


Full-Chain FeCl3 Catalyzation Is Sufficient to Boost Cellulase Secretion and Cellulosic Ethanol along with Valorized Supercapacitor and Biosorbent Using Desirable Corn Stalk


 ,
,  ,
, 
Abstract
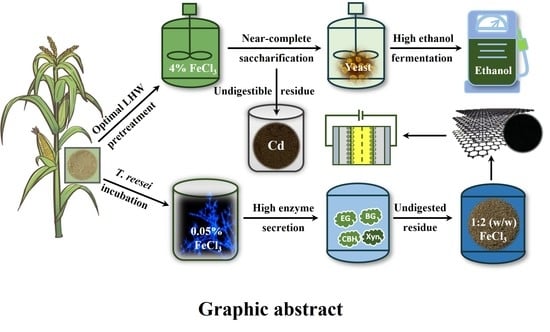
1. Introduction
2. Results and Discussion
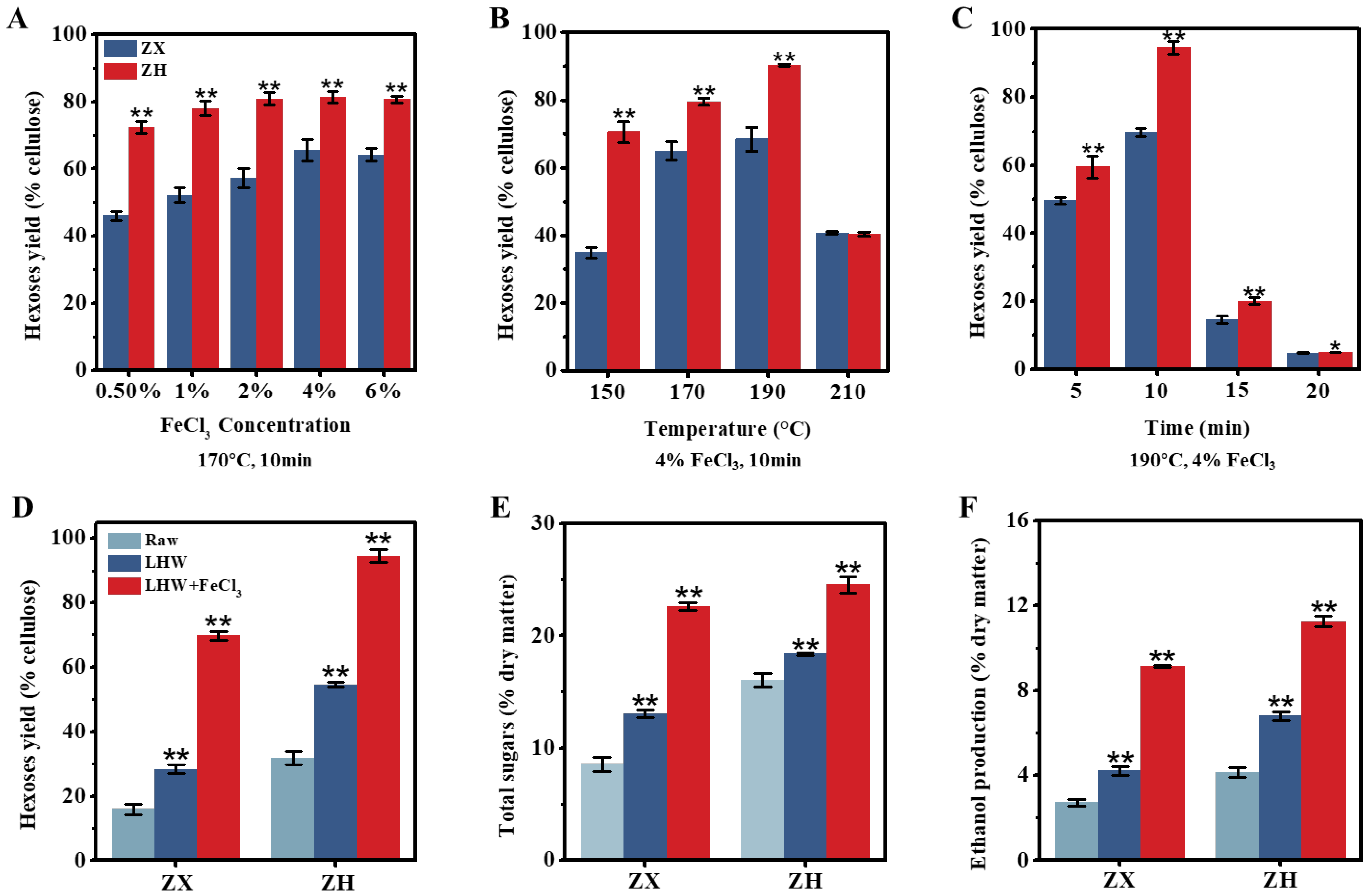
2.1. Enhanced Biomass Saccharification of Corn Stalks for Bioethanol Production under Optimal LHW Pretreatment with FeCl3 Co-Supplement
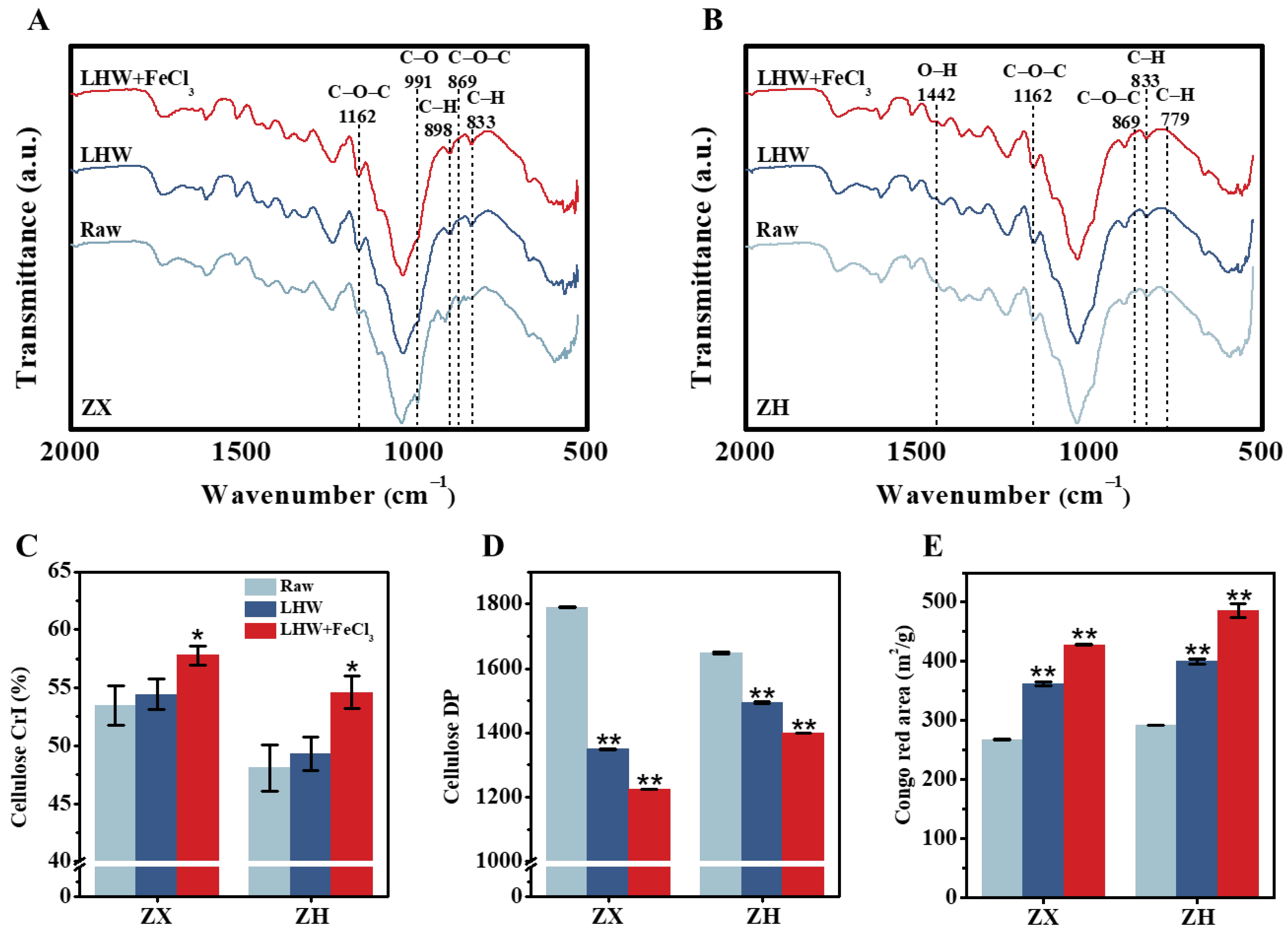
2.2. Improved Lignocellulose Recalcitrance from FeCl3 Catalysis in Corn Stalks
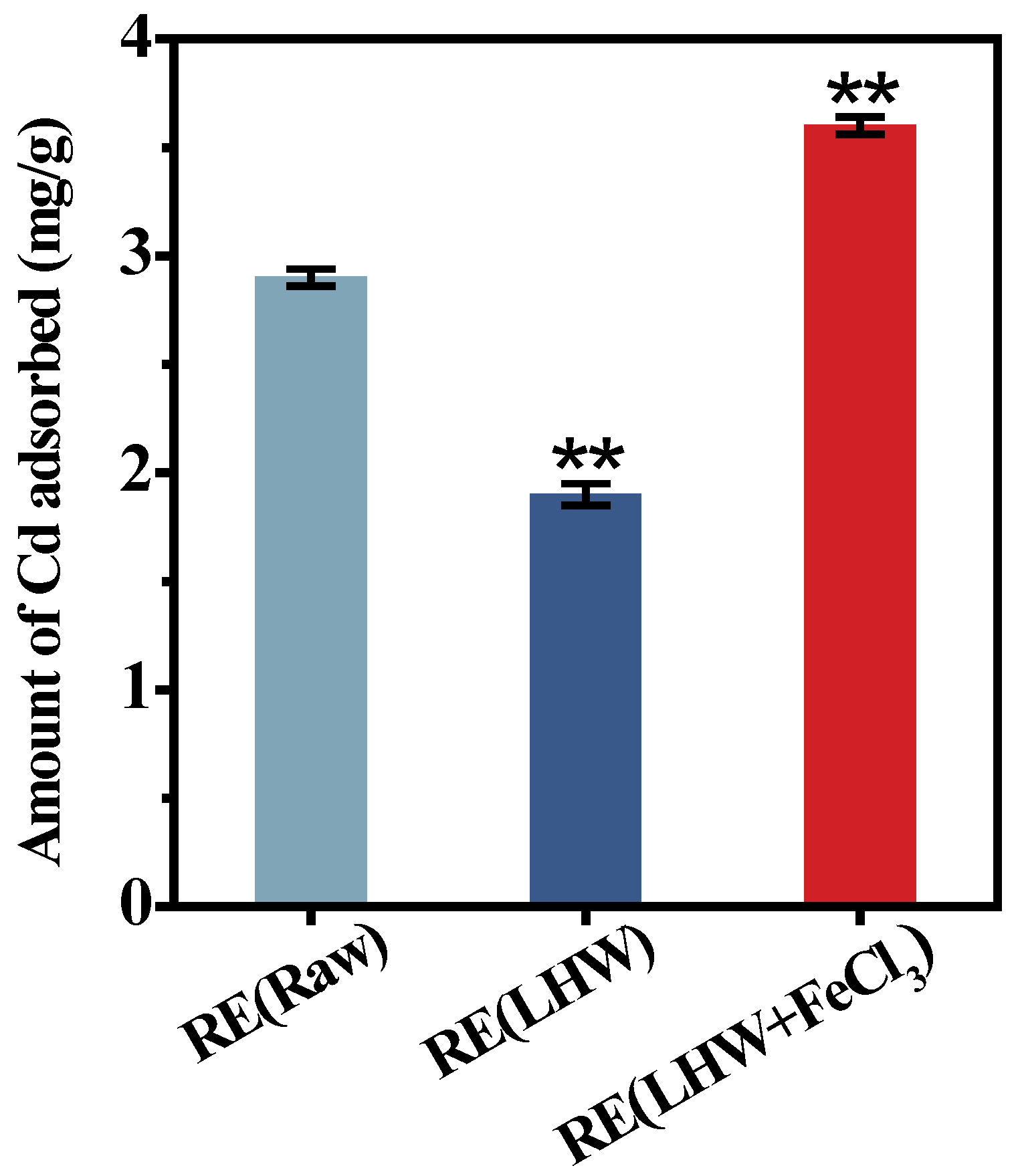
2.3. Enzyme-Undigestible Lignocellulose as Active Biosorbent for Cd Adsorption
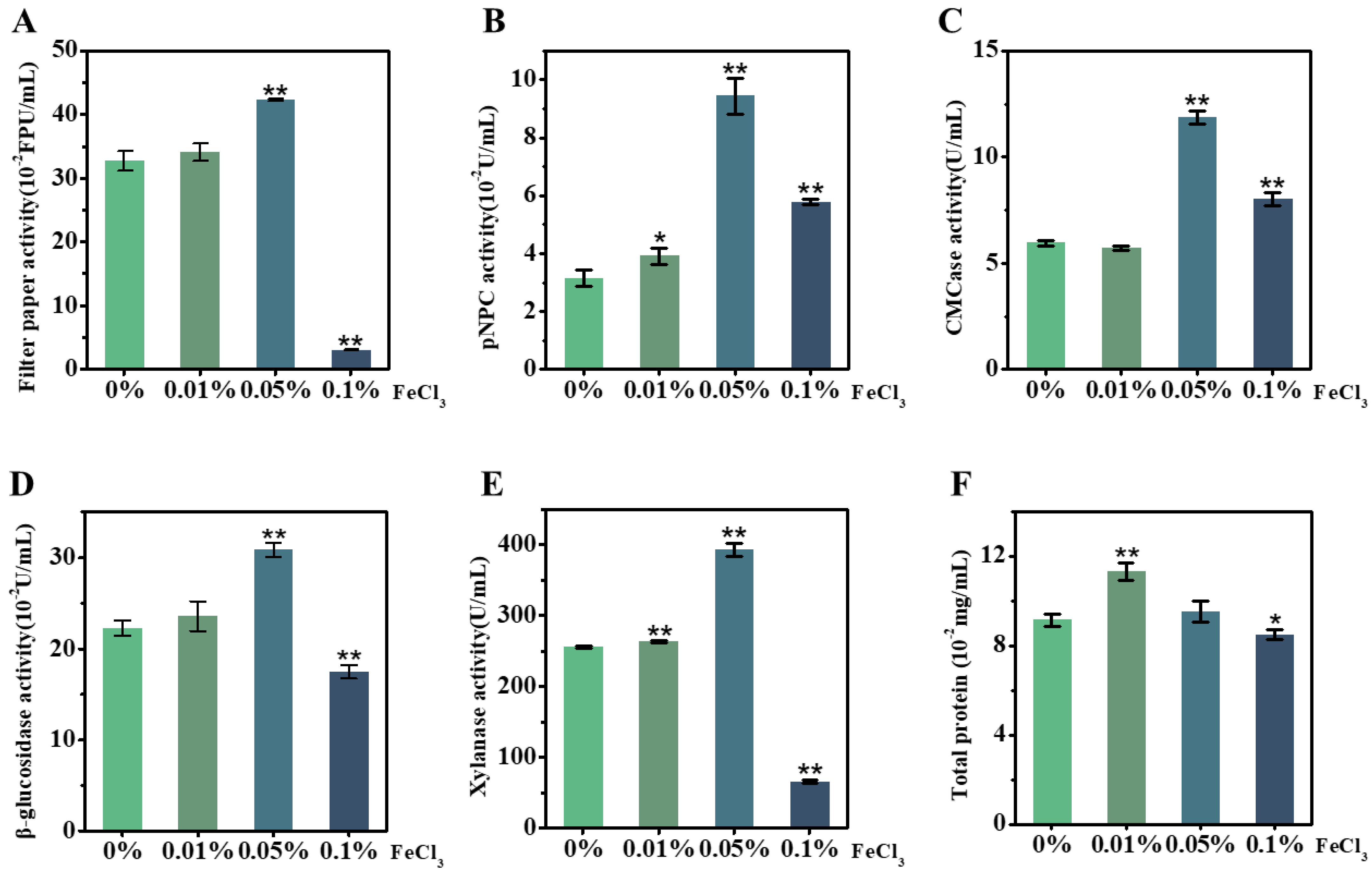
2.4. FeCl3 Supplement for Upgraded Lignocellulose-Degradation-Enzyme Activity Secreted by T. reesei Incubation with Corn Stalk
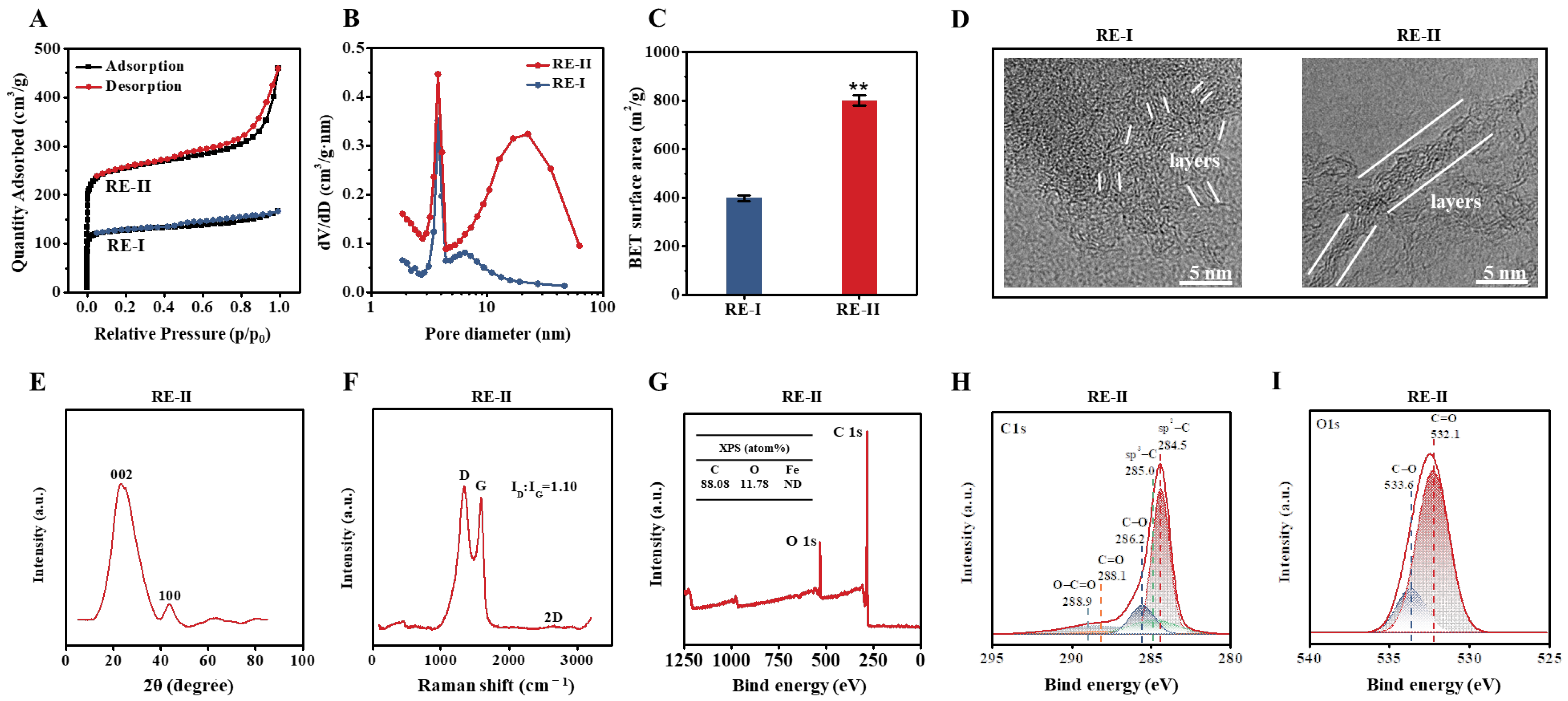
2.5. Porosity-Raised Biocarbon Generated by FeCl3 Activation with T. reesei-Undigested Lignocellulose Residues
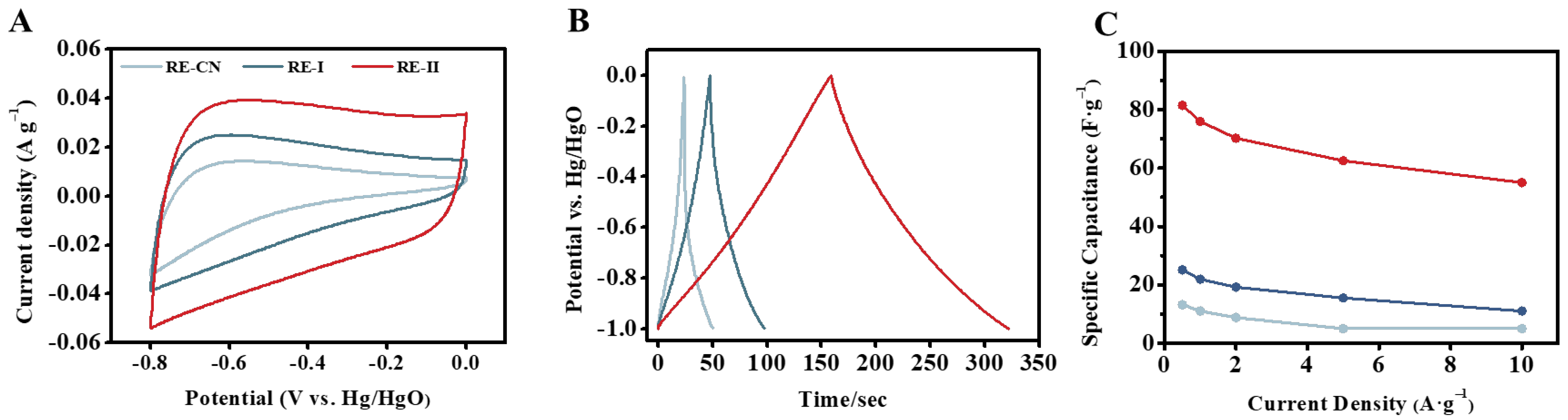
2.6. Improved Supercapacitor Performance of the Porous Carbon Generated by FeCl3 Activation
3. Materials and Methods
3.1. Collection of Mature Stalks in Two Corn Cultivars
3.2. Wall-Polymer Extraction and Determination
3.3. Detection of Wall-Polymer Features
3.4. Lignocellulose Observation and Characterization
3.5. Liquid Hot Water (LHW) Pretreatment Co-Supplied with FeCl3
3.6. Enzymatic Hydrolysis and Yeast Fermentation for Bioethanol Production
3.7. Cd-Adsorption Analysis
3.8. T. reesei Strain Cultivation Induced by FeCl3 and Enzyme-Activity Determination
3.9. Preparation of Biocarbon Materials
3.10. Characterization of Porous Carbons
3.11. Measurement of Electrochemical Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ragauskas, A.J.; Williams, C.K.; Davison, B.H.; Britovsek, G.; Cairney, J.; Eckert, C.A.; Frederick, W.J.; Hallett, J.P.; Leak, D.J.; Liotta, C.L.; et al. The path forward for biofuels and biomaterials. Science 2006, 311, 484–489. [Google Scholar] [CrossRef]
- Rubin, E.M. Genomics of cellulosic biofuels. Nature 2008, 454, 841–845. [Google Scholar] [CrossRef]
- Carroll, A.; Somerville, C. Cellulosic Biofuels. Annu. Rev. Plant Biol. 2009, 60, 165–182. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, C.; Hu, H.; Li, Y.; Sun, D.; Wang, Y.; Peng, L. Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnol. Adv. 2016, 34, 997–1017. [Google Scholar] [CrossRef]
- Lynd, L.R.; Beckham, G.T.; Guss, A.M.; Jayakody, L.N.; Karp, E.M.; Maranas, C.; McCormick, R.L.; Amador-Noguez, D.; Bomble, Y.J.; Davison, B.H.; et al. Toward low-cost biological and hybrid biological/catalytic conversion of cellulosic biomass to fuels. Energy Environ. Sci. 2022, 15, 938–990. [Google Scholar] [CrossRef]
- Yoo, C.G.; Meng, X.; Pu, Y.; Ragauskas, A.J. The critical role of lignin in lignocellulosic biomass conversion and recent pretreatment strategies: A comprehensive review. Bioresour. Technol. 2020, 301, 122784. [Google Scholar] [CrossRef]
- Li, Y.; Liu, P.; Huang, J.; Zhang, R.; Hu, Z.; Feng, S.; Wang, Y.; Wang, L.; Xia, T.; Peng, L. Mild chemical pretreatments are sufficient for bioethanol production in transgenic rice straws overproducing glucosidase. Green Chem. 2018, 20, 2047–2056. [Google Scholar] [CrossRef]
- Sun, D.; Yang, Q.; Wang, Y.; Gao, H.; He, M.; Lin, X.; Lu, J.; Wang, Y.; Kang, H.; Alam, A.; et al. Distinct mechanisms of enzymatic saccharification and bioethanol conversion enhancement by three surfactants under steam explosion and mild chemical pretreatments in bioenergy Miscanthus. Ind. Crops Prod. 2020, 153, 112559. [Google Scholar] [CrossRef]
- Chen, W.-H.; Nižetić, S.; Sirohi, R.; Huang, Z.; Luque, R.M.; Papadopoulos, A.; Sakthivel, R.; Phuong Nguyen, X.; Tuan Hoang, A. Liquid hot water as sustainable biomass pretreatment technique for bioenergy production: A review. Bioresour. Technol. 2022, 344, 126207. [Google Scholar] [CrossRef]
- Wells, J.M.; Drielak, E.; Surendra, K.C.; Kumar Khanal, S. Hot water pretreatment of lignocellulosic biomass: Modeling the effects of temperature, enzyme and biomass loadings on sugar yield. Bioresour. Technol. 2020, 300, 122593. [Google Scholar] [CrossRef]
- Novy, V.; Nielsen, F.; Seiboth, B.; Nidetzky, B. The influence of feedstock characteristics on enzyme production in Trichoderma reesei: A review on productivity, gene regulation and secretion profiles. Biotechnol. Biofuels 2019, 12, 238. [Google Scholar] [CrossRef]
- Kang, K.; Wang, S.; Lai, G.; Liu, G.; Xing, M. Characterization of a novel swollenin from Penicillium oxalicum in facilitating enzymatic saccharification of cellulose. BMC Biotechnol. 2013, 13, 42. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, G.; Wei, X.-Y.; Liu, R.; Gu, J.-J.; Cao, F.-F. Bioselective Synthesis of a Porous Carbon Collector for High-Performance Sodium-Metal Anodes. J. Am. Chem. Soc. 2021, 143, 3280–3283. [Google Scholar] [CrossRef]
- Lizundia, E.; Luzi, F.; Puglia, D. Organic waste valorisation towards circular and sustainable biocomposites. Green Chem. 2022, 24, 5429–5459. [Google Scholar] [CrossRef]
- Chakraborty, R.; Vilya, K.; Pradhan, M.; Nayak, A.K. Recent advancement of biomass-derived porous carbon based materials for energy and environmental remediation applications. J. Mater. Chem. A 2022, 10, 6965–7005. [Google Scholar] [CrossRef]
- Deng, J.; Li, M.; Wang, Y. Biomass-derived carbon: Synthesis and applications in energy storage and conversion. Green Chem. 2016, 18, 4824–4854. [Google Scholar] [CrossRef]
- Dong, J.; Li, S.; Ding, Y. Anchoring nickel-cobalt sulfide nanoparticles on carbon aerogel derived from waste watermelon rind for high-performance asymmetric supercapacitors. J. Alloys Compd. 2020, 845, 155701. [Google Scholar] [CrossRef]
- Zou, K.; Deng, Y.; Wu, W.; Zhang, S.; Chen, G. A novel eutectic solvent precursor for efficiently preparing N-doped hierarchically porous carbon nanosheets with unique surface functional groups and micropores towards dual-carbon lithium-ion capacitors. J. Mater. Chem. A 2021, 9, 13631–13641. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, X.; Wang, L.; Fu, H. Recent advances of biomass derived carbon-based materials for efficient electrochemical energy devices. J. Mater. Chem. A 2022, 10, 9277–9307. [Google Scholar] [CrossRef]
- Tejirian, A.; Xu, F. Inhibition of Cellulase-Catalyzed Lignocellulosic Hydrolysis by Iron and Oxidative Metal Ions and Complexes. Appl. Environ. Microbiol. 2010, 76, 7673–7682. [Google Scholar] [CrossRef]
- Rodriguez, J.; Ferraz, A.; de Mello, M.P. Role of Metals in Wood Biodegradation. In Wood Deterioration and Preservation; American Chemical Society: Washington, DC, USA, 2003; Volume 845, pp. 154–174. [Google Scholar]
- Huang, L.; Forsberg, C.W.; Thomas, D.Y. Purification and characterization of a chloride-stimulated cellobiosidase from Bacteroides succinogenes S85. J. Bacteriol. 1988, 170, 2923–2932. [Google Scholar] [CrossRef]
- Zhao, Y.; Damgaard, A.; Christensen, T.H. Bioethanol from corn stover—A review and technical assessment of alternative biotechnologies. Prog. Energy Combust. Sci. 2018, 67, 275–291. [Google Scholar] [CrossRef]
- Kamireddy, S.R.; Li, J.B.; Tucker, M.; Degenstein, J.; Ji, Y. Effects and Mechanism of Metal Chloride Salts on Pretreatment and Enzymatic Digestibility of Corn Stover. Ind. Eng. Chem. Res. 2013, 52, 1775–1782. [Google Scholar] [CrossRef]
- Yu, H.; Hu, M.; Hu, Z.; Liu, F.; Yu, H.; Yang, Q.; Gao, H.; Xu, C.; Wang, M.; Zhang, G.; et al. Insights into pectin dominated enhancements for elimination of toxic Cd and dye coupled with ethanol production in desirable lignocelluloses. Carbohydr. Polym. 2022, 286, 119298. [Google Scholar] [CrossRef]
- Guo, H.P.; Hong, C.T.; Chen, X.M.; Xu, Y.X.; Liu, Y.; Jiang, D.A.; Zheng, B.S. Different Growth and Physiological Responses to Cadmium of the Three Miscanthus Species. PLoS ONE 2016, 11, e0153475. [Google Scholar] [CrossRef]
- Xu, F.; Yu, J.M.; Tesso, T.; Dowell, F.; Wang, D.H. Qualitative and quantitative analysis of lignocellulosic biomass using infrared techniques: A mini-review. Appl. Energy 2013, 104, 801–809. [Google Scholar] [CrossRef]
- Ong, H.C.; Chen, W.H.; Singh, Y.; Gan, Y.Y.; Chen, C.Y.; Show, P.L. A state-of-the-art review on thermochemical conversion of biomass for biofuel production: A TG-FTIR approach. Energy Convers. Manag. 2020, 209, 11264. [Google Scholar] [CrossRef]
- Pancholi, M.J.; Khristi, A.; Athira, K.M.; Bagchi, D. Comparative Analysis of Lignocellulose Agricultural Waste and Pre-treatment Conditions with FTIR and Machine Learning Modeling. BioEnergy Res. 2022. [Google Scholar] [CrossRef]
- Sammons, R.J.; Harper, D.P.; Labbe, N.; Bozell, J.J.; Elder, T.; Rials, T.G. Characterization of Organosolv Lignins using Thermal and FT-IR Spectroscopic Analysis. BioResources 2013, 8, 2752–2767. [Google Scholar] [CrossRef]
- Zhuang, J.; Li, M.; Pu, Y.; Ragauskas, A.J.; Yoo, C.G. Observation of Potential Contaminants in Processed Biomass Using Fourier Transform Infrared Spectroscopy. Appl. Sci. 2020, 10, 4345. [Google Scholar] [CrossRef]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M.J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef]
- Funahashi, R.; Ono, Y.; Tanaka, R.; Yokoi, M.; Daido, K.; Inamochi, T.; Saito, T.; Horikawa, Y.; Isogai, A. Changes in the degree of polymerization of wood celluloses during dilute acid hydrolysis and TEMPO-mediated oxidation: Formation mechanism of disordered regions along each cellulose microfibril. Int. J. Biol. Macromol. 2018, 109, 914–920. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, Z.; He, J.; Li, Y.; Pan, C.; Wang, C.; Deng, M.-R.; Zhu, H. A myxobacterial LPMO10 has oxidizing cellulose activity for promoting biomass enzymatic saccharification of agricultural crop straws. Bioresour. Technol. 2020, 318, 124217. [Google Scholar] [CrossRef]
- Pihlajaniemi, V.; Sipponen, M.H.; Liimatainen, H.; Sirviö, J.A.; Nyyssölä, A.; Laakso, S. Weighing the factors behind enzymatic hydrolyzability of pretreated lignocellulose. Green Chem. 2016, 18, 1295–1305. [Google Scholar] [CrossRef]
- Xu, C.; Zhu, J.; Yu, H.; Yu, H.; Yang, Y.; Fu, Q.; Zhan, D.; Wang, Y.; Wang, H.; Zhang, Y.; et al. Recyclable cascading of arsenic phytoremediation and lead removal coupled with high bioethanol production using desirable rice straws. Biochem. Eng. J. 2021, 168, 107950. [Google Scholar] [CrossRef]
- Liu, P.; Li, A.; Wang, Y.; Cai, Q.; Yu, H.; Li, Y.; Peng, H.; Li, Q.; Wang, Y.; Wei, X.; et al. Distinct Miscanthus lignocellulose improves fungus secreting cellulases and xylanases for consistently enhanced biomass saccharification of diverse bioenergy crops. Renew. Energy 2021, 174, 799–809. [Google Scholar] [CrossRef]
- Peng, H.; Zhao, W.; Liu, J.; Liu, P.; Yu, H.; Deng, J.; Yang, Q.; Zhang, R.; Hu, Z.; Liu, S.; et al. Distinct cellulose nanofibrils generated for improved Pickering emulsions and lignocellulose-degradation enzyme secretion coupled with high bioethanol production in natural rice mutants. Green Chem. 2022, 24, 2975–2987. [Google Scholar] [CrossRef]
- Chen, L.; Zou, G.; Wang, J.; Wang, J.; Liu, R.; Jiang, Y.; Zhao, G.; Zhou, Z. Characterization of the Ca2+-responsive signaling pathway in regulating the expression and secretion of cellulases in Trichoderma reesei Rut-C30. Mol. Microbiol. 2016, 100, 560–575. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, Y.; Wang, W.; Wei, D. Mn2+ modulates the expression of cellulase genes in Trichoderma reesei Rut-C30 via calcium signaling. Biotechnol. Biofuels 2018, 11, 54. [Google Scholar] [CrossRef]
- Yenenler, A.; Kurt, H.; Sezerman, O.U. Enhancing Enzymatic Properties of Endoglucanase I Enzyme from Trichoderma reesei via Swapping from Cellobiohydrolase I Enzyme. Catalysts 2019, 9, 130. [Google Scholar] [CrossRef]
- Arampatzidou, A.C.; Deliyanni, E.A. Comparison of activation media and pyrolysis temperature for activated carbons development by pyrolysis of potato peels for effective adsorption of endocrine disruptor bisphenol-A. J. Colloid Interface Sci. 2016, 466, 101–112. [Google Scholar] [CrossRef]
- Ferrari, A.C.; Robertson, J. Interpretation of Raman spectra of disordered and amorphous carbon. Phys. Rev. B 2000, 61, 14095–14107. [Google Scholar] [CrossRef]
- Kim, J.B.; Koo, S.H.; Kim, I.H.; Kim, J.T.; Kim, J.G.; Jayaraman, B.; Lim, J.; Kim, S.O. Characteristic dual-domain composite structure of reduced graphene oxide and its application to higher specific capacitance. Chem. Eng. J. 2022, 446, 137390. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, G.; Li, M.-Y.; Yin, Y.-X.; Li, J.-Y.; Li, G.; Wang, W.-P.; Peng, W.; Cao, F.-F.; Guo, Y.-G. Porous carbon for high-energy density symmetrical supercapacitor and lithium-ion hybrid electrochemical capacitors. Chem. Eng. J. 2019, 375, 122020. [Google Scholar] [CrossRef]
- Kong, X.; Zhu, Y.; Lei, H.; Wang, C.; Zhao, Y.; Huo, E.; Lin, X.; Zhang, Q.; Qian, M.; Mateo, W.; et al. Synthesis of graphene-like carbon from biomass pyrolysis and its applications. Chem. Eng. J. 2020, 399, 125808. [Google Scholar] [CrossRef]
- Wang, Y.; Rong, Z.; Wang, Y.; Wang, T.; Du, Q.; Wang, Y.; Qu, J. Graphene-Based Metal/Acid Bifunctional Catalyst for the Conversion of Levulinic Acid to γ-Valerolactone. ACS Sustain. Chem. Eng. 2017, 5, 1538–1548. [Google Scholar] [CrossRef]
- Chen, Z.; Liao, S.; Ge, L.; Amaniampong, P.N.; Min, Y.; Wang, C.; Li, K.; Lee, J.-M. Reduced graphene oxide with controllably intimate bifunctionality for the catalytic transformation of fructose into 2,5-diformylfuran in biphasic solvent systems. Chem. Eng. J. 2020, 379, 122284. [Google Scholar] [CrossRef]
- Lakmini, L.M.N.; Deshan, A.D.K.; Pham, H.D.; Doherty, W.; Rackemann, D.; Dubal, D.P.; Moghaddam, L. High carbon utilization: 5-(Chloromethyl)furfural (CMF) production from rice by-products and transformation of CMF residues into Li-ion energy storage systems. J. Clean. Prod. 2022, 375, 134082. [Google Scholar] [CrossRef]
- Xie, Z.; Shang, X.; Yan, J.; Hussain, T.; Nie, P.; Liu, J. Biomass-derived porous carbon anode for high-performance capacitive deionization. Electrochim. Acta 2018, 290, 666–675. [Google Scholar] [CrossRef]
- Yumak, T.; Yakaboylu, G.A.; Oginni, O.; Singh, K.; Ciftyurek, E.; Sabolsky, E.M. Comparison of the electrochemical properties of engineered switchgrass biomass-derived activated carbon-based EDLCs. Colloids Surf. A Physicochem. Eng. Asp. 2020, 586, 124150. [Google Scholar] [CrossRef]
- Peng, L.; Hocart, C.H.; Redmond, J.W.; Williamson, R.E. Fractionation of carbohydrates in Arabidopsis root cell walls shows that three radial swelling loci are specifically involved in cellulose production. Planta 2000, 211, 406–414. [Google Scholar] [CrossRef]
- Li, F.; Xie, G.; Huang, J.; Zhang, R.; Li, Y.; Zhang, M.; Wang, Y.; Li, A.; Li, X.; Xia, T.; et al. OsCESA9 conserved-site mutation leads to largely enhanced plant lodging resistance and biomass enzymatic saccharification by reducing cellulose DP and crystallinity in rice. Plant Biotechnol. J. 2017, 15, 1093–1104. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, M.; Wang, L.; Tu, Y.; Zhang, J.; Xie, G.; Zou, W.; Li, F.; Guo, K.; Li, Q.; et al. Biomass digestibility is predominantly affected by three factors of wall polymer features distinctive in wheat accessions and rice mutants. Biotechnol. Biofuels 2013, 6, 183. [Google Scholar] [CrossRef]
- Alam, A.; Zhang, R.; Liu, P.; Huang, J.; Wang, Y.; Hu, Z.; Madadi, M.; Sun, D.; Hu, R.; Ragauskas, A.J.; et al. A finalized determinant for complete lignocellulose enzymatic saccharification potential to maximize bioethanol production in bioenergy Miscanthus. Biotechnol. Biofuels 2019, 12, 99. [Google Scholar] [CrossRef]
- Wiman, M.; Dienes, D.; Hansen, M.A.T.; van der Meulen, T.; Zacchi, G.; Lidén, G. Cellulose accessibility determines the rate of enzymatic hydrolysis of steam-pretreated spruce. Bioresour. Technol. 2012, 126, 208–215. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.; Li, Y.; Xiong, K.; Wang, Y.; Li, F.; Liu, M.; Wu, Z.; Tu, Y.; Peng, L. Ammonium oxalate-extractable uronic acids positively affect biomass enzymatic digestibility by reducing lignocellulose crystallinity in Miscanthus. Bioresour. Technol. 2015, 196, 391–398. [Google Scholar] [CrossRef]
- Wu, L.; Feng, S.; Deng, J.; Yu, B.; Wang, Y.; He, B.; Peng, H.; Li, Q.; Hu, R.; Peng, L. Altered carbon assimilation and cellulose accessibility to maximize bioethanol yield under low-cost biomass processing in corn brittle stalk. Green Chem. 2019, 21, 4388–4399. [Google Scholar] [CrossRef]
- Jin, W.; Chen, L.; Hu, M.; Sun, D.; Li, A.; Li, Y.; Hu, Z.; Zhou, S.; Tu, Y.; Xia, T.; et al. Tween-80 is effective for enhancing steam-exploded biomass enzymatic saccharification and ethanol production by specifically lessening cellulase absorption with lignin in common reed. Appl. Energy 2016, 175, 82–90. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Yu, H.; Li, Y.; Wang, Y.; Gao, H.; Peng, H.; Hu, Z.; Wang, H.; Zhang, G.; et al. Optimizing two green-like biomass pretreatments for maximum bioethanol production using banana pseudostem by effectively enhancing cellulose depolymerization and accessibility. Sustain. Energy Fuels 2021, 5, 3467–3478. [Google Scholar] [CrossRef]
- Yu, J.-x.; Wang, L.-y.; Chi, R.-a.; Zhang, Y.-f.; Xu, Z.-g.; Guo, J. Adsorption of Pb2+, Cd2+, Cu2+, and Zn2+ from aqueous solution by modified sugarcane bagasse. Res. Chem. Intermed. 2015, 41, 1525–1541. [Google Scholar] [CrossRef]
- Song, W.; Han, X.; Qian, Y.; Liu, G.; Yao, G.; Zhong, Y.; Qu, Y. Proteomic analysis of the biomass hydrolytic potentials of Penicillium oxalicum lignocellulolytic enzyme system. Biotechnol. Biofuels 2016, 9, 68. [Google Scholar] [CrossRef]
- Oyedotun, K.O.; Barzegar, F.; Mirghni, A.A.; Khaleed, A.A.; Masikhwa, T.M.; Manyala, N. Examination of High-Porosity Activated Carbon Obtained from Dehydration of White Sugar for Electrochemical Capacitor Applications. ACS Sustain. Chem. Eng. 2019, 7, 537–546. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Wall Polymer Level (% Dry Matter) | |||
|---|---|---|---|---|
| Cellulose | Hemicellulose | Lignin | Soluble Sugar | |
| ZX | 32.63 ± 1.49 | 17.82 ± 0.06 | 16.7 ± 0.38 | 17.77 ± 0.38 |
| ZH | 26.37 ± 0.60 ** | 19.30 ± 0.26 ** | 13.05 ± 0.63 ** | 22.87 ± 0.94 ** |
| Inducing Substrate | Increased Enzymatic Activities (%) | Reference | ||||
|---|---|---|---|---|---|---|
| FPA | CBH | EG | BG | Xylanase | ||
| Fe3+ | 29.38 | 199.37 | 99.16 | 38.38 | 53.72 | This study |
| Mn2+ | - * | 94 | 62 | - | - | [40] |
| Co2+ | - | - | 58.62 | - | - | [41] |
| Ca2+ | - | 40 | 80 | - | 40 | [39] |
| Protein Name | Accession No. | iBAQ (×106) | MW [kDa] |
|---|---|---|---|
| Exoglucanase I | P62694 | 3009 | 54.07 |
| Endo-β-1,4-glucanase I | G0RKH9 | 656.77 | 48.21 |
| Endo-β-1,4-glucanase II | G0RB67 | 44.16 | 254.04 |
| Endo-β-1,4-glucanase VII | A0A024SFJ2 | 53.24 | 26.8 |
| β-D-glucosidase | Q12715 | 50.95 | 78.43 |
| β-glucosidase | G0RDY1 | 3.42 | 84.68 |
| Endo-1,4-β-xylanase | A0A1L7H884 | 4144.40 | 20.77 |
| Endo-1,4-β-xylanase I | P36218 | 189.98 | 24.58 |
| Xyloglucanase | A0A024S9Z6 | 192.46 | 87.13 |
| β-xylosidase | Q92458 | 238.57 | 87.19 |
| β-xylanase | Q9P973 | 143.56 | 38.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, X.; Peng, H.; Li, T.; Liu, P.; Gao, H.; Wang, Y.; Tang, J.; Li, Q.; Qi, Z.; et al. Full-Chain FeCl3 Catalyzation Is Sufficient to Boost Cellulase Secretion and Cellulosic Ethanol along with Valorized Supercapacitor and Biosorbent Using Desirable Corn Stalk. Molecules 2023, 28, 2060. https://doi.org/10.3390/molecules28052060
Liu J, Zhang X, Peng H, Li T, Liu P, Gao H, Wang Y, Tang J, Li Q, Qi Z, et al. Full-Chain FeCl3 Catalyzation Is Sufficient to Boost Cellulase Secretion and Cellulosic Ethanol along with Valorized Supercapacitor and Biosorbent Using Desirable Corn Stalk. Molecules. 2023; 28(5):2060. https://doi.org/10.3390/molecules28052060
Chicago/Turabian StyleLiu, Jingyuan, Xin Zhang, Hao Peng, Tianqi Li, Peng Liu, Hairong Gao, Yanting Wang, Jingfeng Tang, Qiang Li, Zhi Qi, and et al. 2023. "Full-Chain FeCl3 Catalyzation Is Sufficient to Boost Cellulase Secretion and Cellulosic Ethanol along with Valorized Supercapacitor and Biosorbent Using Desirable Corn Stalk" Molecules 28, no. 5: 2060. https://doi.org/10.3390/molecules28052060
APA StyleLiu, J., Zhang, X., Peng, H., Li, T., Liu, P., Gao, H., Wang, Y., Tang, J., Li, Q., Qi, Z., Peng, L., & Xia, T. (2023). Full-Chain FeCl3 Catalyzation Is Sufficient to Boost Cellulase Secretion and Cellulosic Ethanol along with Valorized Supercapacitor and Biosorbent Using Desirable Corn Stalk. Molecules, 28(5), 2060. https://doi.org/10.3390/molecules28052060