
Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts

Abstract
1. Introduction
2. Results
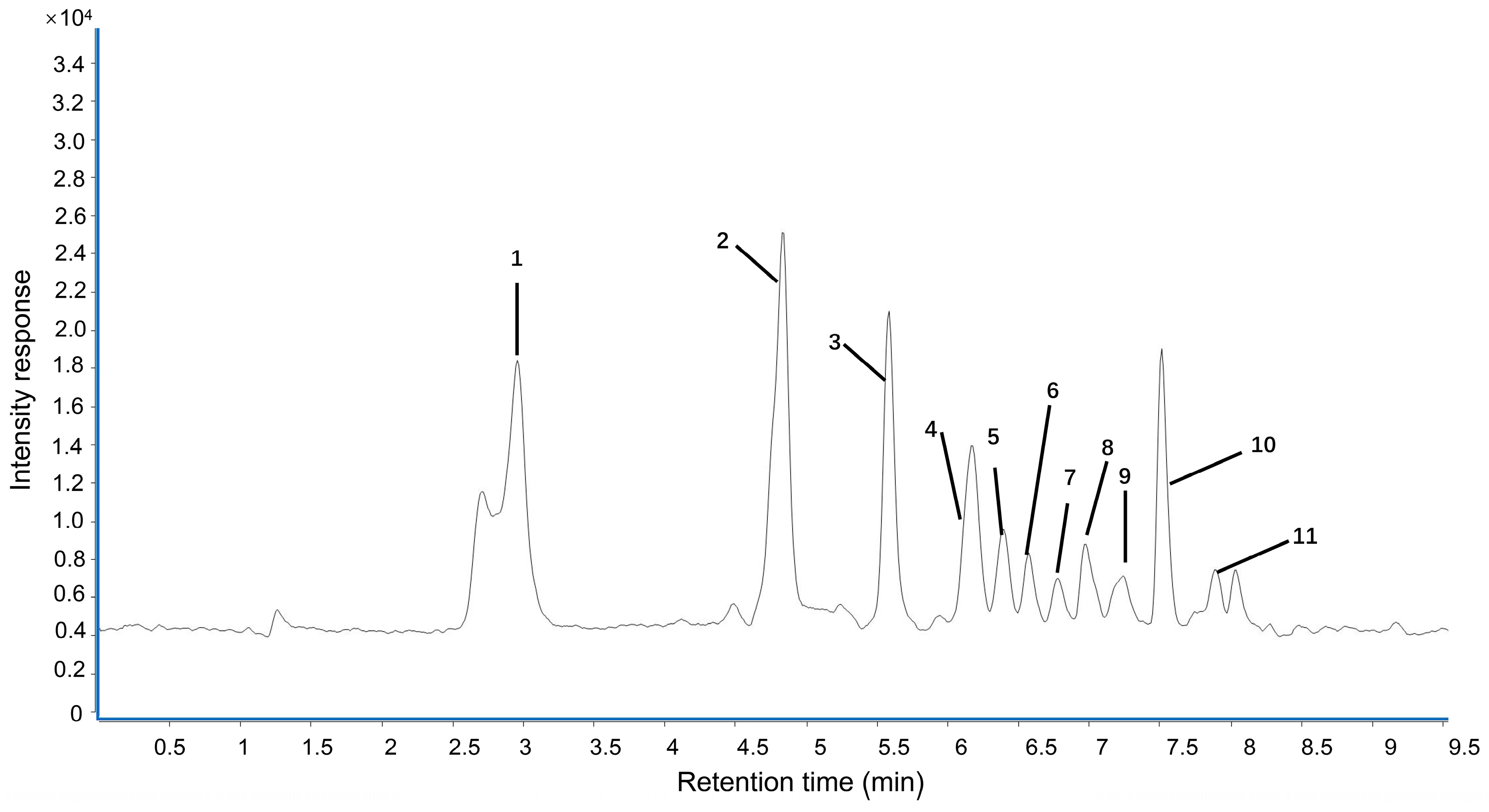
2.1. Polyphenol Contents and Antioxidant Activity of LPPE
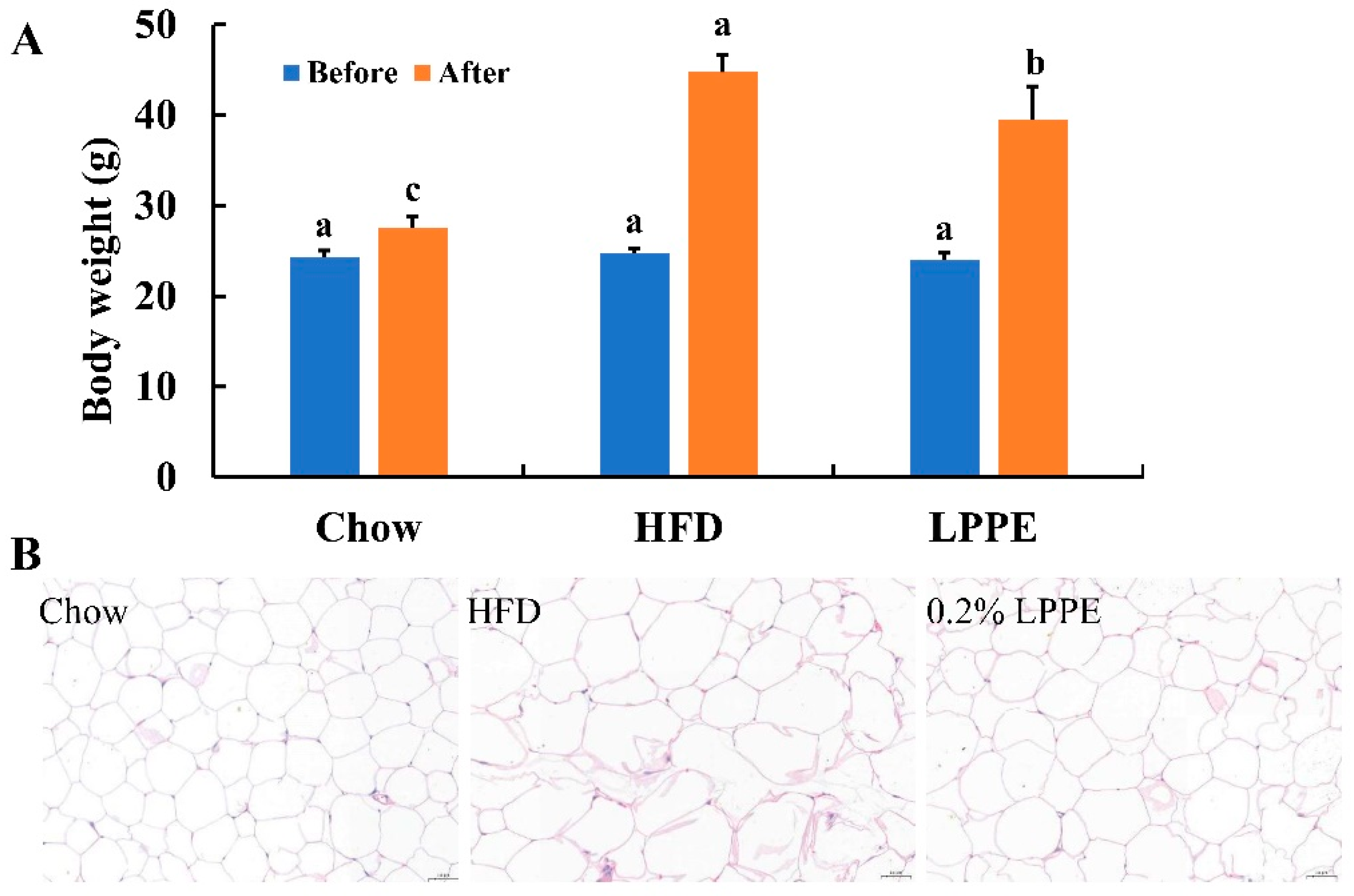
2.2. Effects of LPPE on Body Weight and Cell Size of Epididymal Adipose Tissues in High-Fat Diet-Induced Obese Mice
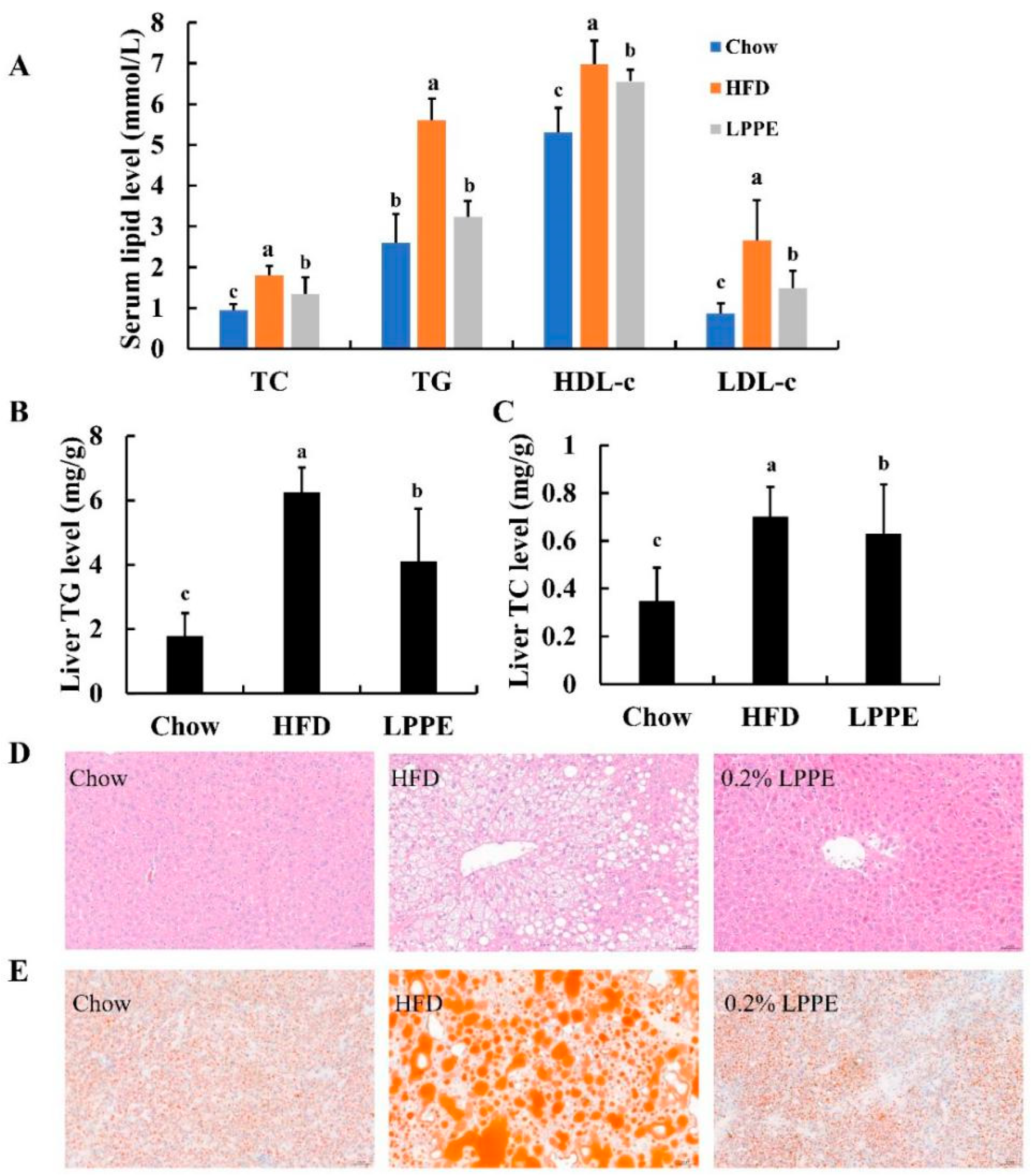
2.3. Effects of LPPE on Serum and Liver Lipids, and Hepatic Steatosis in High-Fat Diet-Induced Obese Mice
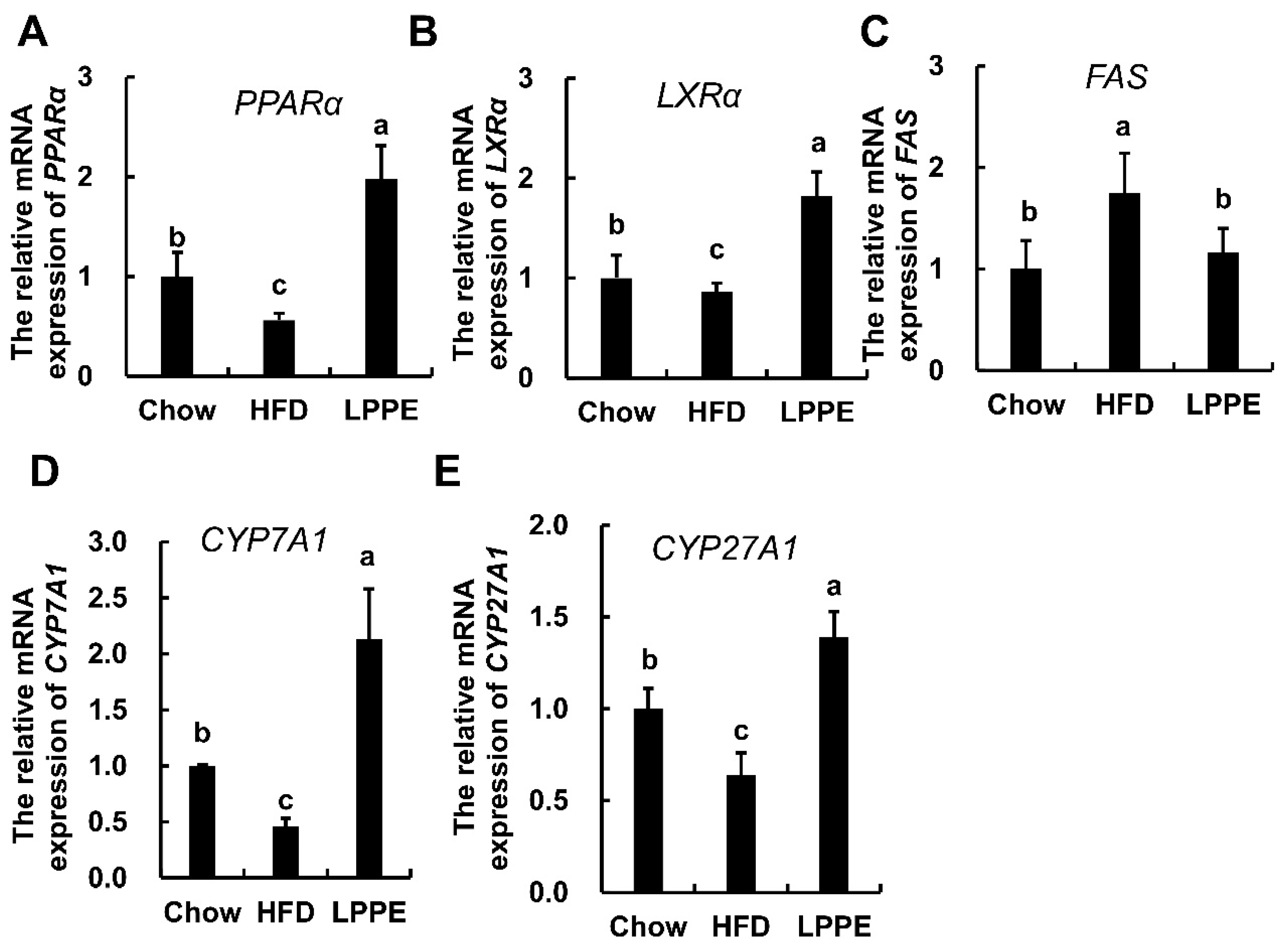
2.4. Effects of LPPE on the Gene Expression Involved in Lipid Metabolism
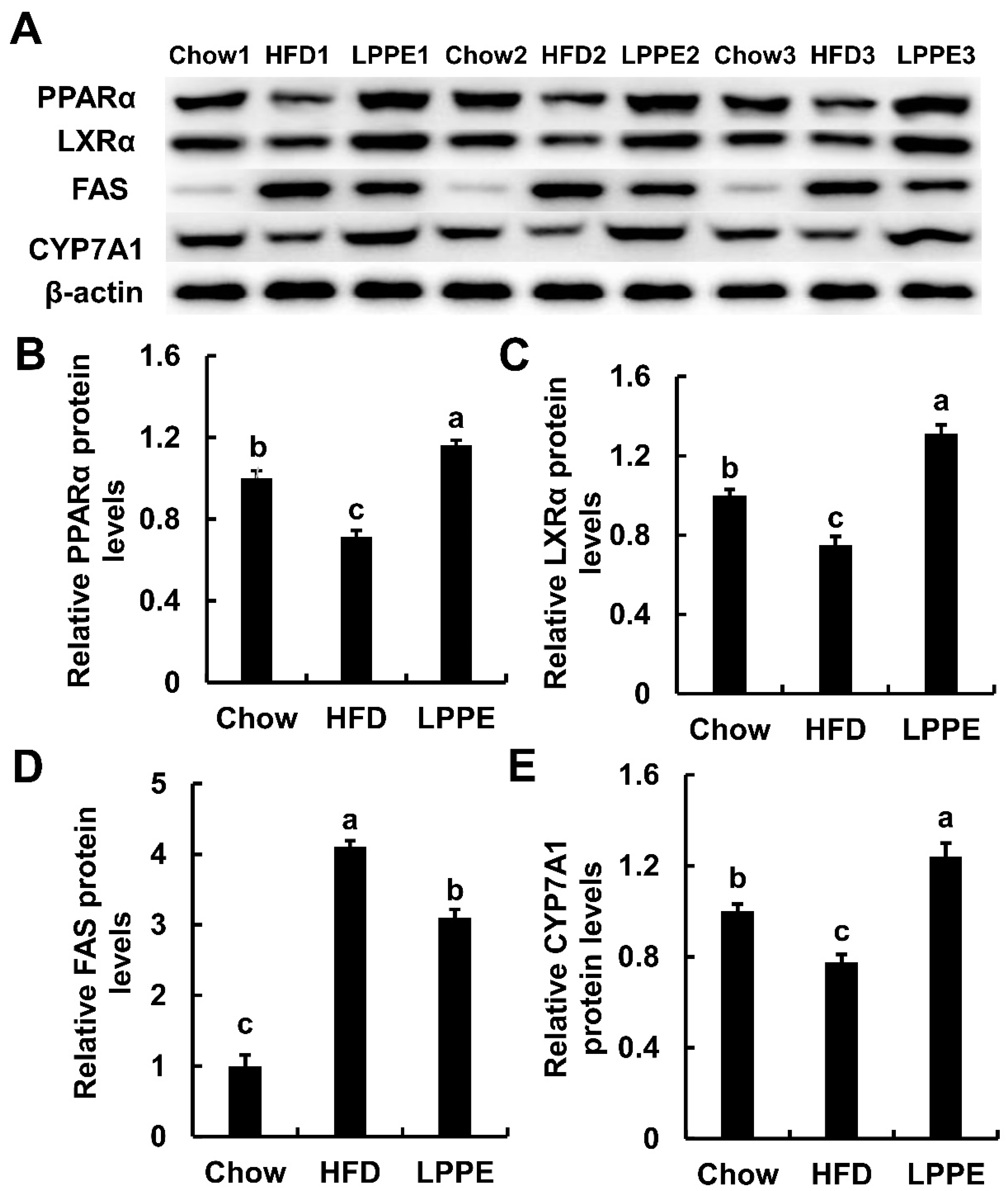
2.5. Effects of LPPE on the Protein Expressions Involved in Lipid Metabolism
3. Discussion
4. Materials and Methods
4.1. Reagents and Standards
4.2. Polyphenol Extracting
4.3. Identification and Quantification of Polyphenols via UPLC-QqQ-MS
4.4. Total Polyphenols and Antioxidant Activities In Vitro
4.5. Animal Experiment
4.6. Histological Analysis
4.7. Serum Lipids
4.8. Liver TG and TC Analysis
4.9. RT-PCR
4.10. Western Blot Assay
4.11. Statistics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Zhang, R.; Khan, S.A.; Lin, Y.; Guo, D.; Pan, X.; Liu, L.; Wei, Z.; Zhang, Y.; Deng, Y.; Zhang, M. Phenolic profiles and cellular antioxidant activity of longan pulp of 24 representative Chinese cultivars. Int. J. Food Prop. 2018, 21, 746–759. [Google Scholar] [CrossRef]
- Li, L.; Xu, J.; Mu, Y.; Han, L.; Liu, R.; Cai, Y.; Huang, X. Chemical characterization and anti-hyperglycaemic effects of polyphenol enriched longan (Dimocarpus longan Lour.) pericarp extracts. J. Funct. Foods 2015, 13, 314–322. [Google Scholar] [CrossRef]
- Zheng, G.; Xu, L.; Wu, P.; Xie, H.; Jiang, Y.; Chen, F.; Wei, X. Polyphenols from longan seeds and their radical-scavenging activity. Food Chem. 2009, 116, 433–436. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, S.; Ho, C.-T.; Bai, N. Phytochemical constituents and biological activities of longan (Dimocarpus longan Lour.) fruit: A review. Food Sci. Hum. Wellness 2020, 9, 95–102. [Google Scholar] [CrossRef]
- Rakariyatham, K.; Zhou, D.; Rakariyatham, N.; Shahidi, F. Sapindaceae (Dimocarpus longan and Nephelium lappaceum) seed and peel by-products: Potential sources for phenolic compounds and use as functional ingredients in food and health applications. J. Funct. Foods 2020, 67, 103846. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, K.; Li, W.; Soteyome, T.; Xiao, H.; Hu, Z. Protective effects of polyphenolic extracts from longan seeds promote healing of deep second-degree burn in mice. Food Funct. 2019, 10, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Rangkadilok, N.; Sitthimonchai, S.; Worasuttayangkurn, L.; Mahidol, C.; Ruchirawat, M.; Satayavivad, J. Evaluation of free radical scavenging and antityrosinase activities of standardized longan fruit extract. Food Chem. Toxicol. 2007, 45, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Morigny, P.; Boucher, J.; Arner, P.; Langin, D. Lipid and glucose metabolism in white adipocytes: Pathways, dysfunction and therapeutics. Nat. Rev. Endocrinol. 2021, 17, 276–295. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zheng, Y.; Tang, W.; Yan, W.; Nie, H.; Fang, J.; Liu, G. Dietary polyphenols in lipid metabolism: A role of gut microbiome. Anim. Nutr. 2020, 6, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Ishida, N.; Iizuka, M.; Kataoka, K.; Okazaki, M.; Shiraishi, K.; Yagi, Y.; Jobu, K.; Yokota, J.; Oishi, M.; Moriyama, H.; et al. Improvement of blood lipid profiles by Goishi tea polyphenols in a randomised, double-blind, placebo-controlled clinical study. Int. J. Food Sci. Nutr. 2018, 69, 598–607. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Burke, A.C.; Huff, M.W. Citrus Flavonoids as Regulators of Lipoprotein Metabolism and Atherosclerosis. Annu. Rev. Nutr. 2016, 36, 275–299. [Google Scholar] [CrossRef]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef]
- Ducheix, S.; Montagner, A.; Theodorou, V.; Ferrier, L.; Guillou, H. The liver X receptor: A master regulator of the gut-liver axis and a target for non alcoholic fatty liver disease. Biochem. Pharmacol. 2013, 86, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.J.; Yu, Y.; Hu, S.K.; Zhang, J.H.; Yang, H.X.; Han, B.; Cheng, Y.; Luo, X.Q. Sesamin ameliorates hepatic steatosis and inflammation in rats on a high-fat diet via LXR alpha and PPAR alpha. Nutr. Res. 2016, 36, 1022–1030. [Google Scholar] [CrossRef]
- Ge, C.X.; Yu, R.; Xu, M.X.; Li, P.Q.; Fan, C.Y.; Li, J.M.; Kong, L.D. Betaine prevented fructose-induced NAFLD by regulating LXRa/PPARcic pathway and alleviating ER stress in rats. Eur. J. Pharmacol. 2016, 770, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Ebbert, J.O.; Jensen, M.D. Fat Depots, Free Fatty Acids, and Dyslipidemia. Nutrients 2013, 5, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Castillo, M.; Ortega, A.; Durant, P.; Pirela, D.; Marquina, M.; Cano, C.; Salazar, J.; Gonzalez, M.C.; Bermudez, V.; Rojas-Quintero, J.; et al. Phytotherapy for Cardiovascular Disease: A Bench-to-Bedside Approach. Curr. Pharm. Des. 2020, 26, 4410–4429. [Google Scholar] [CrossRef]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Rahmani, S.; Asgary, S.; Askari, G.; Keshvari, M.; Hatamipour, M.; Feizi, A.; Sahebkar, A. Treatment of Non-alcoholic Fatty Liver Disease with Curcumin: A Randomized Placebo-controlled Trial. Phytother. Res. 2016, 30, 1540–1548. [Google Scholar] [CrossRef]
- Jiao, X.Y.; Wang, Y.H.; Lin, Y.; Lang, Y.X.; Li, E.H.; Zhang, X.Y.; Zhang, Q.; Feng, Y.; Meng, X.J.; Li, B. Blueberry polyphenols extract as a potential prebiotic with anti-obesity effects on C57BL/6 J mice by modulating the gut microbiota. J. Nutr. Biochem. 2019, 64, 88–100. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Van Hee, V.F.; Bindels, L.B.; De Backer, F.; Cani, P.D.; Delzenne, N.M. Polyphenol-rich extract of pomegranate peel alleviates tissue inflammation and hypercholesterolaemia in high-fat diet-induced obese mice: Potential implication of the gut microbiota. Br. J. Nutr. 2013, 109, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Pan, R.; Li, M.; Li, X.; Zhang, H. HPLC Profile of Longan (cv. Shixia) Pericarp-Sourced Phenolics and Their Antioxidant and Cytotoxic Effects. Molecules 2019, 24, 619. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cao, J.X.; Zhao, T.R.; Liu, Y.P.; Khan, A.; Cheng, G.G. The Bioavailability, Extraction, Biosynthesis and Distribution of Natural Dihydrochalcone: Phloridzin. Int. J. Mol. Sci. 2021, 22, 962. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Xie, H.H.; Hao, J.; Jiang, Y.M.; Wei, X.Y. Flavonoid Glycosides from the Seeds of Litchi chinensis. J. Agric. Food Chem. 2011, 59, 1205–1209. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.; Shi, J.; Chen, F.; Ashraf, M. Extraction and pharmacological properties of bioactive compounds from longan (Dimocarpus longan Lour.) fruit—A review. Food Res. Int. 2011, 44, 1837–1842. [Google Scholar] [CrossRef]
- Sun, J.; Shi, J.; Jiang, Y.; Xue, S.J.; Wei, X. Identification of two polyphenolic compounds with antioxidant activities in longan pericarp tissues. J. Agric. Food Chem. 2007, 55, 5864–5868. [Google Scholar] [CrossRef]
- He, N.; Wang, Z.; Yang, C.; Lu, Y.; Sun, D.; Wang, Y.; Shao, W.; Li, Q. Isolation and identification of polyphenolic compounds in longan pericarp. Sep. Purif. Technol. 2009, 70, 219–224. [Google Scholar] [CrossRef]
- Prasad, K.N.; Yang, B.; Shi, J.; Yu, C.; Zhao, M.; Xue, S.; Jiang, Y. Enhanced antioxidant and antityrosinase activities of longan fruit pericarp by ultra-high-pressure-assisted extraction. J. Pharm. Biomed. Anal. 2010, 51, 471–477. [Google Scholar] [CrossRef]
- Škrovánková, S.; Mišurcová, L.; Machů, L. Antioxidant activity and protecting health effects of common medicinal plants. Adv. Food Nutr. Res. 2012, 67, 75–139. [Google Scholar]
- Tan, S.; Wang, Z.W.; Xiang, Y.Y.; Deng, T.W.; Zhao, X.; Shi, S.Y.; Zheng, Q.R.; Gao, X.X.; Li, W.F. The effects of drying methods on chemical profiles and antioxidant activities of two cultivars of Psidium guajava fruits. LWT-Food Sci. Technol. 2020, 118, 108723. [Google Scholar] [CrossRef]
- Jang, A.; Srinivasan, P.; Lee, N.Y.; Song, H.P.; Lee, J.W.; Lee, M.; Jo, C. Comparison of hypolipidemic activity of synthetic gallic acid-linoleic acid ester with mixture of gallic acid and linoleic acid, gallic acid, and linoleic acid on high-fat diet induced obesity in C57BL/6 Cr Slc mice. Chem.-Biol. Interact. 2008, 174, 109–117. [Google Scholar] [CrossRef]
- Fanaei, H.; Mard, S.A.; Sarkaki, A.; Goudarzi, G.; Khorsandi, L. Gallic acid protects the liver against NAFLD induced by dust exposure and high-fat diet through inhibiting oxidative stress and repressing the inflammatory signaling pathways NF-kbeta/TNF-alpha/IL-6 in Wistar rats. Avicenna J. Phytomed. 2021, 11, 527–540. [Google Scholar]
- Cheng, H.; Xu, N.; Zhao, W.; Su, J.; Liang, M.; Xie, Z.; Wu, X.; Li, Q. (-)-Epicatechin regulates blood lipids and attenuates hepatic steatosis in rats fed high-fat diet. Mol. Nutr. Food Res. 2017, 61, 1700303. [Google Scholar] [CrossRef]
- Salvadó, M.J.; Casanova, E.; Fernández-Iglesias, A.; Arola, L.; Bladé, C. Roles of proanthocyanidin rich extracts in obesity. Food Funct. 2015, 6, 1053–1071. [Google Scholar] [CrossRef]
- Baselga-Escudero, L.; Pascual-Serrano, A.; Ribas-Latre, A.; Casanova, E.; Salvado, M.J.; Arola, L.; Arola-Arnal, A.; Blade, C. Long-term supplementation with a low dose of proanthocyanidins normalized liver miR-33a and miR-122 levels in high-fat diet-induced obese rats. Nutr. Res. 2015, 35, 337–345. [Google Scholar] [CrossRef]
- Najafian, M.; Jahromi, M.Z.; Nowroznejhad, M.J.; Khajeaian, P.; Kargar, M.M.; Sadeghi, M.; Arasteh, A. Phloridzin reduces blood glucose levels and improves lipids metabolism in streptozotocin-induced diabetic rats. Mol. Biol. Rep. 2012, 39, 5299–5306. [Google Scholar] [CrossRef]
- Liu, H.Z.; Chen, Y.E.; Wen, Y.F.; Zhu, S.M.; Huang, S.; He, L.; Hou, S.Z.; Lai, X.P.; Chen, S.X.; Dai, Z.H.; et al. Phloridzin Ameliorates Lipid Deposition in High-Fat-Diet-Fed Mice with Nonalcoholic Fatty Liver Disease via Inhibiting the mTORC1/SREBP-1c Pathway. J. Agric. Food Chem. 2021, 69, 8671–8683. [Google Scholar] [CrossRef]
- Tang, Y.-Y.; He, X.-M.; Sun, J.; Li, C.-B.; Li, L.; Sheng, J.-F.; Xin, M.; Li, Z.-C.; Zheng, F.-J.; Liu, G.-M.; et al. Polyphenols and Alkaloids in Byproducts of Longan Fruits (Dimocarpus longan Lour.) and Their Bioactivities. Molecules 2019, 24, 1186. [Google Scholar] [CrossRef]
- Sinha, R.A.; Rajak, S.; Singh, B.K.; Yen, P.M. Hepatic Lipid Catabolism via PPARα-Lysosomal Crosstalk. Int. J. Mol. Sci. 2020, 21, 2391. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Kimmel, R.; Stroup, D. Regulation of cholesterol 7 alpha-hydroxylase gene (CYP7A1) transcription by the liver orphan receptor (LXR alpha). Gene 2001, 262, 257–265. [Google Scholar] [CrossRef]
- Zhao, L.; Lei, W.; Deng, C.; Wu, Z.; Sun, M.; Jin, Z.; Song, Y.; Yang, Z.; Jiang, S.; Shen, M.; et al. The roles of liver X receptor α in inflammation and inflammation-associated diseases. J. Cell. Physiol. 2021, 236, 4807–4828. [Google Scholar] [CrossRef]
- Abenavoli, L.; Larussa, T.; Corea, A.; Procopio, A.C.; Boccuto, L.; Dallio, M.; Federico, A.; Luzza, F. Dietary Polyphenols and Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 494. [Google Scholar] [CrossRef]
- Yang, D.-J.; Chang, Y.-Y.; Hsu, C.-L.; Liu, C.-W.; Lin, Y.-L.; Lin, Y.-H.; Liu, K.-C.; Chen, Y.-C. Antiobesity and Hypolipidemic Effects of Polyphenol-Rich Longan (Dimocarpus longans Lour.) Flower Water Extract in Hypercaloric-Dietary Rats. J. Agric. Food Chem. 2010, 58, 2020–2027. [Google Scholar] [CrossRef]
- Liu, C.-W.; Yang, D.-J.; Chang, Y.-Y.; Hsu, C.-L.; Tseng, J.-K.; Chang, M.-H.; Wang, M.; Chen, Y.-C. Polyphenol-rich longan (Dimocarpus longan Lour.)-flower-water-extract attenuates nonalcoholic fatty liver via decreasing lipid peroxidation and downregulating matrix metalloproteinases-2 and-9. Food Res. Int. 2012, 45, 444–449. [Google Scholar] [CrossRef]
- Ke, Z.L.; Tan, S.; Li, H.Y.; Jiang, S.S.; Li, Y.P.; Chen, R.; Li, M.X. Tangeretin improves hepatic steatosis and oxidative stress through the Nrf2 pathway in high fat diet-induced nonalcoholic fatty liver disease mice. Food Funct. 2022, 13, 2782–2790. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | Retention Time (min) | Fragmentor (V) | MS [M-H]- | MS/MS (m/z) | Contents (mg/g) |
|---|---|---|---|---|---|---|
| 1 | Gallic acid | 2.966 | 100 | 168.9 | 125, 106.8 | 24.080 ± 2.791 |
| 2 | Proanthocyanidin B2 | 4.838 | 100 | 577 | 289, 407 | 2.106 ± 0.245 |
| 3 | Epicatechin | 5.588 | 130 | 289 | 109, 203 | 7.592 ± 0.231 |
| 4 | Proanthocyanidin A2 | 6.131 | 150 | 575 | 285, 423 | 24.382 ± 2.859 |
| 5 | Syringic acid | 6.426 | 100 | 197 | 122.8, 182 | 2.361 ± 0.106 |
| 6 | p-hydroxybenzoic acid | 6.584 | 130 | 137 | 137 | 0.279 ± 0.061 |
| 7 | Poncirin | 6.755 | 150 | 593 | 284.9 | 0.607 ± 0.044 |
| 8 | Ferulic acid | 6.984 | 90 | 192.9 | 134, 149 | 1.402 ± 0.077 |
| 9 | Rutin | 7.253 | 160 | 609 | 299.9 | 0.050 ± 0.004 |
| 10 | Phlorizin | 7.521 | 150 | 434.9 | 272.9, 167 | 38.894 ± 3.765 |
| 11 | Methyl hesperidin | 7.849 | 135 | 623 | 315, 338.7 | 0.488 ± 0.004 |
| Total polyphenols | 285.350 ± 36.430 (mg GAE/g) | |||||
| DPPH | 231.350 ± 21.640 (mg Vc/g) | |||||
| ABTS | 252.380 ± 31.150 (mg Vc /g) | |||||
| FRAP | 558.220 ± 59.810 (mg Vc/g) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, S.; Ke, Z.; Zhou, C.; Luo, Y.; Ding, X.; Luo, G.; Li, W.; Shi, S. Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts. Molecules 2023, 28, 2083. https://doi.org/10.3390/molecules28052083
Tan S, Ke Z, Zhou C, Luo Y, Ding X, Luo G, Li W, Shi S. Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts. Molecules. 2023; 28(5):2083. https://doi.org/10.3390/molecules28052083
Chicago/Turabian StyleTan, Si, Zunli Ke, Chongbing Zhou, Yuping Luo, Xiaobo Ding, Gangjun Luo, Wenfeng Li, and Shengyou Shi. 2023. "Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts" Molecules 28, no. 5: 2083. https://doi.org/10.3390/molecules28052083
APA StyleTan, S., Ke, Z., Zhou, C., Luo, Y., Ding, X., Luo, G., Li, W., & Shi, S. (2023). Polyphenol Profile, Antioxidant Activity, and Hypolipidemic Effect of Longan Byproducts. Molecules, 28(5), 2083. https://doi.org/10.3390/molecules28052083