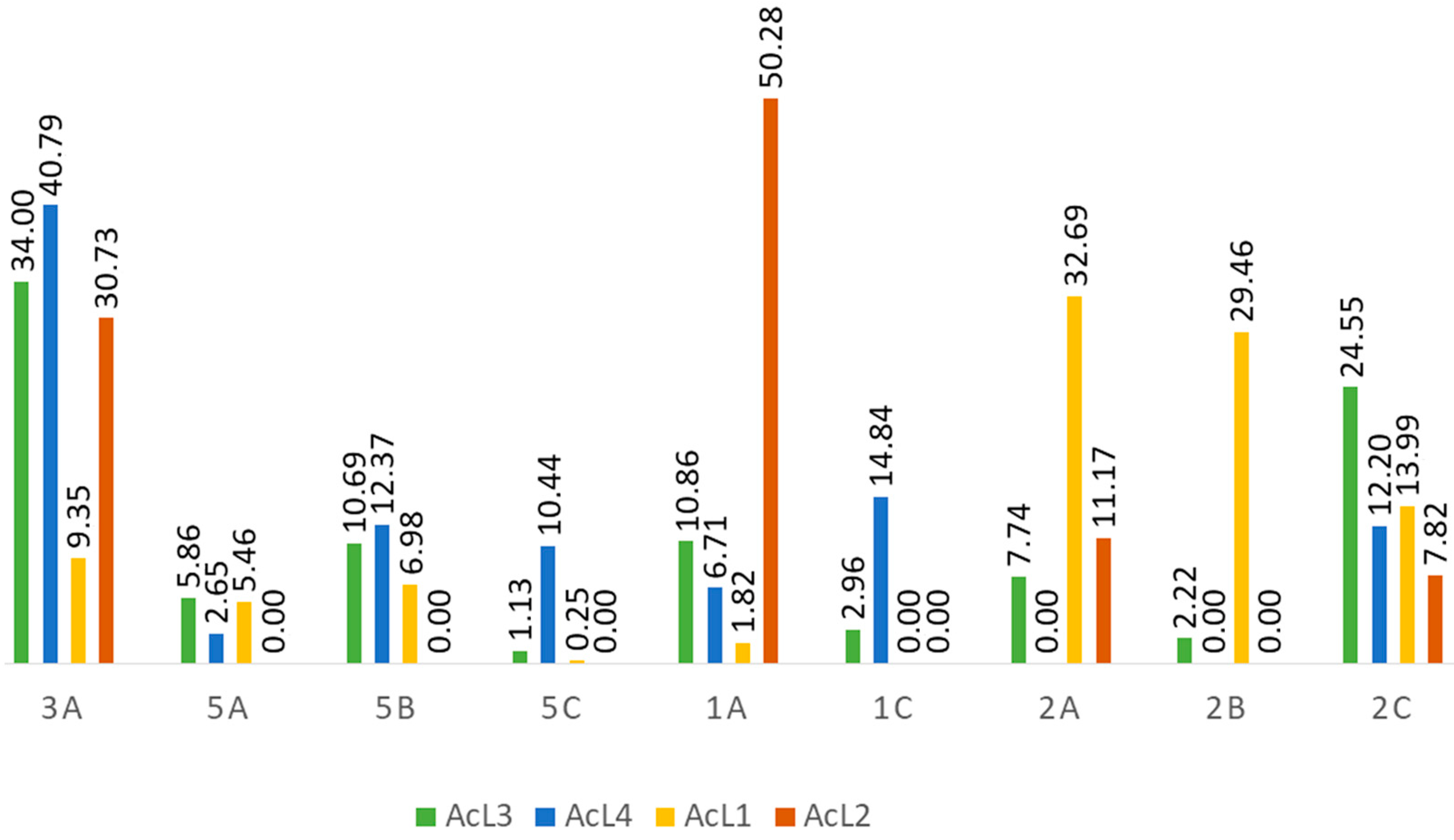
2.5. Acetogenins Are Present in All Fractions
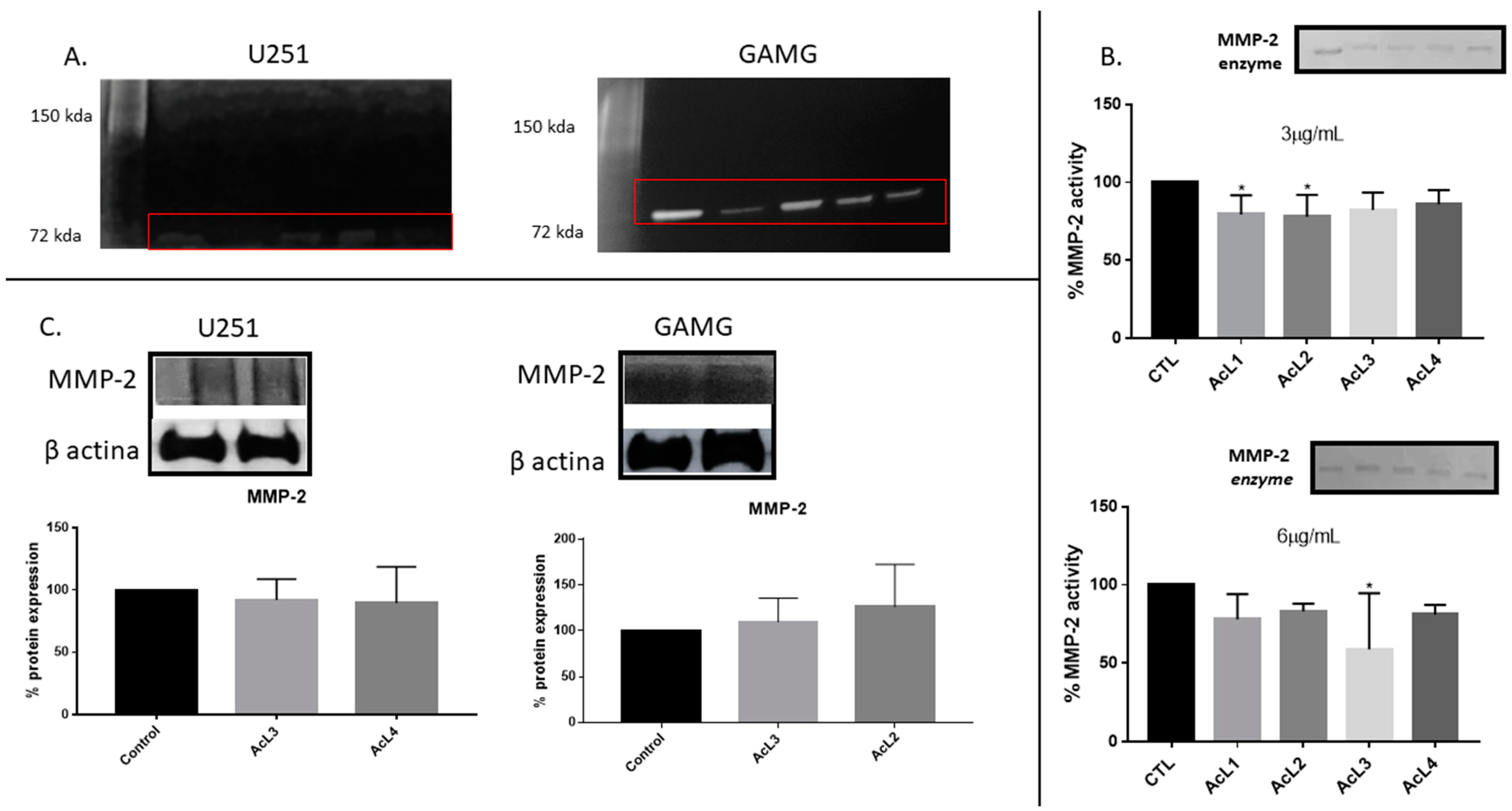
2.6. A. coriacea Fractions Inhibited Activity but Do Not Alter MMP-2 Expression on GBM Cells
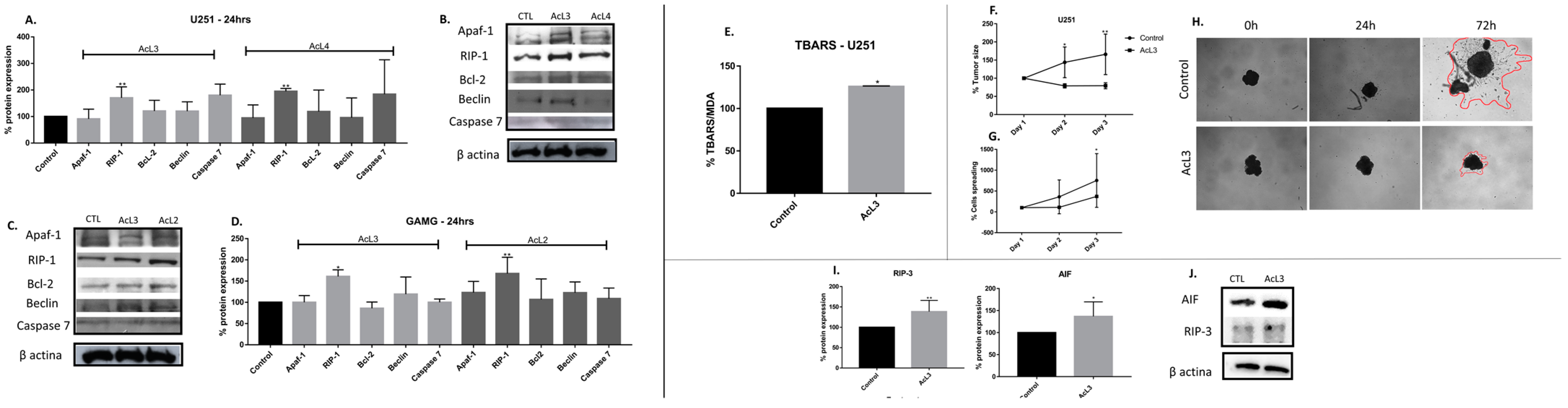
2.7. A. coriacea Fractions Induced Superexpression of RIP-1 on GBM Cell Lines
2.8. A. coriacea Fraction Increased TBARS Production, Inhibits Spheroids Growth and Induces Necroptosis by Overexpression of RIP-3 and AIF Proteins
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. Chemical Characterization of the Acetogenins from A. coriacea Fractions
4.3. Cell lines and Cultures and Treatments
4.4. MTT Assay and Selectivity Index
4.5. Trypan Blue Exclusion Viable Cell Assay
4.6. Acridine Orange/Propidium Iodide Double Staining Cell Death Assessment
4.7. Wound-Healing Assay
4.8. Chemical Characterization of the Acetogenins from A. coriacea Fractions
4.9. Zymography
4.10. Analysis of Protein Expression by Western Blotting
4.11. Expression of TBARS
4.12. Spheroids Formation
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Taylor, O.G.; Brzozowski, J.S.; Skelding, K.A. Glioblastoma Multiforme: An Overview of Emerging Therapeutic Targets. Front. Oncol. 2019, 9, 963. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, Y.; Banerjee, A.; Pal, I.; Biswas, A.; Das, S.; Dey, K.K.; Kapoor, N.; Ghosh, A.K.; Mitra, P.; Mandal, M. Delineation of Crosstalk between HSP27 and MMP-2/MMP-9: A Synergistic Therapeutic Avenue for Glioblastoma Management. Biochim. Biophys. Acta (BBA) Gen. Subj. 2019, 1863, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Roomi, M.W.; Kalinovsky, T.; Rath, M.; Niedzwiecki, A. Modulation of MMP-2 and MMP-9 Secretion by Cytokines, Inducers and Inhibitors in Human Glioblastoma T-98G Cells. Oncol. Rep. 2017, 37, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.N.d.S.A.O.; Dutra, L.M.; Lorenzo, V.P.; Almeida, J.R.G.D.S. Phytochemicals and Biological Properties of Annona Coriacea Mart. (Annonaceae): A Systematic Review from 1971 to 2020. Chem. Biol. Interact. 2021, 336, 109390. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.-C.; Liou, J.-R.; Wu, T.-Y.; Chang, F.-R.; Wu, Y.-C. Acetogenins from Annonaceae. Prog. Chem. Org. Nat. Prod. 2016, 101, 113–230. [Google Scholar] [CrossRef]
- Li, Y.; Ye, J.; Chen, Z.; Wen, J.; Li, F.; Su, P.; Lin, Y.; Hu, B.; Wu, D.; Ning, L.; et al. Annonaceous Acetogenins Mediated Up-Regulation of Notch2 Exerts Growth Inhibition in Human Gastric Cancer Cells in Vitro. Oncotarget 2017, 8, 21140–21152. [Google Scholar] [CrossRef][Green Version]
- Bermejo, A.; Figadere, B.; Zafra-Polo, M.-C.; Barrachina, I.; Estornell, E.; Cortes, D. Acetogenins from Annonaceae: Recent Progress in Isolation, Synthesis and Mechanisms of Action. Nat. Prod. Rep. 2005, 22, 269–303. [Google Scholar] [CrossRef]
- Jacobo-Herrera, N.; Pérez-Plasencia, C.; Castro-Torres, V.A.; Martínez-Vázquez, M.; González-Esquinca, A.R.; Zentella-Dehesa, A. Selective Acetogenins and Their Potential as Anticancer Agents. Front. Pharmacol. 2019, 10, 783. [Google Scholar] [CrossRef]
- Terezan, A.P.; Junqueira, J.G.M.; Wakui, V.G.; Kato, L.; Oliveira, C.M.A.; Martins, C.H.G.; Santiago, M.B.; Severino, V.G.P. Qualitative Analysis of the Acetogenins from Annona coriacea (Annonaceae) Leaves by HPLC-Q-Orbitrap and Their Antibacterial Potential against Oral Pathogens. Nat. Prod. Res. 2020, 36, 765–771. [Google Scholar] [CrossRef]
- Gutiérrez, M.T.; Durán, A.G.; Mejías, F.J.R.; Molinillo, J.M.G.; Megias, D.; Valdivia, M.M.; Macías, F.A. Bio-Guided Isolation of Acetogenins from Annona cherimola Deciduous Leaves: Production of Nanocarriers to Boost the Bioavailability Properties. Molecules 2020, 25, 4861. [Google Scholar] [CrossRef]
- Naik, A.V.; Sellappan, K. Assessment of Genotoxic Potential of Annonacin and Annona muricata L. Extracts on Human Breast Cancer (MCF-7) Cells. Adv. Tradit. Med. 2020, 21, 779–789. [Google Scholar] [CrossRef]
- Yap, C.V.; Subramaniam, K.S.; Khor, S.W.; Chung, I. Annonacin Exerts Antitumor Activity through Induction of Apoptosis and Extracellular Signal-Regulated Kinase Inhibition. Pharmacogn. Res. 2017, 9, 378–383. [Google Scholar] [CrossRef]
- Yiallouris, A.; Patrikios, I.; Johnson, E.O.; Sereti, E.; Dimas, K.; Ford, C.D.; Fedosova, N.U.; Graier, W.F.; Sokratous, K.; Kyriakou, K.; et al. Annonacin Promotes Selective Cancer Cell Death via NKA-Dependent and SERCA-Dependent Pathways. Cell Death Dis. 2018, 9, 764. [Google Scholar] [CrossRef]
- Ratnayake, S.; Gu, Z.-M.; Miesbauer, L.; Smith, D.; Wood, K.; Evert, D.; McLaughlin, J. Parvifloracin and Parviflorin: Cytotoxic Bistetrahydrofuran Acetogenins with 35 Carbons from Asimina parviflora (Annonaceae). Can. J. Chem. 2011, 72, 287–293. [Google Scholar] [CrossRef]
- Wu, F.E.; Zhao, G.X.; Zeng, L.; Zhang, Y.; Schwedler, J.T.; McLaughlin, J.L.; Sastrodihardjo, S. Additional Bioactive Acetogenins, Annomutacin and (2,4-Trans and Cis)-10R-Annonacin-A-Ones, from the Leaves of Annona muricata. J. Nat. Prod. 1995, 58, 1430–1437. [Google Scholar] [CrossRef]
- Zhao, G.X.; Rieser, M.J.; Hui, Y.H.; Miesbauer, L.R.; Smith, D.L.; McLaughlin, J.L. Biologically Active Acetogenins from Stem Bark of Asimina Triloba. Phytochemistry 1993, 33, 1065–1073. [Google Scholar] [CrossRef]
- Liaw, C.-C.; Chang, F.-R.; Wu, C.-C.; Chen, S.-L.; Bastow, K.F.; Hayashi, K.; Nozaki, H.; Lee, K.-H.; Wu, Y.-C. Nine New Cytotoxic Monotetrahydrofuranic Annonaceous Acetogenins from Annona montana. Planta Med. 2004, 70, 948–959. [Google Scholar] [CrossRef]
- Rieser, M.J.; Gu, Z.M.; Fang, X.P.; Zeng, L.; Wood, K.V.; McLaughlin, J.L. Five Novel Mono-Tetrahydrofuran Ring Acetogenins from the Seeds of Annona muricata. J. Nat. Prod. 1996, 59, 100–108. [Google Scholar] [CrossRef]
- Liu, X.X.; Alali, F.Q.; Pilarinou, E.; McLaughlin, J.L. Glacins A and B: Two Novel Bioactive Mono-Tetrahydrofuran Acetogenins from Annona glabra. J. Nat. Prod. 1998, 61, 620–624. [Google Scholar] [CrossRef]
- Woo, M.H.; Kim, D.H.; McLaughlin, J.L. Asitrilobins A and B: Cytotoxic Mono-THF Annonaceous Acetogenins from the Seeds of Asimina Triloba. Phytochemistry 1999, 50, 1033–1040. [Google Scholar] [CrossRef]
- Kim, E.-J.; Tian, F.; Woo, M.-H. Asitrocin, (2,4)-Cis- and Trans-Asitrocinones: Novel Bioactive Mono-Tetrahydrofuran Acetogenins from Asimina Triloba Seeds. J. Nat. Prod. 2000, 63, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Tantithanaporn, S.; Wattanapiromsakul, C.; Itharat, A.; Keawpradub, N. Cytotoxic Activity of Acetogenins and Styryl Lactones Isolated from Goniothalamus undulatus Ridl. Root Extracts against a Lung Cancer Cell Line (COR-L23). Phytomedicine 2011, 18, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.V.; Pinheiro, M.L.B.; Xavier, C.M.; Silva, J.R.A.; Amaral, A.C.F.; Souza, A.D.L.; Barison, A.; Campos, F.R.; Ferreira, A.G.; Machado, G.M.C.; et al. A Pyrimidine-β-Carboline and Other Alkaloids from Annona foetida with Antileishmanial Activity. J. Nat. Prod. 2006, 69, 292–294. [Google Scholar] [CrossRef]
- Alves, T.; Gonçalves, M.; Correia, F.; Silva, V.; Sousa, P.; Carvalho, M.; Braz-Filho, R.; Dall’Oglio, E. New Acetogenins from the Seeds of Annona coriacea. Helv. Chim. Acta 2014, 97, 1469–1474. [Google Scholar] [CrossRef]
- Jolad, S.D.; Hoffmann, J.J.; Cole, J.R.; Barry, C.E.I.; Bates, R.B.; Linz, G.S.; Konig, W.A. Desacetyluvaricin from Uvaria accuminata, Configuration of Uvaricin at C-36. J. Nat. Prod. 1985, 48, 644–645. [Google Scholar] [CrossRef] [PubMed]
- Mangal, M.; Khan, M.I.; Agarwal, S.M. Acetogenins as Potential Anticancer Agents. Anticancer Agents Med. Chem. 2015, 16, 138–159. [Google Scholar] [CrossRef]
- Itharat, A.; Houghton, P.J.; Eno-Amooquaye, E.; Burke, P.J.; Sampson, J.H.; Raman, A. In Vitro Cytotoxic Activity of Thai Medicinal Plants Used Traditionally to Treat Cancer. J. Ethnopharmacol. 2004, 90, 33–38. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental Regulation of Tumor Progression and Metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Ragasa, C.Y.; Soriano, G.; Torres, O.B.; Don, M.-J.; Shen, C.-C. Acetogenins from Annona muricata. Pharmacogn. J. 2012, 4, 32–37. [Google Scholar] [CrossRef]
- Sasaki, S.; Maruta, K.; Naito, H.; Sugihara, H.; Hiratani, K.; Maeda, M. New Calcium-Selective Electrodes Based on Annonaceous Acetogenins and Their Analogs with Neighboring Bistetrahydrofuran. Tetrahedron Lett. 1995, 36, 5571–5574. [Google Scholar] [CrossRef]
- Hazra, S.; Guha, R.; Jongkey, G.; Palui, H.; Mishra, A.; Vemuganti, G.K.; Basak, S.K.; Mandal, T.K.; Konar, A. Modulation of Matrix Metalloproteinase Activity by EDTA Prevents Posterior Capsular Opacification. Mol. Vis 2012, 18, 1701–1711. [Google Scholar]
- Belizário, J.; Vieira-Cordeiro, L.; Enns, S. Necroptotic Cell Death Signaling and Execution Pathway: Lessons from Knockout Mice. Available online: https://www.hindawi.com/journals/mi/2015/128076/ref/ (accessed on 30 January 2019).
- Lee, Y.-J.; Park, K.-S.; Nam, H.-S.; Cho, M.-K.; Lee, S.-H. Apigenin Causes Necroptosis by Inducing ROS Accumulation, Mitochondrial Dysfunction, and ATP Depletion in Malignant Mesothelioma Cells. Korean J. Physiol. Pharmacol. 2020, 24, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Agu, K.C.; Okolie, N.P.; Falodun, A.; Engel-Lutz, N. In Vitro Anticancer Assessments of Annona muricata Fractions and in vitro Antioxidant Profile of Fractions and Isolated Acetogenin (15-Acetyl Guanacone). J. Cancer Res. Pract. 2018, 5, 53–66. [Google Scholar] [CrossRef]
- de Pedro, N.; Cautain, B.; Melguizo, A.; Vicente, F.; Genilloud, O.; Peláez, F.; Tormo, J.R. Mitochondrial Complex I Inhibitors, Acetogenins, Induce HepG2 Cell Death through the Induction of the Complete Apoptotic Mitochondrial Pathway. J. Bioenerg. Biomembr. 2013, 45, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.N.F.; Silva-Oliveira, R.J.; Oliveira Silva, V.A.; Rosa, M.N.; Vital, P.S.; Barbosa, M.C.S.; dos Santos, F.V.; Junqueira, J.G.M.; Severino, V.G.P.; Oliveira, B.G.; et al. Annona coriacea Mart. Fractions Promote Cell Cycle Arrest and Inhibit Autophagic Flux in Human Cervical Cancer Cell Lines. Molecules 2019, 24, 3963. [Google Scholar] [CrossRef]
- Torres, M.P.; Rachagani, S.; Purohit, V.; Pandey, P.; Joshi, S.; Moore, E.D.; Johansson, S.L.; Singh, P.K.; Ganti, A.K.; Batra, S.K. Graviola: A Novel Promising Natural-Derived Drug That Inhibits Tumorigenicity and Metastasis of Pancreatic Cancer Cells in Vitro and in Vivo through Altering Cell Metabolism. Cancer Lett. 2012, 323, 29–40. [Google Scholar] [CrossRef]
- Han, B.; Wang, T.-D.; Shen, S.-M.; Yu, Y.; Mao, C.; Yao, Z.-J.; Wang, L.-S. Annonaceous Acetogenin Mimic AA005 Induces Cancer Cell Death via Apoptosis Inducing Factor through a Caspase-3-Independent Mechanism. BMC Cancer 2015, 15, 139. [Google Scholar] [CrossRef]
- Ong, C.S.; Zhou, X.; Han, J.; Huang, C.Y.; Nashed, A.; Khatri, S.; Mattson, G.; Fukunishi, T.; Zhang, H.; Hibino, N. In Vivo Therapeutic Applications of Cell Spheroids. Biotechnol. Adv. 2018, 36, 494–505. [Google Scholar] [CrossRef]
- Alexandre, G.P.; Simão, J.L.S.; Tavares, M.O.A.; Zuffo, I.M.S.; Prado, S.V.; DE Paiva, J.A.; Mustapha, A.N.; DE Oliveira, A.E.; Kato, L.; Severino, V.G.P. Dereplication by HPLC-ESI-MS and Antioxidant Activity of Phenolic Compounds from Banisteriopsis laevifolia (Malpighiaceae). An. Acad. Bras. Ciências 2022, 94, e20201844. [Google Scholar] [CrossRef]
- Silva, A.G.; Lopes, C.F.B.; Carvalho Júnior, C.G.; Thomé, R.G.; dos Santos, H.B.; Reis, R.; de Azambuja Ribeiro, R.I.M. WIN55,212-2 Induces Caspase-Independent Apoptosis on Human Glioblastoma Cells by Regulating HSP70, P53 and Cathepsin D. Toxicol. Vitr. 2019, 57, 233–243. [Google Scholar] [CrossRef]
- Ogbole, O.O.; Segun, P.A.; Adeniji, A.J. In Vitro Cytotoxic Activity of Medicinal Plants from Nigeria Ethnomedicine on Rhabdomyosarcoma Cancer Cell Line and HPLC Analysis of Active Extracts. BMC Complement. Altern. Med. 2017, 17, 494. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Ismail, M.; Imam, M.U.; Azmi, N.H.; Fathy, S.F.; Foo, J.B.; Abu Bakar, M.F. Mechanistic Basis for Protection of Differentiated SH-SY5Y Cells by Oryzanol-Rich Fraction against Hydrogen Peroxide-Induced Neurotoxicity. BMC Complement. Altern. Med. 2014, 14, 467. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, K.M.; Nunes, D.A.D.F.; Gomes, I.N.D.F.; Da Silva, S.L.; Ribeiro, R.I.M.D.A. Inhibition of Gelatinase Activity of MMP-2 and MMP-9 by Extracts of Bauhinia ungulata L. Biosci. J. 2015, 31, 584–590. [Google Scholar] [CrossRef]
- Silva, A.G.; Silva, V.A.O.; Oliveira, R.J.S.; de Rezende, A.R.; Chagas, R.C.R.; Pimenta, L.P.S.; Romão, W.; Santos, H.B.; Thomé, R.G.; Reis, R.M.; et al. Matteucinol, Isolated from Miconia Chamissois, Induces Apoptosis in Human Glioblastoma Lines via the Intrinsic Pathway and Inhibits Angiogenesis and Tumor Growth in Vivo. Investig. New Drugs 2020, 38, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Ernster, L.; Nordenbrand, K. [92a] Microsomal Lipid Peroxidation. In Methods in Enzymology; Oxidation and Phosphorylation; Academic Press: Cambridge, MA, USA, 1967; Volume 10, pp. 574–580. [Google Scholar]
- Foty, R. A Simple Hanging Drop Cell Culture Protocol for Generation of 3D Spheroids. J. Vis. Exp. 2011, e2720. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time | Compound | Fractions | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AcL3 | AcL4 | AcL1 | AcL2 | ||||||||||||||||||
| m/z | Erro (ppm) | Peak Height | Peak Area | m/z | Erro (ppm) | Peak Height | Peak Area | m/z | Erro (ppm) | Peak Height | Peak Area | m/z | Erro (ppm) | Peak Height | Peak Area | ||||||
| 7.24 | 3a | 645.45862 | 1.30 | 2.74 × 108 | 2.16 × 109 | 645.45795 | 0.26 | 3.94 × 108 | 3.16 × 109 | 645.45868 | 1.40 | 8.22 × 107 | 1.13 × 109 | 645.45831 | 0.82 | 9.1 × 107 | 1.1 × 109 | ||||
| 15.96 | 5a | 613.46798 | 0.05 | 1.2 × 108 | 3.72 × 108 | 613.46802 | 0.11 | 1.33 × 108 | 2.05 × 108 | 613.46875 | 1.31 | 7.51 × 107 | 6.6 × 108 | - | - | - | - | ||||
| 17.24 | 5b | 613.46887 | 1.50 | 4.99 × 107 | 6.79 × 108 | 613.46863 | 1.11 | 7.14 × 107 | 9.58 × 108 | 613.46838 | 0.71 | 3.10 × 107 | 8.43 × 108 | - | - | - | - | ||||
| 18.95 | 5c | 635.45056 | 1.05 | 2.81 × 107 | 7.15 × 107 | 635.44995 | 0.09 | 3.57 × 107 | 8.09 × 108 | 635.45050 | 0.96 | 1.94 × 107 | 3.0 × 107 | - | - | - | - | ||||
| 22.70 | 1a | 639.48425 | 1.01 | 7.20 × 107 | 6.90 × 108 | 639.48401 | 0.64 | 4.80 × 107 | 5.20 × 108 | 639.48407 | 0.74 | 7.40 × 107 | 2.2 × 108 | 639.48425 | 1.01 | 4.50 × 107 | 1.80 × 109 | ||||
| 24.62 | 1c | 661.46613 | 0.89 | 1.22 × 108 | 1.88 × 108 | 661.46552 | −0.03 | 6.14 × 107 | 1.15 × 109 | - | - | - | - | - | - | - | - | ||||
| 25.60 | 2a | 619.45569 | 1.16 | 1.45 × 107 | 4.92 × 108 | - | - | - | - | 619.45593 | 1.54 | 4.13 × 107 | 3.95 × 109 | 619.45538 | 0.66 | 2.50 × 107 | 4.00 × 108 | ||||
| 28.30 | 2b | 597.47363 | 1.00 | 4.58 × 107 | 1.41 × 108 | - | - | - | - | 597.47351 | 0.80 | 3.03 × 108 | 3.56 × 109 | - | - | - | - | ||||
| 29.17 | 2c | 597.47406 | 1.73 | 1.10 × 108 | 1.56 × 109 | 597.47314 | 0.18 | 4.86 × 107 | 9.45 × 108 | 597.47406 | 1.72 | 2.24 × 108 | 1.69 × 109 | 597.47416 | 1.86 | 9.50 × 107 | 2.80 × 108 | ||||
| Classes of Compounds | Possible Metabolites | References |
|---|---|---|
| 2c | annonacin, goniothalamicin. annonacin-A, annoreticuin, cis-annonacin, cis-goniothalamicin, arianacin, javoracin, glacin-A, glacin-B, asitrilobin-B, rolliacocin, asitrocin, montalicin-C, montalicin-D, cis-annoreticuim, montalicin-F | [10,11,12,13,14,15,16,17,18,19,20,21,22] |
| 3a | coriheptocin-A, coriheptocin-B | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, L.R.; Oliveira, A.G.S.; Arantes, A.; Junqueira, J.G.M.; Alexandre, G.P.; Severino, V.G.P.; Reis, R.M.; Kim, B.; Ribeiro, R.I.M.A. Acetogenins-Rich Fractions of Annona coriacea Suppress Human Glioblastoma Viability and Migration by Regulating Necroptosis and MMP-2 Activity In Vitro. Molecules 2023, 28, 3809. https://doi.org/10.3390/molecules28093809
Sousa LR, Oliveira AGS, Arantes A, Junqueira JGM, Alexandre GP, Severino VGP, Reis RM, Kim B, Ribeiro RIMA. Acetogenins-Rich Fractions of Annona coriacea Suppress Human Glioblastoma Viability and Migration by Regulating Necroptosis and MMP-2 Activity In Vitro. Molecules. 2023; 28(9):3809. https://doi.org/10.3390/molecules28093809
Chicago/Turabian StyleSousa, Lorena R., Ana Gabriela S. Oliveira, Antônio Arantes, João Gabriel M. Junqueira, Gerso P. Alexandre, Vanessa G. P. Severino, Rui Manuel Reis, Bonglee Kim, and Rosy I. M. A. Ribeiro. 2023. "Acetogenins-Rich Fractions of Annona coriacea Suppress Human Glioblastoma Viability and Migration by Regulating Necroptosis and MMP-2 Activity In Vitro" Molecules 28, no. 9: 3809. https://doi.org/10.3390/molecules28093809
APA StyleSousa, L. R., Oliveira, A. G. S., Arantes, A., Junqueira, J. G. M., Alexandre, G. P., Severino, V. G. P., Reis, R. M., Kim, B., & Ribeiro, R. I. M. A. (2023). Acetogenins-Rich Fractions of Annona coriacea Suppress Human Glioblastoma Viability and Migration by Regulating Necroptosis and MMP-2 Activity In Vitro. Molecules, 28(9), 3809. https://doi.org/10.3390/molecules28093809