

Marine Fish-Derived Lysophosphatidylcholine: Properties, Extraction, Quantification, and Brain Health Application

 ,
,  , , ,
, , , 
Abstract
1. Introduction
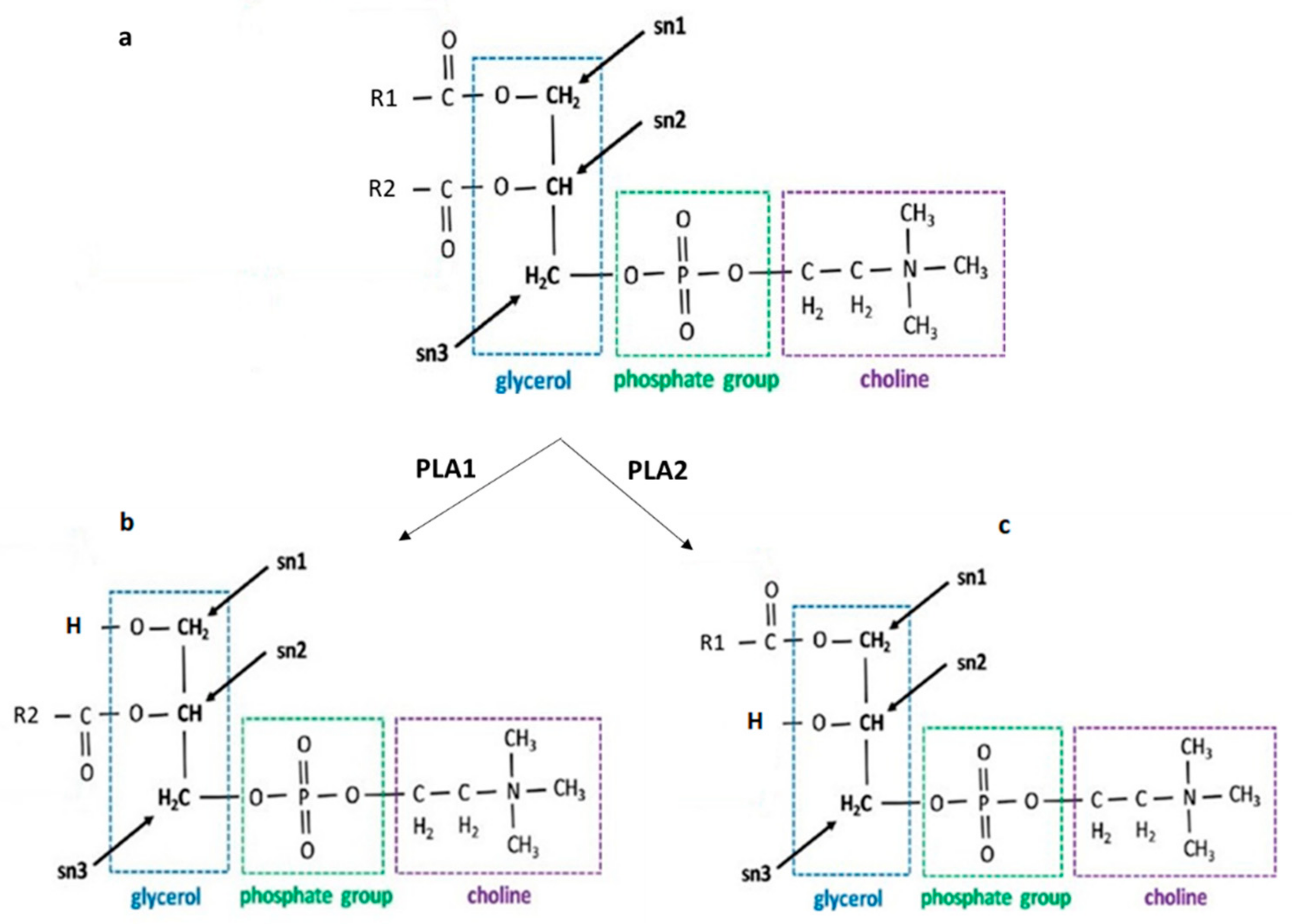
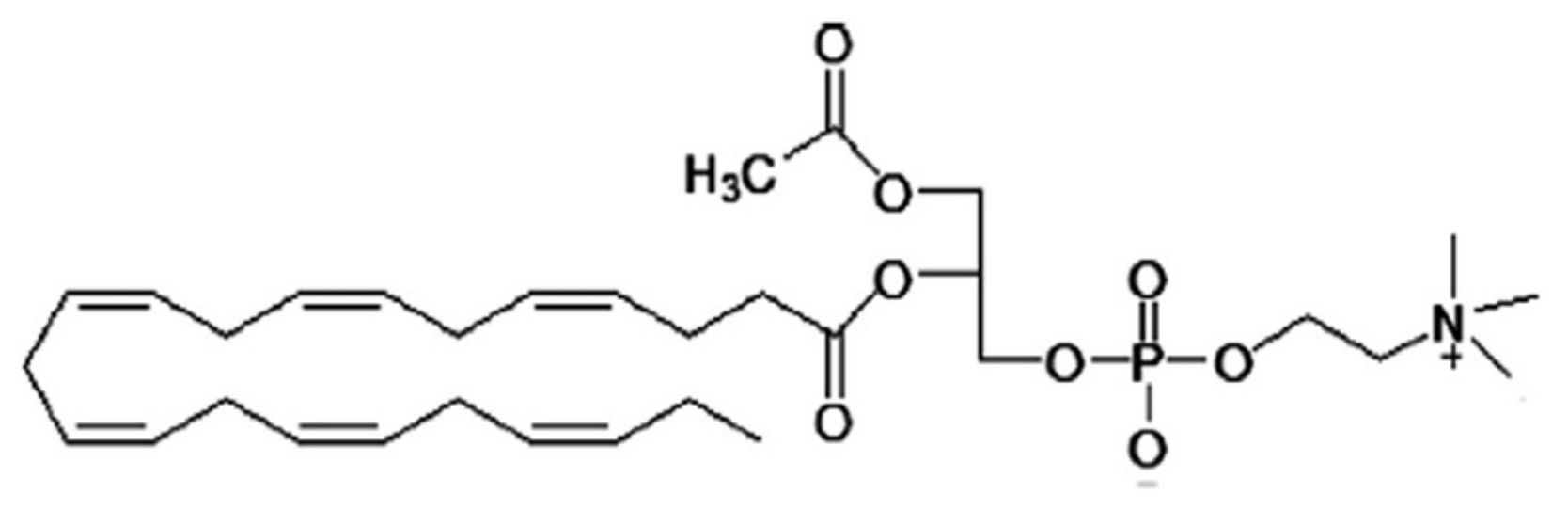
2. Structure and Biosynthesis of LPC
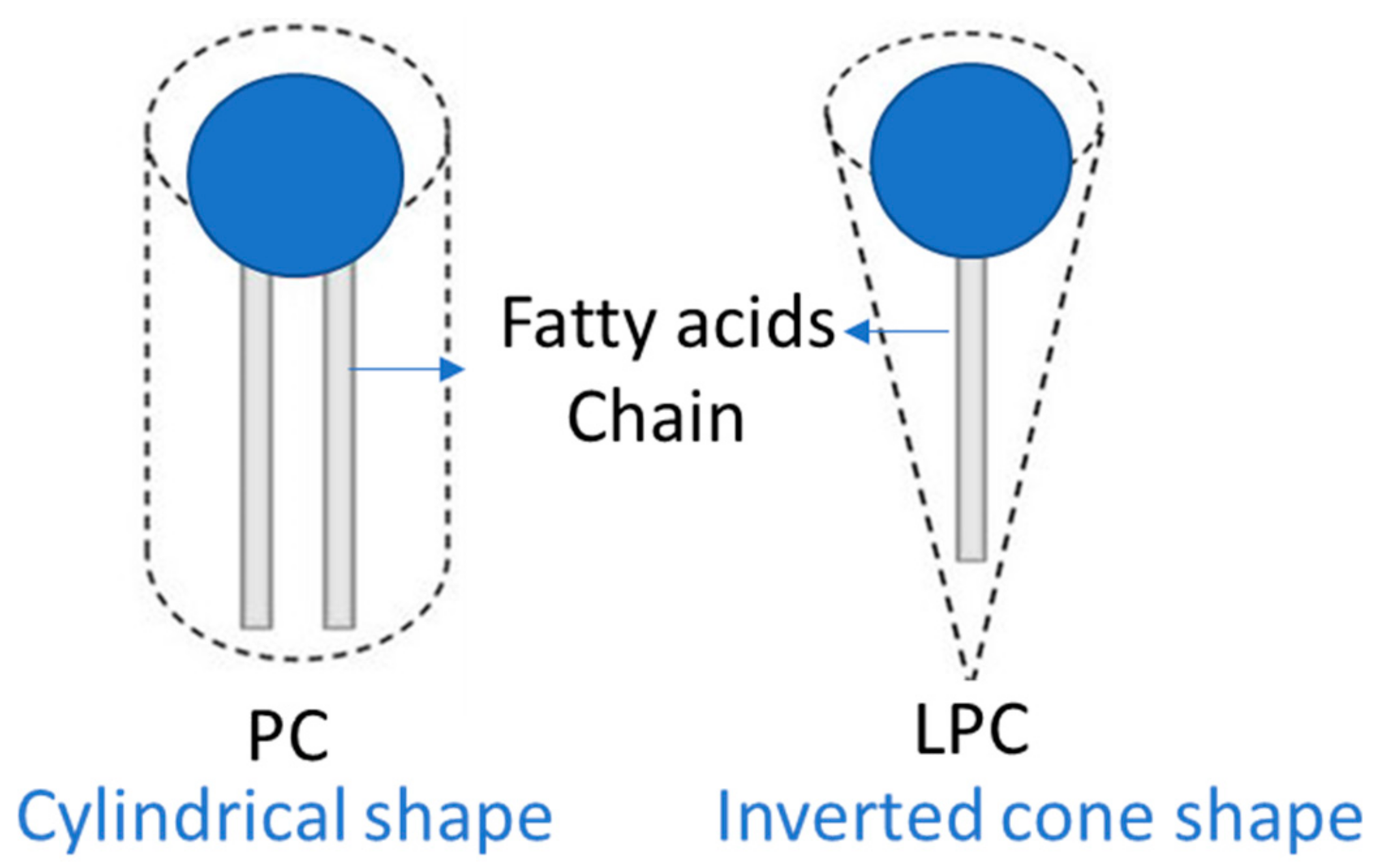
3. Properties of LPC
4. Marine Based LPC Compared to Terrestrial Based LPC
5. Factors Affecting Phospholipid Composition
6. Extraction and Quantification of LPC
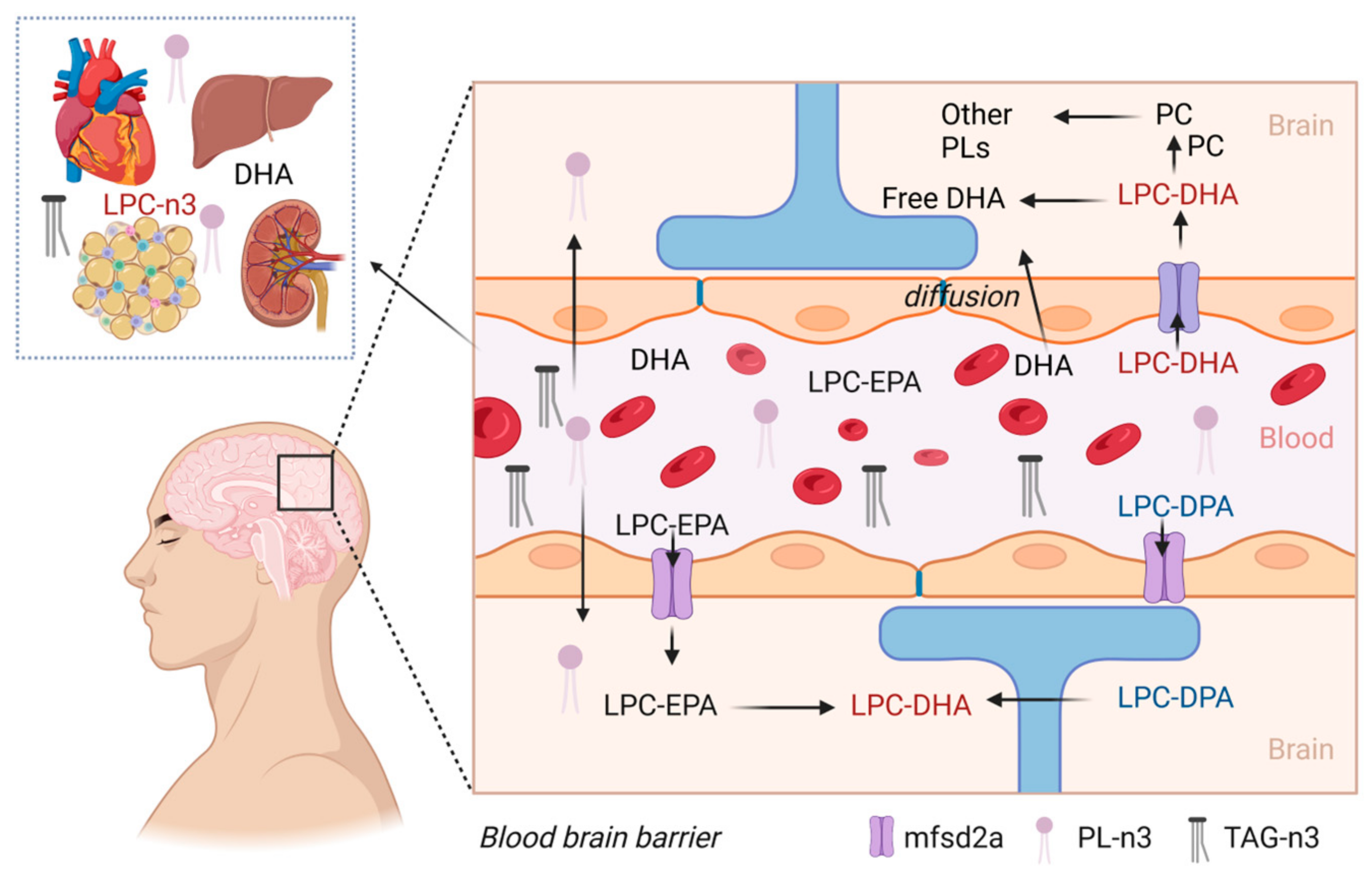
7. Neuroprotection and Brain Health Application of LPC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of PL Form | Model System | Treatment and Duration | Findings | References |
|---|---|---|---|---|
| 1-DHA-LPC 2-DHA-LPC | adult mice | 1 mg/d (oral administration); 1 month | -Increased the percentage of DHA and C20:4 in the plasma -Improved DHA level and C22:6/C20:4 ratio in all regions of the brain -LPC-DHA treatment remarkably improved memory and spatial learning -Increased molecular species of DHA-PC, DHA-PE in the plasma and BNDF level (brain derived neurotrophic factor) in all brain regions | [14] |
| sn-1 LPC-EPA | adult mice | 1 mg DHA per d or ∼40 mg/kg body weight | -Improved memory and cognition. -LPC-EPA in the brain increased from 0.03 to 4 μmol/g (>100-fold) -Little effect on free EPA -DHA was increased 2-fold by LPC-EPA but not by free EPA. -LPC-EPA also increased DHA concentration in the retina and improved brain-derived neurotrophic factors in the brain | [15] |
| Labelled LPC | adult squirrel monkeys | 1.36 μmol | -LPC in the plasma is taken up by the brain, metabolized in brain tissue and acts as a precursor of PC and choline | [96] |
| LPC-DHA | APOE3- and APOE4-TR mice | 4 to 12 months | -LPC-DHA-enriched krill oil could increase DHA level in the brain -Improved memory relevant behaviour -LPC-DHA supplements could be preventive for some level of age-related neurodegeneration | [16] |
| LPC | adult mice | - | -Revealed the protection against LPC-induced demyelinating lesion and cognitive deficits -Could be a feasible and promising therapeutic treatment against demyelinating diseases | [97] |
| LPC-DHA | expectant mice | DHA-LPC (68.8 mg/mL, oral administration); 4 d | -Improved the DHA level in the brain | [98] |
8. Role of LPC in Drug Delivery
9. LPC for the Delivery of Other Bioactive Ingredients
10. Safety of LPC
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, X.; Gross, R.W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrom. Rev. 2005, 24, 367–412. [Google Scholar] [CrossRef]
- Özdemir, N.Ş.; Parrish, C.C.; Parzanini, C.; Mercier, A. Neutral and polar lipid fatty acids in five families of demersal and pelagic fish from the deep Northwest Atlantic. ICES J. Mar. Sci. 2019, 76, 1807–1815. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [PubMed]
- El-Bacha, T.; Torres, A.G. Phospholipids: Physiology. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 352–359. [Google Scholar]
- Lu, F.S.; Nielsen, N.S.; Timm-Heinrich, M.; Jacobsen, C. Oxidative stability of marine phospholipids in the liposomal form and their applications. Lipids 2011, 46, 3–23. [Google Scholar] [PubMed]
- Xie, M. Phospholipids. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 214–217. [Google Scholar]
- Zhang, T.-T.; Xu, J.; Wang, Y.-M.; Xue, C.-H. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids. Prog. Lipid Res. 2019, 75, 100997. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.; Carne, A.; Bekhit, A.E.D. Marine omega-3 (n-3) phospholipids: A comprehensive review of their properties, sources, bioavailability, and relation to brain health. Compr. Rev. Food Sci. Food Saf. 2020, 19, 64–123. [Google Scholar] [CrossRef]
- Bickerton, A.S.; Roberts, R.; Fielding, B.A.; Hodson, L.; Blaak, E.E.; Wagenmakers, A.J.; Gilbert, M.; Karpe, F.; Frayn, K.N. Preferential uptake of dietary fatty acids in adipose tissue and muscle in the postprandial period. Diabetes 2007, 56, 168–176. [Google Scholar] [CrossRef]
- Murota, K. Digestion and absorption of dietary glycerophospholipids in the small intestine: Their significance as carrier molecules of choline and n-3 polyunsaturated fatty acids. Biocatal. Agric. Biotechnol. 2020, 26, 101633. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Zhang, X.; Wenk, M.R.; Goh, E.L.; Silver, D.L. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Hachem, M.; Belkouch, M.; Van, A.L.; Picq, M.; Bernoud-Hubac, N.; Lagarde, M. Brain targeting with docosahexaenoic acid as a prospective therapy for neurodegenerative diseases and its passage across blood brain barrier. Biochimie 2020, 170, 203–211. [Google Scholar] [CrossRef]
- Thies, F.; Pillon, C.; Moliere, P.; Lagarde, M.; Lecerf, J. Preferential incorporation of sn-2 lysoPC DHA over unesterified DHA in the young rat brain. Am. J. Physiol. 1994, 267 Pt 2, R1273–R1279. [Google Scholar] [CrossRef]
- Sugasini, D.; Yalagala, P.C.R.; Goggin, A.; Tai, L.M.; Subbaiah, P.V. Enrichment of brain docosahexaenoic acid (DHA) is highly dependent upon the molecular carrier of dietary DHA: Lysophosphatidylcholine is more efficient than either phosphatidylcholine or triacylglycerol. J. Nutr. Biochem. 2019, 74, 108231. [Google Scholar] [CrossRef] [PubMed]
- Yalagala, P.C.R.; Sugasini, D.; Dasarathi, S.; Pahan, K.; Subbaiah, P.V. Dietary lysophosphatidylcholine-EPA enriches both EPA and DHA in the brain: Potential treatment for depression. J. Lipid Res. 2019, 60, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Scheinman, S.B.; Sugasini, D.; Zayed, M.; Yalagala, P.C.R.; Marottoli, F.M.; Subbaiah, P.V.; Tai, L.M. LPC-DHA/EPA-Enriched Diets Increase Brain DHA and Modulate Behavior in Mice That Express Human APOE4. Front. Neurosci. 2021, 15, 690410. [Google Scholar] [CrossRef] [PubMed]
- Poumès-Ballihaut, C.; Langelier, B.; Houlier, F.; Alessandri, J.M.; Durand, G.; Latge, C.; Guesnet, P. Comparative bioavailability of dietary α-linolenic and docosahexaenoic acids in the growing rat. Lipids 2001, 36, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Kitson, A.P.; Metherel, A.H.; Chen, C.T.; Domenichiello, A.F.; Trépanier, M.-O.; Berger, A.; Bazinet, R.P. Effect of dietary docosahexaenoic acid (DHA) in phospholipids or triglycerides on brain DHA uptake and accretion. J. Nutr. Biochem. 2016, 33, 91–102. [Google Scholar] [CrossRef]
- van Hoogevest, P. Review–An update on the use of oral phospholipid excipients. Eur. J. Pharm. Sci. 2017, 108, 1–12. [Google Scholar] [CrossRef]
- Haq, M.; Suraiya, S.; Ahmed, S.; Chun, B.-S. Phospholipids from marine source: Extractions and forthcoming industrial applications. J. Funct. Foods 2021, 80, 104448. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Ahmmed, F.; Carne, A.; Tian, H.S.; Bekhit, A.E.-D.A. Fish roe phospholipids and health: Composition, extraction, storage and brain health application. In Fish Roe; Elsevier: Amsterdam, The Netherlands, 2022; pp. 93–142. [Google Scholar]
- Ferreira, I.; Rauter, A.P.; Bandarra, N.M. Marine Sources of DHA-Rich Phospholipids with Anti-Alzheimer Effect. Mar. Drugs 2022, 20, 662. [Google Scholar] [CrossRef]
- Tan, S.T.; Ramesh, T.; Toh, X.R.; Nguyen, L.N. Emerging roles of lysophospholipids in health and disease. Prog. Lipid Res. 2020, 80, 101068. [Google Scholar] [CrossRef]
- Hachem, M.; Nacir, H. Emerging Role of Phospholipids and Lysophospholipids for Improving Brain Docosahexaenoic Acid as Potential Preventive and Therapeutic Strategies for Neurological Diseases. Int. J. Mol. Sci. 2022, 23, 3969. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of lysophosphatidylcholine (LPC) in atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef]
- Law, S.-H.; Chan, M.-L.; Marathe, G.K.; Parveen, F.; Chen, C.-H.; Ke, L.-Y. An updated review of lysophosphatidylcholine metabolism in human diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed]
- Mnasri, T.; Hérault, J.; Gauvry, L.; Loiseau, C.; Poisson, L.; Ergan, F.; Pencréac’h, G. Lipase-catalyzed production of lysophospholipids. OCL 2017, 24, D405. [Google Scholar] [CrossRef]
- Thiés, F.; Delachambre, M.C.; Bentejac, M.; Lagarde, M.; Lecerf, J. Unsaturated fatty acids esterified in 2-acyl-l-lysophosphatidylcholine bound to albumin are more efficiently taken up by the young rat brain than the unesterified form. J. Neurochem. 1992, 59, 1110–1116. [Google Scholar] [CrossRef]
- Bernoud, N.; Fenart, L.; Molière, P.; Dehouck, M.P.; Lagarde, M.; Cecchelli, R.; Lecerf, J. Preferential transfer of 2-docosahexaenoyl-1-lysophosphatidylcholine through an in vitro blood-brain barrier over unesterified docosahexaenoic acid. J. Neurochem. 1999, 72, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Devos, M.; Poisson, L.; Ergan, F.; Pencreac’h, G. Enzymatic hydrolysis of phospholipids from Isochrysis galbana for docosahexaenoic acid enrichment. Enzym. Microb. Technol. 2006, 39, 548–554. [Google Scholar] [CrossRef]
- Poisson, L.; Devos, M.; Godet, S.; Ergan, F.; Pencreac’h, G. Acyl migration during deacylation of phospholipids rich in docosahexaenoic acid (DHA): An enzymatic approach for evidence and study. Biotechnol. Lett. 2009, 31, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Pencreac’h, G.; Ergan, F.; Poisson, L. Production of lysophospholipids rich in DHA. Lipid Technol. 2011, 23, 250–252. [Google Scholar] [CrossRef]
- Hachem, M.; Géloën, A.; Van, A.L.; Foumaux, B.; Fenart, L.; Gosselet, F.; Da Silva, P.; Breton, G.; Lagarde, M.; Picq, M.; et al. Efficient Docosahexaenoic Acid Uptake by the Brain from a Structured Phospholipid. Mol. Neurobiol. 2016, 53, 3205–3215. [Google Scholar] [CrossRef]
- Lagarde, M.; Hachem, M.; Picq, M.; Guichardant, M.; Bernoud-Hubac, N. AceDoPC, a structured phospholipid to target the brain with docosahexaenoic acid. OCL 2016, 23, D102. [Google Scholar] [CrossRef]
- Bernoud-Hubac, N.; Hachem, M.; Van, A.L.; Picq, M.; Lagarde, M. Specific Uptake of DHA by the Brain from a Structured Phospholipid, AceDoPC®; EDP Sciences: Les Ulis, France, 2017. [Google Scholar]
- Lagarde, M.; Hachem, M.; Bernoud-Hubac, N.; Picq, M.; Véricel, E.; Guichardant, M. Biological properties of a DHA-containing structured phospholipid (AceDoPC) to target the brain. Prostaglandins Leukot. Essent. Fat. Acids 2015, 92, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Subbaiah, P.V. Phospholipid and fatty acid specificity of endothelial lipase: Potential role of the enzyme in the delivery of docosahexaenoic acid (DHA) to tissues. Biochim. Biophys. Acta 2007, 1771, 1319–1328. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.-G. Lipase-catalyzed synthesis of lysophosphatidylcholine using organic cosolvent for in situ water activity control. J. Am. Oil Chem. Soc. 2000, 77, 791–797. [Google Scholar] [CrossRef]
- Virto, C.; Adlercreutz, P. Lysophosphatidylcholine synthesis with Candida antarctica lipase B (Novozym 435). Enzym. Microb. Technol. 2000, 26, 630–635. [Google Scholar] [CrossRef]
- Han, J.J.; Rhee, J.S. Effect of salt hydrate pairs for water activity control on lipase-catalyzed synthesis of lysophospholipids in a solvent-free system. Enzym. Microb. Technol. 1998, 22, 158–164. [Google Scholar] [CrossRef]
- Yang, G.; Yang, R.; Hu, J. Lysophosphatidylcholine synthesis by lipase-catalyzed ethanolysis. J. Oleo Sci. 2015, 64, 443–447. [Google Scholar] [CrossRef] [PubMed]
- D’Arrigo, P.; Servi, S. Synthesis of lysophospholipids. Molecules 2010, 15, 1354–1377. [Google Scholar] [CrossRef]
- Panganamala, R.; Horrocks, L.A.; Geer, J.C.; Cornwell, D.G. Positions of double bonds in the monounsaturated alk-1-enyl groups from the plasmalogens of human heart and brain. Chem. Phys. Lipids 1971, 6, 97–102. [Google Scholar] [CrossRef]
- Robinson, N.; Saunders, L. The physical properties of lysolecithin and its sols: Part I.—Solubilities, surface and interfacial tensions. J. Pharm. Pharmacol. 1958, 10, 384–391. [Google Scholar] [CrossRef]
- Aldrich, S. LPC Product Information. Available online: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/product/documents/321/842/l4129pis.pdf (accessed on 8 January 2023).
- Kapoor, V.; Zaharieva, M.M.; Berger, M.R. Erufosine Induces Autophagy and Apoptosis in Oral Squamous Cell Carcinoma: Role of the Akt–mTOR Signaling Pathway. In Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection, and Aging; Elsevier: Amsterdam, The Netherlands, 2014; pp. 229–245. [Google Scholar]
- Tsushima, T.; Matsubara, K.; Ohkubo, T.; Inoue, Y.; Takahashi, K. Docosahexaenoic-and eicosapentaenoic acid-bound lysophospholipids are more effective in suppressing angiogenesis than conjugated docosahexaenoic acid. J. Oleo Sci. 2012, 61, 427–432. [Google Scholar] [CrossRef]
- Mathew, S.; Raman, M.; Kalarikkathara Parameswaran, M.; Rajan, D.P. Assessment of nutritional quality of fish. In Fish and Fishery Products Analysis; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–62. [Google Scholar]
- Moradi, Y.; Bakar, J.; Motalebi, A.A.; Syed Muhamad, S.H.; Che Man, Y. A Review on Fish Lipid: Composition and Changes During Cooking Methods. J. Aquat. Food Prod. Technol. 2011, 20, 379–390. [Google Scholar] [CrossRef]
- Gowda, S.B.G.; Minami, Y.; Gowda, D.; Chiba, H.; Hui, S.-P. Detection and characterization of lipids in eleven species of fish by non-targeted liquid chromatography/mass spectrometry. Food Chem. 2022, 393, 133402. [Google Scholar] [CrossRef] [PubMed]
- Aksu, M.I.; Dogan, M.; Sirkecioglu, A.N. Changes in the Total Lipid, Neutral Lipid, Phospholipid and Fatty Acid Composition of Phospholipid Fractions during Pastırma Processing, a Dry-Cured Meat Product. Korean J. Food Sci. Anim. Resour. 2017, 37, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Malau-Aduli, A.E.; Siebert, B.D.; Bottema, C.D.; Pitchford, W.S. Breed comparison of the fatty acid composition of muscle phospholipids in Jersey and Limousin cattle. J. Anim. Sci. 1998, 76, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Oz, F.; Kotan, G. Effects of different cooking methods and fat levels on the formation of heterocyclic aromatic amines in various fishes. Food Control 2016, 67, 216–224. [Google Scholar] [CrossRef]
- Nanda, P.; Sahu, N. Analysis of lipid and lipid-fractions of some freshwater fishes and their inter-relationship. J. Indian Fish. Assoc. 2005, 32, 87–94. [Google Scholar]
- Drazen, J.C. Depth related trends in proximate composition of demersal fishes in the eastern North Pacific. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 203–219. [Google Scholar] [CrossRef]
- Keriko, J.; Chege, C.; Magu, M.; Githua, M. Fish lipid contents and classes of selected fish species found in Lake Naivasha (Kenya) and the fish feeding habits of the lake’s inhabitants. Afr. J. Pharm. Pharmacol. 2010, 4, 745–753. [Google Scholar]
- Boselli, E.; Pacetti, D.; Lucci, P.; Frega, N.G. Characterization of Phospholipid Molecular Species in the Edible Parts of Bony Fish and Shellfish. J. Agric. Food Chem. 2012, 60, 3234–3245. [Google Scholar] [CrossRef]
- Voronin, V.P.; Nemova, N.N.; Ruokolainen, T.R.; Artemenkov, D.V.; Rolskii, A.Y.; Orlov, A.M.; Murzina, S.A. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules 2021, 11, 704. [Google Scholar] [CrossRef] [PubMed]
- El-Sebaiy, L.A.; Metwalli, S.M. Changes in some chemical characteristics and lipid composition of salted fermented Bouri Fish muscle (Mugil cephalus). Food Chem. 1989, 31, 41–50. [Google Scholar] [CrossRef]
- Pacetti, D.; Lucci, P.; Mozzon, M.; Gagliardi, R.; Fiorini, D.; Frega, N.G. Influence of deep-fat frying process on phospholipid molecular species composition of Sardina pilchardus fillet. Food Control 2015, 48, 155–162. [Google Scholar] [CrossRef]
- Nomikos, T.; Karantonis, H.C.; Skarvelis, C.; Demopoulos, C.A.; Zabetakis, I. Antiatherogenic properties of lipid fractions of raw and fried fish. Food Chem. 2006, 96, 29–35. [Google Scholar] [CrossRef]
- Roldan, H.A.; Roura, S.I.; Montecchia, C.L.; Pérez Borla, O.; Crupkin, M. Lipid changes in frozen stored fillets from pre-and postspawned hake (Merluccius hubbsi Marini). J. Food Biochem. 2005, 29, 187–204. [Google Scholar] [CrossRef]
- Kaneniwa, M.; Yokoyama, M.; Murata, Y.; Kuwahara, R. Enzymatic Hydrolysis of Lipids in Muscle of Fish and Shellfish During Cold Storage; Shahidi, F., Spanier, A.M., Ho, C.-T., Braggins, T., Eds.; Quality of Fresh and Processed Foods: Boston, MA, USA; Springer: Boston, MA, USA, 2004; pp. 113–119. [Google Scholar]
- Takahashi, K.; Zama, K. Compositional Changes in Molecular Species of Fish Muscle Phosphatidylcholine during Frozen Storage. Res. Rep. Fac. Fish. Hokkaido Univ. 1986, 37, 80–84. [Google Scholar]
- Hanaoka, K.; Toyomizu, M. Acceleration of phospholipid decomposition in fish muscle by freezing. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 465–468. [Google Scholar] [CrossRef]
- Fauske, K.R.; Bernhard, A.; Fjære, E.; Myrmel, L.S.; Frøyland, L.; Kristiansen, K.; Liaset, B.; Madsen, L. Effects of Frozen Storage on Phospholipid Content in Atlantic Cod Fillets and the Influence on Diet-Induced Obesity in Mice. Nutrients 2018, 10, 695. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Jiang, Q.; Yang, F.; Wang, B. Phospholipid molecular species composition of Chinese traditional low-salt fermented fish inoculated with different starter cultures. Food Res. Int. 2018, 111, 87–96. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Carne, A.; Tian, H.S.; Bekhit, A.E.-D.A. The effect of pulsed electric fields on the extracted total lipid yield and the lipidomic profile of hoki roe. Food Chem. 2022, 384, 132476. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Carne, A.; Tian, H.S.; Bekhit, A.E.-D.A. Use of fungal and bacterial protease preparations to enhance extraction of lipid from fish roe: Effect on lipidomic profile of extracted oil. Food Chem. X 2022, 16, 100499. [Google Scholar] [CrossRef] [PubMed]
- Khiari, Z.; Ndagijimana, M.; Betti, M. Low molecular weight bioactive peptides derived from the enzymatic hydrolysis of collagen after isoelectric solubilization/precipitation process of turkey by-products. Poult. Sci. 2014, 93, 2347–2362. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, M.K.; Ahmmed, F.; Stewart, I.; Carne, A.; Tian, H.S.; Bekhit, A.E.-D.A. Omega-3 phospholipids in Pacific blue mackerel (Scomber australasicus) processing by-products. Food Chem. 2021, 353, 129451. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, M.K.; Bunga, S.; Stewart, I.; Tian, H.; Carne, A.; Bekhit, A.E.-D.A. Simple and efficient one-pot extraction method for phospholipidomic profiling of total oil and lecithin by phosphorus-31 nuclear magnetic resonance measurements. J. Agric. Food Chem. 2020, 68, 14286–14296. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Carne, A.; Ahmmed, F.; Stewart, I.; Tian, H.S.; Bekhit, A.E.-D.A. Positional distribution of fatty acids and phospholipid composition in King salmon (Oncorhynchus tshawytscha) head, roe and skin using nuclear magnetic resonance spectroscopy. Food Chem. 2021, 363, 130302. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- MacKenzie, A.; Vyssotski, M.; Nekrasov, E. Quantitative analysis of dairy phospholipids by 31P NMR. J. Am. Oil Chem. Soc. 2009, 86, 757–763. [Google Scholar] [CrossRef]
- Vyssotski, M.; Lagutin, K.; MacKenzie, A.; Mitchell, K.; Scott, D. Phospholipids of New Zealand edible brown algae. Lipids 2017, 52, 629–639. [Google Scholar] [CrossRef]
- Garcia, C.; Lutz, N.W.; Confort-Gouny, S.; Cozzone, P.J.; Armand, M.; Bernard, M. Phospholipid fingerprints of milk from different mammalians determined by 31P NMR: Towards specific interest in human health. Food Chem. 2012, 135, 1777–1783. [Google Scholar] [CrossRef]
- Park, S.-E.; Yu, H.-Y.; Ahn, S. Development and validation of a simple method to quantify contents of phospholipids in krill oil by fourier-transform infrared spectroscopy. Foods 2021, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Freire, M.; Moaddel, R.; Sun, K.; Fabbri, E.; Zhang, P.; Khadeer, M.; Salem, N., Jr.; Ferrucci, L.; Semba, R.D. Targeted Metabolomics Shows Low Plasma Lysophosphatidylcholine 18:2 Predicts Greater Decline of Gait Speed in Older Adults: The Baltimore Longitudinal Study of Aging. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Patrick, R.P. Role of phosphatidylcholine-DHA in preventing APOE4-associated Alzheimer’s disease. FASEB J. 2019, 33, 1554–1564. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D. Perspective: The Potential Role of Circulating Lysophosphatidylcholine in Neuroprotection against Alzheimer Disease. Adv. Nutr. 2020, 11, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Belkouch, M.; Hachem, M.; Elgot, A.; Van, A.L.; Picq, M.; Guichardant, M.; Lagarde, M.; Bernoud-Hubac, N. The pleiotropic effects of omega-3 docosahexaenoic acid on the hallmarks of Alzheimer’s disease. J. Nutr. Biochem. 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Van, A.L.; Sakayori, N.; Hachem, M.; Belkouch, M.; Picq, M.; Lagarde, M.; Osumi, N.; Bernoud-Hubac, N. Mechanisms of DHA transport to the brain and potential therapy to neurodegenerative diseases. Biochimie 2016, 130, 163–167. [Google Scholar] [CrossRef]
- Nishikimi, M.; Shoaib, M.; Choudhary, R.C.; Aoki, T.; Miyara, S.J.; Yagi, T.; Hayashida, K.; Takegawa, R.; Yin, T.; Becker, L.B.; et al. Preserving Brain LPC-DHA by Plasma Supplementation Attenuates Brain Injury after Cardiac Arrest. Ann. Neurol. 2022, 91, 389–403. [Google Scholar] [CrossRef]
- Chauveau, F.; Cho, T.H.; Perez, M.; Guichardant, M.; Riou, A.; Aguettaz, P.; Picq, M.; Lagarde, M.; Berthezène, Y.; Nighoghossian, N.; et al. Brain-targeting form of docosahexaenoic acid for experimental stroke treatment: MRI evaluation and anti-oxidant impact. Curr. Neurovascular Res. 2011, 8, 95–102. [Google Scholar] [CrossRef]
- Knuplez, E.; Marsche, G. An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. Int. J. Mol. Sci. 2020, 21, 4501. [Google Scholar] [CrossRef]
- Engel, K.M.; Schiller, J.; Galuska, C.E.; Fuchs, B. Phospholipases and Reactive Oxygen Species Derived Lipid Biomarkers in Healthy and Diseased Humans and Animals—A Focus on Lysophosphatidylcholine. Front. Physiol. 2021, 12, 732319. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, J.; Liu, Y.; Wu, R.; Yang, H.; Hu, X.; Ling, X. Studies on diagnostic biomarkers and therapeutic mechanism of Alzheimer’s disease through metabolomics and hippocampal proteomics. Eur. J. Pharm. Sci. 2017, 105, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Mulder, C.; Wahlund, L.O.; Teerlink, T.; Blomberg, M.; Veerhuis, R.; van Kamp, G.J.; Scheltens, P.; Scheffer, P.G. Decreased lysophosphatidylcholine/phosphatidylcholine ratio in cerebrospinal fluid in Alzheimer’s disease. J. Neural Transm. 2003, 110, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.O.; Grösgen, S.; Riemenschneider, M.; Tanila, H.; Grimm, H.S.; Hartmann, T. From brain to food: Analysis of phosphatidylcholins, lyso-phosphatidylcholins and phosphatidylcholin-plasmalogens derivates in Alzheimer’s disease human post mortem brains and mice model via mass spectrometry. J. Chromatogr. A 2011, 1218, 7713–7722. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J.; Shih, C.; Cashman, J.R. Docosahexaenoic acid is a strong inhibitor of prostaglandin but not leukotriene biosynthesis. Proc. Natl. Acad. Sci. USA 1983, 80, 3581–3584. [Google Scholar] [CrossRef]
- Lukiw, W.J.; Cui, J.G.; Marcheselli, V.L.; Bodker, M.; Botkjaer, A.; Gotlinger, K.; Serhan, C.N.; Bazan, N.G. A role for docosahexaenoic acid-derived neuroprotectin D1 in neural cell survival and Alzheimer disease. J. Clin. Investig. 2005, 115, 2774–2783. [Google Scholar] [CrossRef]
- Bazan, N.G. The onset of brain injury and neurodegeneration triggers the synthesis of docosanoid neuroprotective signaling. Cell. Mol. Neurobiol. 2006, 26, 901–913. [Google Scholar] [CrossRef]
- Chen, P.; Véricel, E.; Lagarde, M.; Guichardant, M. Poxytrins, a class of oxygenated products from polyunsaturated fatty acids, potently inhibit blood platelet aggregation. FASEB J. 2011, 25, 382–388. [Google Scholar] [CrossRef]
- Illingworth, D.R.; Portman, O.W. The uptake and metabolism of plasma lysophosphatidylcholine in vivo by the brain of squirrel monkeys. Biochem. J. 1972, 130, 557–567. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, X.; Li, Y.; Chen, S.; Liu, S.; Yu, Z.; Wang, W. Transforming growth factor-β1 protects against LPC-induced cognitive deficit by attenuating pyroptosis of microglia via NF-κB/ERK1/2 pathways. J. Neuroinflamm. 2022, 19, 1–20. [Google Scholar] [CrossRef]
- Tsushima, T.; Ohkubo, T.; Onoyama, K.; Linder, M.; Takahashi, K. Lysophosphatidylserine form DHA maybe the most effective as substrate for brain DHA accretion. Biocatal. Agric. Biotechnol. 2014, 3, 303–309. [Google Scholar] [CrossRef]
- Hachem, M.; Nacir, H.; Picq, M.; Belkouch, M.; Bernoud-Hubac, N.; Windust, A.; Meiller, L.; Sauvinet, V.; Feugier, N.; Lambert-Porcheron, S.; et al. Docosahexaenoic Acid (DHA) Bioavailability in Humans after Oral Intake of DHA-Containing Triacylglycerol or the Structured Phospholipid AceDoPC(®). Nutrients 2020, 12, 251. [Google Scholar] [CrossRef] [PubMed]
- Sovic, A.; Panzenboeck, U.; Wintersperger, A.; Kratzer, I.; Hammer, A.; Levak-Frank, S.; Frank, S.; Rader, D.J.; Malle, E.; Sattler, W. Regulated expression of endothelial lipase by porcine brain capillary endothelial cells constituting the blood-brain barrier. J. Neurochem. 2005, 94, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Alakbarzade, V.; Hameed, A.; Quek, D.Q.; Chioza, B.A.; Baple, E.L.; Cazenave-Gassiot, A.; Nguyen, L.N.; Wenk, M.R.; Ahmad, A.Q.; Sreekantan-Nair, A.; et al. A partially inactivating mutation in the sodium-dependent lysophosphatidylcholine transporter MFSD2A causes a non-lethal microcephaly syndrome. Nat. Genet. 2015, 47, 814–817. [Google Scholar] [CrossRef]
- Chan, J.P.; Wong, B.H.; Chin, C.F.; Galam, D.L.A.; Foo, J.C.; Wong, L.C.; Ghosh, S.; Wenk, M.R.; Cazenave-Gassiot, A.; Silver, D.L. The lysolipid transporter Mfsd2a regulates lipogenesis in the developing brain. PLoS Biol. 2018, 16, e2006443. [Google Scholar] [CrossRef]
- Alqaraghuli, H.G.J.; Kashanian, S.; Rafipour, R. A review on targeting nanoparticles for breast cancer. Curr. Pharm. Biotechnol. 2019, 20, 1087–1107. [Google Scholar] [CrossRef]
- Khan, A.R.; Liu, M.; Khan, M.W.; Zhai, G. Progress in brain targeting drug delivery system by nasal route. J. Control. Release 2017, 268, 364–389. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Yang, Q.; Cai, N.; Zhang, Z. A cocktail of betulinic acid, parthenolide, honokiol and ginsenoside Rh2 in liposome systems for lung cancer treatment. Nanomedicine 2020, 15, 41–54. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.; Li, W.; Li, H.; Bao, J.; Yang, C.; Wang, A.; Wei, J.; Chen, S.; Jin, H. Role of Nrf2 in the antioxidation and oxidative stress induced developmental toxicity of honokiol in zebrafish. Toxicol. Appl. Pharmacol. 2019, 373, 48–61. [Google Scholar] [CrossRef]
- Zhou, H.; Zheng, B.; McClements, D.J. Encapsulation of lipophilic polyphenols in plant-based nanoemulsions: Impact of carrier oil on lipid digestion and curcumin, resveratrol and quercetin bioaccessibility. Food Funct. 2021, 12, 3420–3432. [Google Scholar] [CrossRef]
- Ralston, E.; Blumenthal, R.; Weinstein, J.N.; Sharrow, S.O.; Henkart, P. Lysophosphatidylcholine in liposomal membranes. Enhanced permeability but little effect on transfer of a water-soluble fluorescent marker into human lymphocytes. Biochim. Biophys. Acta (BBA)-Biomembr. 1980, 597, 543–551. [Google Scholar] [CrossRef]
- Ghosh, A.; Banik, S.; Suzuki, Y.; Mibe, Y.; Rikimura, S.; Komamoto, T.; Kuromi, K.; Yamada, K.; Sato, H.; Onoue, S. Lysophosphatidylcholine-based liposome to improve oral absorption and nephroprotective effects of astaxanthin. J. Sci. Food Agric. 2022, 103, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhan, Q.; Yi, K.; Chen, N.; Li, X.; Yang, S.; Hou, X.; Zhao, J.; Yuan, X.; Kang, C. Engineering Lipusu with lysophosphatidylcholine for improved tumor cellular uptake and anticancer efficacy. J. Mater. Chem. B 2022, 10, 1833–1842. [Google Scholar] [CrossRef]
- Chávez-Zamudio, R.; Ochoa-Flores, A.A.; Soto-Rodríguez, I.; Garcia-Varela, R.; García, H.S. Preparation, characterization and bioavailability by oral administration of O/W curcumin nanoemulsions stabilized with lysophosphatidylcholine. Food Funct. 2017, 8, 3346–3354. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Scalia, R.; Lefer, A.M. Lysophosphatidylcholine promotes P-selectin expression in platelets and endothelial cells: Possible involvement of protein kinase C activation and its inhibition by nitric oxide donors. Circ. Res. 1996, 78, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, K.E.; Andersson, L.; Nilsson, J.; Björkbacka, H. Nanomolar concentrations of lysophosphatidylcholine recruit monocytes and induce pro-inflammatory cytokine production in macrophages. Biochem. Biophys. Res. Commun. 2008, 370, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.; Rani, M.S.; Gerber, C.E.; Mukhopadhyay, C.; Ransohoff, R.M.; Chisolm, G.M.; Kottke-Marchant, K. Lysophosphatidylcholine regulates human microvascular endothelial cell expression of chemokines. J. Mol. Cell. Cardiol. 2003, 35, 1375–1384. [Google Scholar] [CrossRef]
- Kougias, P.; Chai, H.; Lin, P.H.; Lumsden, A.B.; Yao, Q.; Chen, C. Lysophosphatidylcholine and secretory phospholipase A2 in vascular disease: Mediators of endothelial dysfunction and atherosclerosis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2005, 12, RA5–RA16. [Google Scholar]
- Kim, E.A.; Ae Kim, J.; Park, M.H.; Jung, S.C.; Suh, S.H.; Pang, M.-G.; Kim, Y.J. Lysophosphatidylcholine induces endothelial cell injury by nitric oxide production through oxidative stress. J. Matern.-Fetal Neonatal Med. 2009, 22, 325–331. [Google Scholar] [CrossRef]
- Schmitz, G.; Ruebsaamen, K. Metabolism and atherogenic disease association of lysophosphatidylcholine. Atherosclerosis 2010, 208, 10–18. [Google Scholar] [CrossRef]
- Chaudhuri, P.; Colles, S.M.; Damron, D.S.; Graham, L.M. Lysophosphatidylcholine inhibits endothelial cell migration by increasing intracellular calcium and activating calpain. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 218–223. [Google Scholar] [CrossRef]
- Li, B.; Tian, S.; Liu, X.; He, C.; Ding, Z.; Shan, Y. Sulforaphane protected the injury of human vascular endothelial cell induced by LPC through up-regulating endogenous antioxidants and phase II enzymes. Food Funct. 2015, 6, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liang, Y.; Song, F.; Xu, S.; Nian, L.; Zhou, X.; Wang, S. TSG attenuates LPC-induced endothelial cells inflammatory damage through notch signaling inhibition. IUBMB Life 2016, 68, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, A.; Tokunaga, M.; Yamamoto, T.; Inoue, S.; Obama, K.; Kawahara, K.; Nakayama, H. Oxidized LDL and lysophosphatidylcholine stimulate plasminogen activator inhibitor-1 expression through reactive oxygen species generation and ERK1/2 activation in 3T3-L1 adipocytes. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2011, 1811, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Kumagai, T.; Ebina, K. A Synthetic Biotinylated Peptide, BP 21, Inhibits the Induction of mRNA Expression of Inflammatory Substances by Oxidized-and Lyso-Phosphatidylcholine. Drug Dev. Res. 2014, 75, 246–256. [Google Scholar] [CrossRef]
- Hasegawa, H.; Lei, J.; Matsumoto, T.; Onishi, S.; Suemori, K.; Yasukawa, M. Lysophosphatidylcholine enhances the suppressive function of human naturally occurring regulatory T cells through TGF-β production. Biochem. Biophys. Res. Commun. 2011, 415, 526–531. [Google Scholar] [CrossRef]
- Qin, X.; Qiu, C.; Zhao, L. Lysophosphatidylcholine perpetuates macrophage polarization toward classically activated phenotype in inflammation. Cell. Immunol. 2014, 289, 185–190. [Google Scholar] [CrossRef]
- Huang, Y.; Schäfer-Elinder, L.; Wu, R.; Claesson, H.; Frostegård, J. Lysophosphatidylcholine (LPC) induces proinflammatory cytokines by a platelet-activating factor (PAF) receptor-dependent mechanism. Clin. Exp. Immunol. 1999, 116, 326–331. [Google Scholar] [CrossRef]
- Yamamoto, N.; Homma, S.; Millman, I. Identification of the serum factor required for in vitro activation of macrophages. Role of vitamin D3-binding protein (group specific component, Gc) in lysophospholipid activation of mouse peritoneal macrophages. J. Immunol. 1991, 147, 273–280. [Google Scholar] [CrossRef]
- Dial, E.J.; Tran, D.M.; Romero, J.J.; Zayat, M.; Lichtenberger, L.M. A direct role for secretory phospholipase A2 and lysophosphatidylcholine in the mediation of LPS-induced gastric injury. Shock 2010, 33, 634–638. [Google Scholar] [CrossRef]




| Study | Objective | Method | Result |
|---|---|---|---|
| Thiés et al. [13] | To investigate the efficacy of 2-acyl-LPC linked to plasma albumin as a carrier of unsaturated FAs to the brain | Perfusion of 20-day-old rats with labeled 14C fatty acids in either non-esterified or esterified form in LPC labeled with 3H on the choline and 14C fatty acid groups | 2-acyl-LPC was an efficient delivery form of unsaturated FAs to the rat brain, suggesting that the FA delivery form could control their fate in tissues |
| Thiés et al. [13] | To compare the uptake and metabolism of 3H-DHA esterified at the sn-2 position of LPC-DHA and in a non-esterified form in twenty-day-old rats | Perfusion of 100 µL of lipid solution with LPC containing 1 µCi 3H-DHA (12 nmol) into the tail vein for 30 s | 3H-LPC-DHA was favourably improved in th e brain (4–5% of injected radioactivity) over the non-esterified form of DHA (0.3–0.4%), suggesting that the developing brain favorably employed sn-2-LPC-DHA compared to unesterified DHA |
| Sugasini et al. [14] | To determine the suitability of different sources of DHA supplementation in enhancing brain DHA levels | Feeding 20-day-old rats with DHA from triacylglycerol (TAG), natural PC, or LPC for four weeks | LPC (at sn-1 or sn-2 position) efficiently enhanced the DHA level in the brain, while DHA from TAG or natural PC (where DHA is esterified at sn-2 position) was not suitable for enhancing brain DHA content |
| Hachem et al. [33,99] | To examine the cerebral accretion of AceDoPC®, a stabilized form of LPC-DHA, in comparison to other forms of DHA | In vitro and in vivo studies investigating the passage of non-esterified 14C-DHA, 14C-PC-DHA, and 14C-AceDoPC® across a model of a blood–brain barrier (BBB) and the effects of the same forms of DHA injected into a twenty-day-old rat tail vein, respectively | AceDoPC® was a privileged and precise carrier of DHA to the brain, not in the other studied organs, when compared with DHA-containing PC and non-esterified DHA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmmed, M.K.; Hachem, M.; Ahmmed, F.; Rashidinejad, A.; Oz, F.; Bekhit, A.A.; Carne, A.; Bekhit, A.E.-D.A. Marine Fish-Derived Lysophosphatidylcholine: Properties, Extraction, Quantification, and Brain Health Application. Molecules 2023, 28, 3088. https://doi.org/10.3390/molecules28073088
Ahmmed MK, Hachem M, Ahmmed F, Rashidinejad A, Oz F, Bekhit AA, Carne A, Bekhit AE-DA. Marine Fish-Derived Lysophosphatidylcholine: Properties, Extraction, Quantification, and Brain Health Application. Molecules. 2023; 28(7):3088. https://doi.org/10.3390/molecules28073088
Chicago/Turabian StyleAhmmed, Mirja Kaizer, Mayssa Hachem, Fatema Ahmmed, Ali Rashidinejad, Fatih Oz, Adnan A. Bekhit, Alan Carne, and Alaa El-Din A. Bekhit. 2023. "Marine Fish-Derived Lysophosphatidylcholine: Properties, Extraction, Quantification, and Brain Health Application" Molecules 28, no. 7: 3088. https://doi.org/10.3390/molecules28073088
APA StyleAhmmed, M. K., Hachem, M., Ahmmed, F., Rashidinejad, A., Oz, F., Bekhit, A. A., Carne, A., & Bekhit, A. E.-D. A. (2023). Marine Fish-Derived Lysophosphatidylcholine: Properties, Extraction, Quantification, and Brain Health Application. Molecules, 28(7), 3088. https://doi.org/10.3390/molecules28073088