
Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
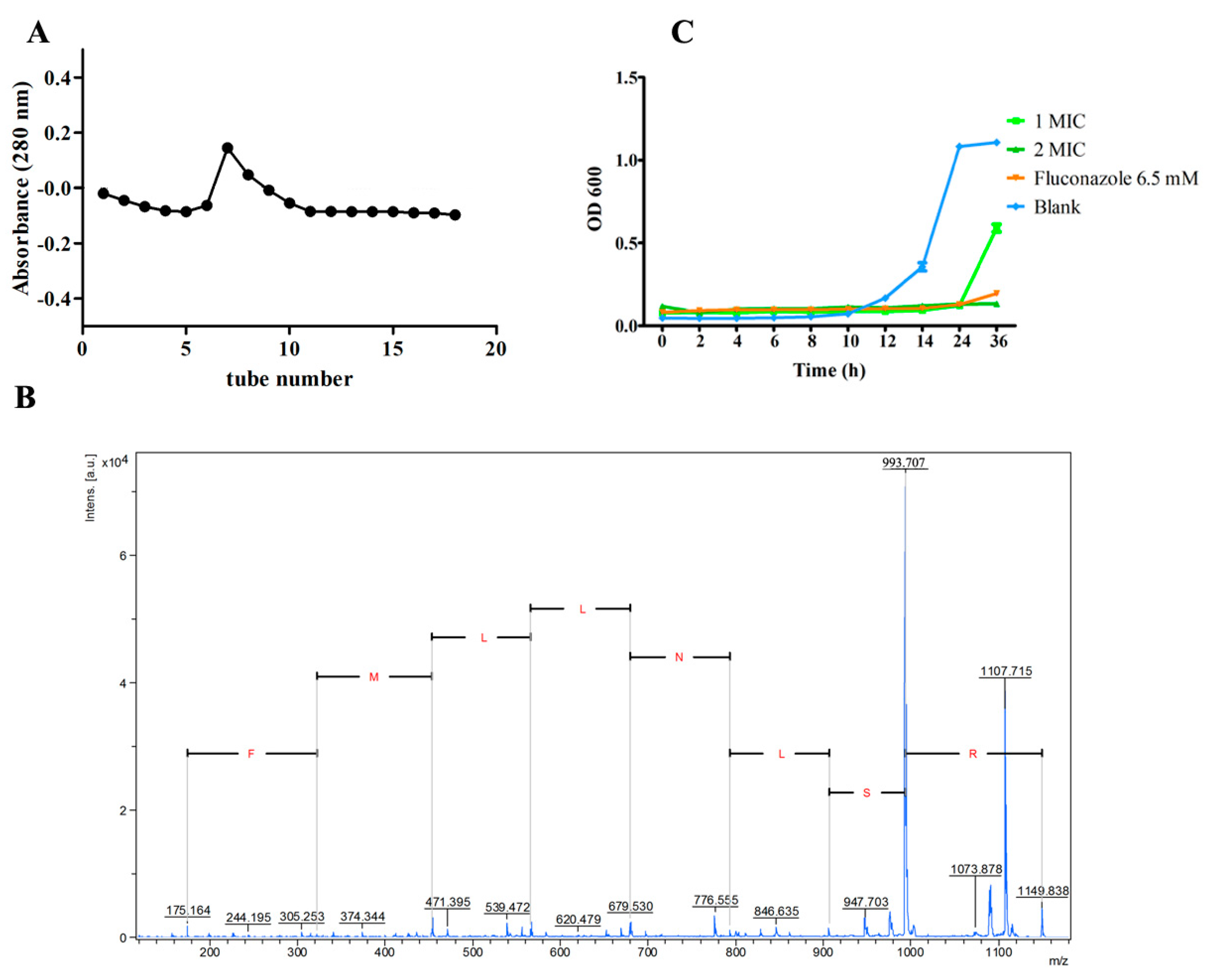
2.1. Peptide Isolation and Identification
2.2. Antifungal Activity of NpRS
2.3. Structure and Conformation Characterization of NpRS
2.4. Antifungal Mechanism of NpRS
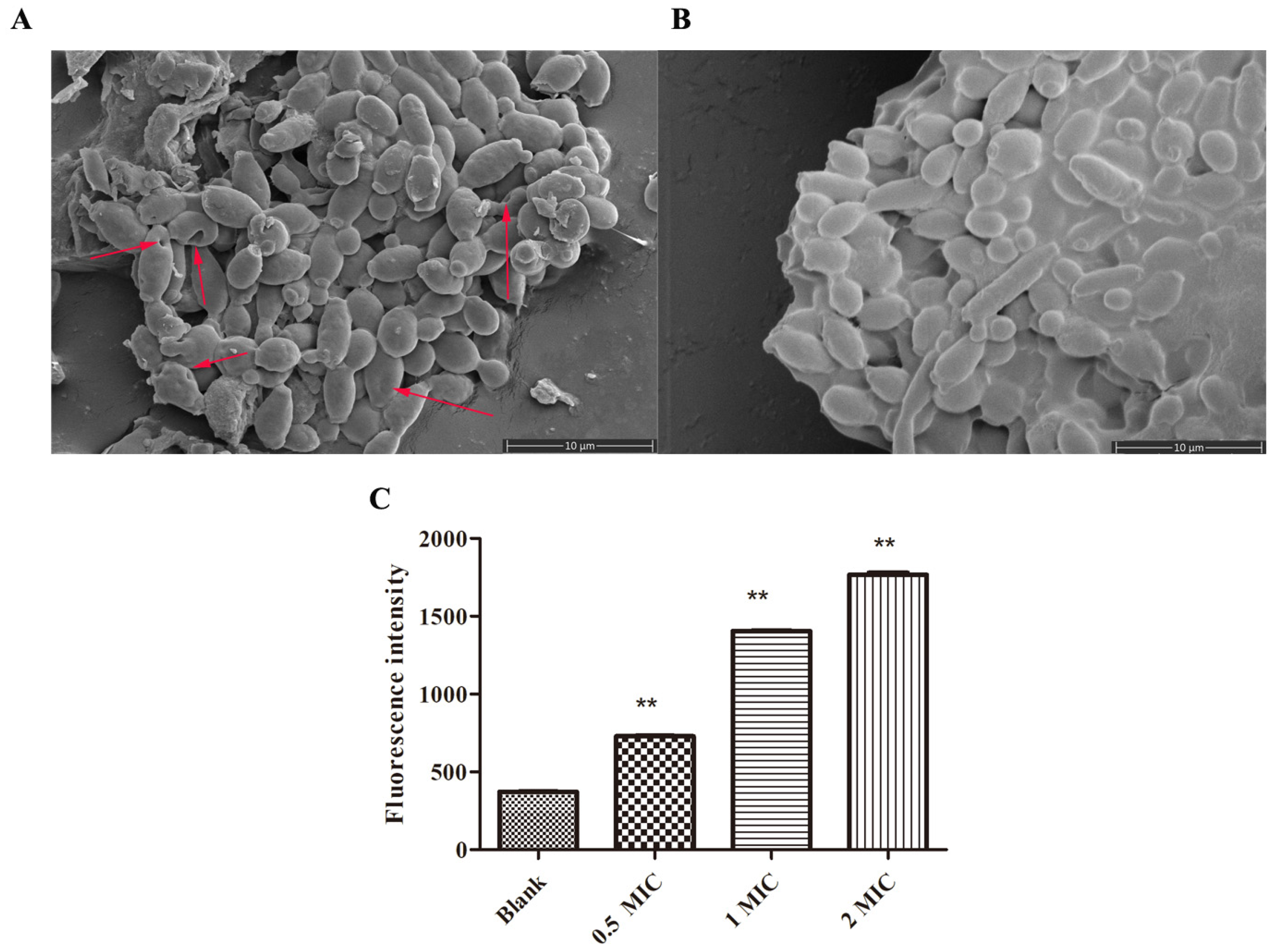
2.4.1. SEM and Membrane Potential Studies for C. albicans after NpRS Treatment
2.4.2. Transcriptomic Analysis
2.4.3. Drug Resistance Analysis
2.5. Cytotoxicity of NpRS against Mammalian Cell
3. Materials and Methods
3.1. Material and Reagents
3.2. Extraction and Isolation of NpRS
3.3. Peptides’ Identification by MALDI-TOF MS/MS Analysis
3.4. Antifungal Activity Evaluation by Agar Diffusion Assay
3.5. Minimum Inhibition Concentration (MIC) Assay
3.6. Time-Kill Kinetics Analysis
3.7. Synergetic Interaction Analysis with Fluconazole
3.8. Secondary Structure Analysis by Circular Dichroism (CD)
3.9. Spatial Structure Analysis by Nuclear Magnetic Resonance (NMR)
3.10. Scanning Electron Microscopy (SEM) Analysis
3.11. Membrane Depolarization Analysis
3.12. Transcriptome Analysis
3.13. Drug Resistance Evaluation by Sequential Passaging
3.14. Cytotoxicity Assay against Mammalian Cell
3.15. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Molecular Mechanisms in Candida albicans and Beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B. Evolution of antifungal-drug resistance: Mechanisms and pathogen fitness. Nat. Rev. Microbiol. 2005, 3, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Krysan, D.J. Antifungal Drug Development: Challenges, Unmet Clinical Needs, and New Approaches. Cold Spring Harb. Perspect. Med. 2014, 4, a019703. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, E383–E392. [Google Scholar] [CrossRef]
- Soltani, S.; Keymanesh, K.; Sardari, S. In silico analysis of antifungal peptides: Determining the lead template sequence of potent antifungal peptides. Expert Opin. Drug Discov. 2007, 2, 837–847. [Google Scholar] [CrossRef]
- Egorov, T.A.; Odintsova, T.I.; Pukhalsky, V.A.; Grishin, E.V. Diversity of wheat anti-microbial peptides. Peptides 2005, 26, 2064–2073. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Schaaper, W.M.M.; Posthuma, G.A.; Plasman, H.H.; Sijtsma, L.; Fant, F.; Borremans, F.A.M.; Thevissen, K.; Broekaert, W.F.; Meloen, R.H.; van Amerongen, A. Synthetic peptides derived from the beta(2)-beta(3) loop of Raphanus sativus antifungal protein 2 that mimic the active site. J. Pept. Res. 2001, 57, 409–418. [Google Scholar] [CrossRef]
- Dlozi, P.N.; Gladchuk, A.; Crutchley, R.D.; Keuler, N.; Coetzee, R.; Dube, A. Cathelicidins and defensins antimicrobial host defense peptides in the treatment of TB and HIV: Pharmacogenomic and nanomedicine approaches towards improved therapeutic outcomes. Biomed. Pharmacother. 2022, 151, 113189. [Google Scholar] [CrossRef]
- Lee, M.-R.; Raman, N.; Gellman, S.H.; Lynn, D.M.; Palecek, S.P. Hydrophobicity and Helicity Regulate the Antifungal Activity of 14-Helical beta-Peptides. ACS Chem. Biol. 2014, 9, 1613–1621. [Google Scholar] [CrossRef]
- Li, S.; Li, N.; Wang, Y.; Wang, Q.; Li, R.; Zhang, M.; Panichayupakaranant, P.; Gao, Y.; Chen, H. Structural, functional and proteomic differences of proteins extracted from white garlic and Laba garlic. Food Res. Int. 2022, 155, 111047. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-R.; Shi, Q.-S.; Liang, Q.; Huang, X.-M.; Chen, Y.-B. Antifungal effect and mechanism of garlic oil on Penicillium funiculosum. Appl. Microbiol. Biotechnol. 2014, 98, 8337–8346. [Google Scholar] [CrossRef] [PubMed]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; GWasef, L.; Elewa, Y.H.; AAl-Sagan, A.; Abd El-Hack, M.E.; Taha, A.E.; MAbd-Elhakim, Y.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Int. J. Appl. Mech. 2020, 12, 872. [Google Scholar] [CrossRef]
- Balach, O.; Gazel, D.; Eksi, F.; Zer, Y.; Karsligil, T.; Al Azzawi, S. In Vitro Effects of Natural Garlic juice on Some Fungal Strains. Eur. J. Ther. 2020, 26, 298–302. [Google Scholar] [CrossRef]
- Gong, X.; Su, X.; Liu, H. Diallyl Trisulfide, the Antifungal Component of Garlic Essential Oil and the Bioactivity of Its Nanoemulsions Formed by Spontaneous Emulsification. Molecules 2021, 26, 7186. [Google Scholar] [CrossRef]
- Ledezma, E.; Apitz-Castro, R. Ajoene the main active compound of garlic (Allium sativum): A new antifungal agent. Rev. Iberoam. De Micol. 2006, 23, 75–80. [Google Scholar] [CrossRef]
- Al-Ghanayem, A.A.; Joseph, B.; Scaria, B.; Alhussaini, M.S. Phytochemical analysis and synergetic effects on antifungal properties of garlic and ginger extracts along with honey and lemon against Candida spp. Biosci. Res. 2019, 16, 3413–3420. [Google Scholar]
- Prajapati, M.; Shah, M.; Ranginwala, A.; Agrawal, P.; Acharya, D.; Thakkar, S. Antifungal effects of tulsi, garlic, cinnamon and lemongrass in powder and oil form on Candida albicans: An in vitro study. J. Oral Maxillofac. Pathol. 2021, 25, 306–312. [Google Scholar]
- Gao, X.; Chen, Y.; Chen, Z.; Xue, Z.; Jia, Y.; Guo, Q.; Ma, Q.; Zhang, M.; Chen, H. Identification and antimicrobial activity evaluation of three peptides from laba garlic and the related mechanism. Food Funct. 2019, 10, 4486–4496. [Google Scholar] [CrossRef]
- Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.P.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7047. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Xue, Z.; Jia, Y.; Li, R.; He, C.; Chen, H. The structure-mechanism relationship and mode of actions of antimicrobial peptides: A review. Trends Food Sci. Technol. 2021, 109, 103–115. [Google Scholar] [CrossRef]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and arginine-rich antimicrobial peptides: Structures and mechanisms of action. Biochim. Et Biophys. Acta-Biomembr. 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [PubMed]
- Petit, V.W.; Rolland, J.-L.; Blond, A.; Cazevieille, C.; Djediat, C.; Peduzzi, J.; Goulard, C.; Bachere, E.; Dupont, J.; Destoumieux-Garzon, D.; et al. A hemocyanin-derived antimicrobial peptide from the penaeid shrimp adopts an alpha-helical structure that specifically permeabilizes fungal membranes. Biochim. Et Biophys. Acta-Gen. Subj. 2016, 1860, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.A.; Cole, G.B.; Nguyen, H.Q.; Sharpe, S.; Deber, C.M. Influence of hydrocarbon-stapling on membrane interactions of synthetic antimicrobial peptides. Bioorganic Med. Chem. 2018, 26, 1189–1196. [Google Scholar] [CrossRef]
- Glibowicka, M.; He, S.; Deber, C.M. Enhanced proteolytic resistance of cationic antimicrobial peptides through lysine side chain analogs and cyclization. Biochem. Biophys. Res. Commun. 2022, 612, 105–109. [Google Scholar] [CrossRef]
- Silva, A.L.; Bezerra, L.P.; Freitas, C.D.T.; Silva, A.F.B.; Mesquita, F.P.; Neto, N.A.S.; Oliveira, J.P.B.; Aguiar, T.K.B.; Nagano, C.S.; Carneiro, R.F.; et al. Luffa operculata seed proteins: Identification by LC-ESI-MS/MS and biotechnological potential against Candida albicans and C. krusei. Anal. Biochem. 2022, 655, 114851. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Liu, X.; Zhong, K.; Huang, Y.; Gao, H. Antibacterial Activity of Shikimic Acid from Pine Needles of Cedrus deodara against Staphylococcus aureus through Damage to Cell Membrane. Int. J. Mol. Sci. 2015, 16, 27145–27155. [Google Scholar] [CrossRef]
- Ahmed, O.; Mohamed, H.; Salem, W.; Afifi, M.; Song, Y. Efficacy of Ethanolic Extract of Syzygium aromaticum in the Treatment of Multidrug-Resistant Pseudomonas aeruginosa Clinical Isolates Associated with Urinary Tract Infections. Evid.-Based Complement. Altern. Med. 2021, 2021, 6612058. [Google Scholar] [CrossRef]
- Bandara, H.M.H.N.; Wood, D.L.A.; Vanwonterghem, I.; Hugenholtz, P.; Cheung, B.P.K.; Samaranayake, L.P. Fluconazole resistance in Candida albicans is induced by Pseudomonas aeruginosa quorum sensing. Sci. Rep. 2020, 10, 7769. [Google Scholar] [CrossRef]
- Liu, T.T.; Lee, R.E.B.; Barker, K.S.; Lee, R.E.; Wei, L.; Homayouni, R.; Rogers, P.D. Genome-wide expression profiling of the response to azole, polyene, echinocandin, and pyrimidine antifungal agents in Candida albicans. Antimicrob. Agents Chemother. 2005, 49, 2226–2236. [Google Scholar] [CrossRef]
- Yang, L.-B.; Guo, G.; Tian, Z.-Q.; Zhou, L.-X.; Zhu, L.-J.; Peng, J.; Sun, C.-Q.; Huang, M.-J. TMT-based quantitative proteomic analysis of the effects of novel antimicrobial peptide AMP-17 against Candida albicans. J. Proteom. 2022, 250, 104385. [Google Scholar] [CrossRef]
- Kong, C.; Chee, C.-F.; Richter, K.; Thomas, N.; Abd Rahman, N.; Nathan, S. Suppression of Staphylococcus aureus biofilm formation and virulence by a benzimidazole derivative, UM-C162. Sci. Rep. 2018, 8, 2758. [Google Scholar] [CrossRef] [PubMed]
- Mulkern, A.J.; Oyama, L.B.; Cookson, A.R.; Creevey, C.J.; Wilkinson, T.J.; Olleik, H.; Maresca, M.; da Silva, G.C.; Fontes, P.P.; Bazzolli, D.M.S.; et al. Microbiome-derived antimicrobial peptides offer therapeutic solutions for the treatment of Pseudomonas aeruginosa infections. NPJ Biofilms Microbiomes 2022, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Pang, X.; Wu, Y.; Wu, Y.; Shi, Y.; Zhang, X.; Chen, Q. Synergistic Antibacterial Mechanism of Mannosylerythritol Lipid-A and Lactic Acid on Listeria monocytogenes Based on Transcriptomic Analysis. Foods 2022, 11, 2660. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhou, D.; Steitz, T.A.; Polikanov, Y.S.; Gagnon, M.G. Ribosome-Targeting Antibiotics: Modes of Action, Mechanisms of Resistance, and Implications for Drug Design. Annu. Rev. Biochem. 2018, 87, 451–478. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaeberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, H.; Roudbarmohammadi, S.; Delavari, H.H.; Roudbary, M. Antifungal effects of indolicidin-conjugated gold nanoparticles against fluconazole-resistant strains of Candida albicans isolated from patients with burn infection. Int. J. Nanomed. 2019, 14, 5323–5338. [Google Scholar] [CrossRef]
- Manoharlal, R.; Gaur, N.A.; Panwar, S.L.; Morschhaeuser, J.; Prasad, R. Transcriptional activation and increased mRNA stability contribute to overexpression of CDR1 in azole-resistant Candida albicans. Antimicrob. Agents Chemother. 2008, 52, 1481–1492. [Google Scholar] [CrossRef]
- Yang, S.-T.; Kim, J.I.; Shin, S.Y. Effect of dimerization of a beta-turn antimicrobial peptide, PST13-RK, on antimicrobial activity and mammalian cell toxicity. Biotechnol. Lett. 2009, 31, 233–237. [Google Scholar] [CrossRef]
- Murinda, S.E.; Rashid, K.A.; Roberts, R.F. In vitro assessment of the cytotoxicity of nisin, pediocin, and selected colicins on simian virus 40-transfected human colon and Vero monkey kidney cells with trypan blue staining viability assays. J. Food Prot. 2003, 66, 847–853. [Google Scholar] [CrossRef]
- Karthik, R.; Manigandan, V.; Ebenezar, K.K.; Kavitha, M.; Saravanan, R. Structural characterization, teratogenicity and in vitro avian antimicrobial activity of posterior salivary gland (PSG) toxin from cuttlefish, Sepia prashadi. Int. J. Biol. Macromol. 2019, 124, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Fialho, T.L.; Carrijo, L.C.; Magalhaes Junior, M.J.; Baracat-Pereira, M.C.; Piccoli, R.H.; de Abreu, L.R. Extraction and identification of antimicrobial peptides from the Canastra artisanal minas cheese. Food Res. Int. 2018, 107, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Vinod, V.; Jayakumar, J.; Parasmal, S.; Biswas, R. Antifungal activity of Syzygium samarangense leaf extracts against Candida. Lett. Appl. Microbiol. 2021, 73, 1. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, P.; Bidaud, A.-L.; Dannaoui, E. In vitro synergy of isavuconazole in combination with colistin against Candida auris. Sci. Rep. 2020, 10, 1. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial Activity and Mechanism of Action of Chlorogenic Acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Taggar, R.; Jangra, M.; Dwivedi, A.; Bansal, K.; Patil, P.B.; Bhattacharyya, M.S.; Nandanwar, H.; Sahoo, D.K. Bacteriocin isolated from the natural inhabitant of Allium cepa against Staphylococcus aureus. World J. Microbiol. Biotechnol. 2021, 37, 1–6. [Google Scholar] [CrossRef]
- Wang, Q.R.; Li, N.N.; Wang, Y.J.; Li, R.L.; Jia, Y.A.; Zhou, J.N.; Liu, J.Y.; Zhang, M.; Zhuang, P.W.; He, C.W.; et al. Studies on the key constituents and the related mechanisms of Clerodendranthus spicatus in the treatment of diabetes based on network pharmacology. J. Ethnopharmacol. 2023, 303, 115949. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, Y.; Zhou, J.; Wang, J.; Zhang, M.; Chen, H. Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.). Molecules 2023, 28, 3098. https://doi.org/10.3390/molecules28073098
Li S, Wang Y, Zhou J, Wang J, Zhang M, Chen H. Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.). Molecules. 2023; 28(7):3098. https://doi.org/10.3390/molecules28073098
Chicago/Turabian StyleLi, Shuqin, Yajie Wang, Jingna Zhou, Jia Wang, Min Zhang, and Haixia Chen. 2023. "Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.)" Molecules 28, no. 7: 3098. https://doi.org/10.3390/molecules28073098
APA StyleLi, S., Wang, Y., Zhou, J., Wang, J., Zhang, M., & Chen, H. (2023). Structural Characterization, Cytotoxicity, and the Antifungal Mechanism of a Novel Peptide Extracted from Garlic (Allium sativa L.). Molecules, 28(7), 3098. https://doi.org/10.3390/molecules28073098