
Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
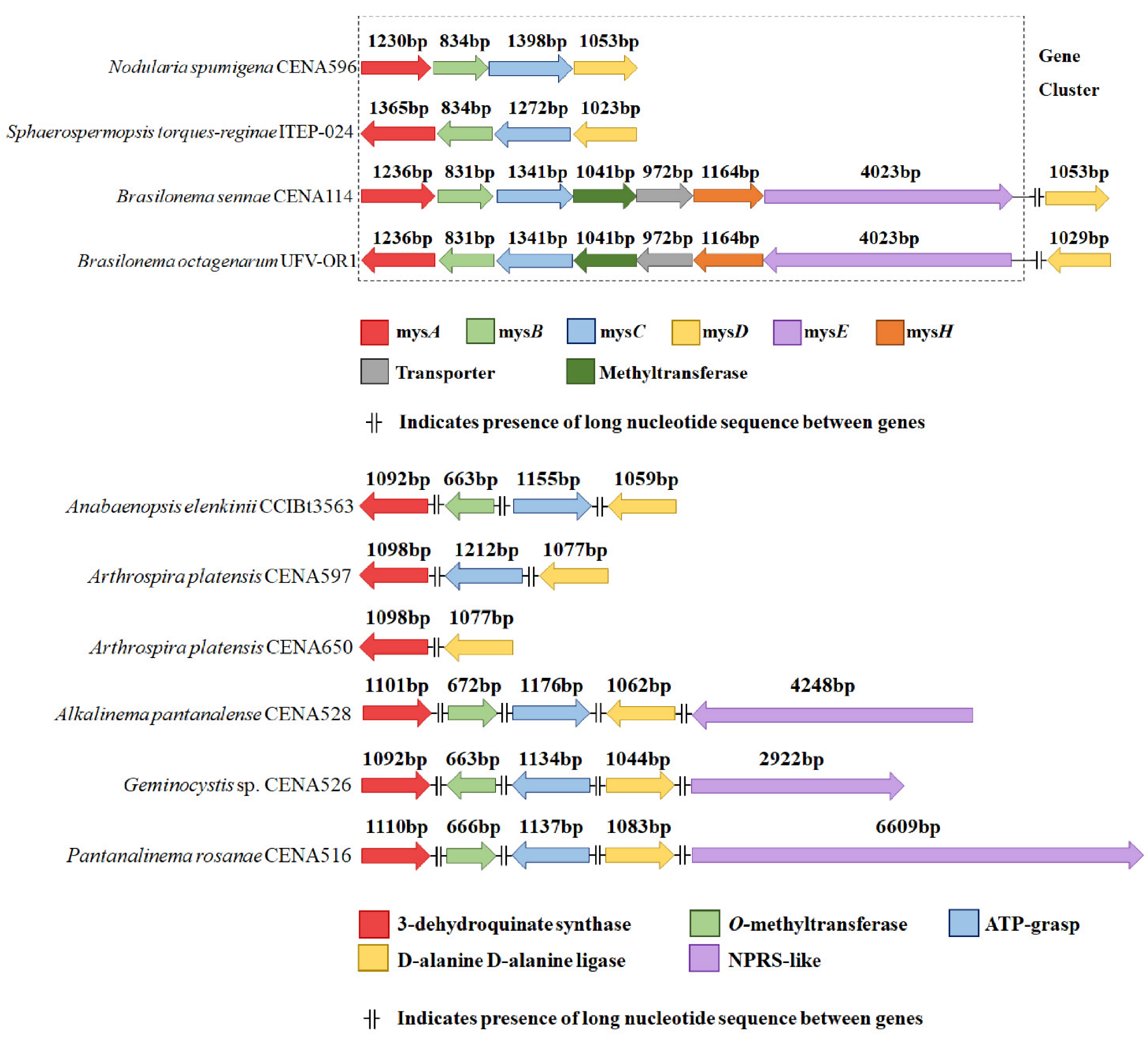
2.1. Gene Cluster Manual Annotation
2.2. Analysis of MAA and Carotenoid Content
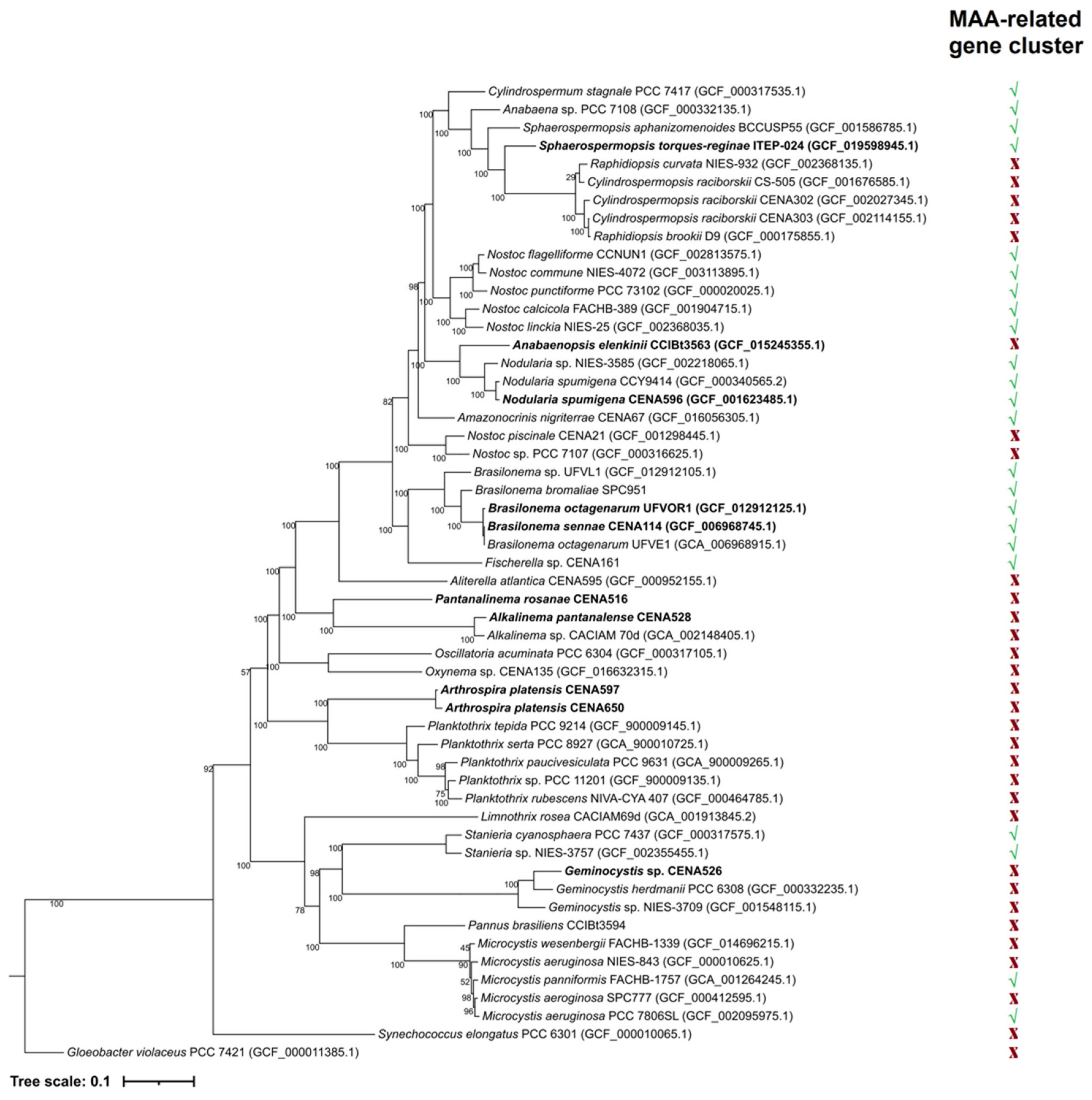
2.3. Phylogenetic Analyses
3. Materials and Methods
3.1. Cyanobacterial Strains
3.2. MAA Induction via UVA+UVB
3.3. Gene Cluster Manual Annotation
3.4. MAA Extraction and Analysis
3.5. Phylogenetic Analyses
3.6. Total Carotenoid Content Determination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Castenholz, R.W. Cyanobacteria. Oxygenic photosynthetic bacteria. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2015; Volume 410, pp. 1–4. [Google Scholar]
- Cottingham, K.L.; Ewing, H.A.; Greer, M.L.; Carey, C.C.; Weathers, K.C. Cyanobacteria as biological drivers of lake nitrogen and phosphorus cycling. Ecosphere 2015, 6, 1–19. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D.P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Rosic, N.N. Recent advances in the discovery of novel marine natural products and mycosporine-like amino acid UV-absorbing compounds. Appl. Microbiol. Biotechnol. 2021, 105, 7053–7067. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Knowland, J. Ultraviolet radiation screening compounds. Biol. Rev. Camb. Philos. Soc. 1999, 74, 311–345. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.P.; Gacesa, R.; Long, P.F.; Young, A.R. Molecular photoprotection of human keratinocytes in vitro by the naturally occurring mycosporine-like amino acid palythine. Br. J. Dermatol. 2018, 178, 1353–1363. [Google Scholar] [CrossRef]
- Balskus, E.P.; Walsh, C.T. The genetic and molecular basis for sunscreen biosynthesis in cyanobacteria. Science 2010, 329, 1653–1656. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Genome mining of mycosporine-like amino acid (MAA) synthesizing and non-synthesizing cyanobacteria: A bioinformatics study. Genomics 2010, 95, 120–128. [Google Scholar] [CrossRef]
- Zhang, L.; Li, L.; Wu, Q. Protective effects of mycosporine-like amino acids of Synechocystis sp. PCC 6803 and their partial characterization. J. Photochem. Photobiol. B Biol. 2007, 86, 240–245. [Google Scholar] [CrossRef]
- Spence, E.; Dunlap, W.C.; Shick, J.M.; Long, P.F. Redundant pathways of sunscreen biosynthesis in a cyanobacterium. ChemBioChem 2012, 13, 531–533. [Google Scholar] [CrossRef]
- Pope, M.A.; Spence, E.; Seralvo, V.; Gacesa, R.; Heidelberger, S.; Weston, A.J.; Long, P.F. O-Methyltransferase is shared between the pentose phosphate and shikimate pathways and is essential for mycosporine-like amino acid biosynthesis in Anabaena variabilis ATCC 29413. ChemBioChem 2015, 16, 320–327. [Google Scholar] [CrossRef]
- Hu, C.; Völler, G.; Süßmuth, R.; Dittmann, E.; Kehr, J.C. Functional assessment of mycosporine-like amino acids in Microcystis aeruginosa strain PCC 7806. Environ. Microbiol. 2015, 17, 1548–1559. [Google Scholar] [CrossRef]
- Katoch, M.; Mazmouz, R.; Chau, R.; Pearson, L.A.; Pickford, R.; Neilan, B.A. Heterologous production of cyanobacterial mycosporine-like amino acids mycosporine-ornithine and mycosporine-lysine in Escherichia coli. Appl. Environ. Microbiol. 2016, 82, 6167–6173. [Google Scholar] [CrossRef]
- Gao, Q.; Garcia-Pichel, F. An ATP-grasp ligase involved in the last biosynthetic step of the iminomycosporine shinorine in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2011, 193, 5923–5928. [Google Scholar] [CrossRef]
- Lawrence, K.P.; Long, P.F.; Young, A.R. Mycosporine-like amino acids for skin photoprotection. Curr. Med. Chem. 2018, 25, 5512–5527. [Google Scholar] [CrossRef]
- Geraldes, V.; Pinto, E. Mycosporine-like amino acids (MAAs): Biology, chemistry and identification features. Pharmaceuticals 2021, 14, 63. [Google Scholar] [CrossRef]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates 1, 2. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef]
- Gorham, P.R.; McLachlan, J.; Hammer, U.T.; Kim, W.K. Isolation and culture of toxic strains of Anabaena flos-aquae (Lyngb.) de Bréb. Int. Verein. Theor. Angew. Limmol. 1964, 15, 796–804. [Google Scholar] [CrossRef]
- Popin, R.V.; Rigonato, J.; Abreu, V.A.C.; Andreote, A.P.D.; Silveira, S.B.; Odebrecht, C.; Fiore, M.F. Draft genome assembly of the bloom-forming cyanobacterium Nodularia spumigena strain CENA596 in shrimp production ponds. Genome Announc. 2016, 4, e00466–e16. [Google Scholar] [CrossRef]
- Lima, S.T.; Alvarenga, D.O.; Etchegaray, A.; Fewer, D.P.; Jokela, J.; Varani, A.M.; Sanz, M.; Dorr, F.A.; Pinto, E.; Sivonen, K.; et al. Genetic organization of anabaenopeptin and spumigin biosynthetic gene clusters in the cyanobacterium Sphaerospermopsis torques-reginae ITEP-024. ACS Chem. Biol. 2017, 12, 769–778. [Google Scholar] [CrossRef]
- Andreote, A.P.D.; Vaz, M.G.M.V.; Genuário, D.B.; Barbiero, L.; Rezende-Filho, A.T.; Fiore, M.F. Nonheterocytous cyanobacteria from Brazilian saline-alkaline lakes. J. Phycol. 2014, 50, 675–684. [Google Scholar] [CrossRef]
- Delbaje, E.; Andreote, A.P.D.; Pellegrinetti, T.A.; Cruz, R.B.; Branco, L.H.; Fiore, M.F. Phylogenomic analysis of Anabaenopsis elenkinii (Nostocales, Cyanobacteria). Int. J. Syst. Evol. Microbiol. 2021, 71, 004648. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.F.; Sant’Anna, C.L.; Azevedo, M.T.D.P.; Komárek, J.; Kaštovský, J.; Sulek, J.; Lorenzi, A.A.S. The cyanobacterial genus Brasilonema, gen. nov., a molecular and phenotypic evaluation 1. J. Phycol. 2007, 43, 789–798. [Google Scholar] [CrossRef]
- Aguiar, R.; Fiore, M.F.; Franco, M.W.; Ventrella, M.C.; Lorenzi, A.S.; Vanetti, C.A.; Alfenas, A.C. A novel epiphytic cyanobacterial species from the genus Brasilonema causing damage to Eucalyptus leaves. J. Phycol. 2008, 44, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Fuseda, Y.; Satou, K. Toward a data mining service from large and heterogeneous genome databases in GenomeNet. Genome Inform. 1999, 10, 304–305. [Google Scholar]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. The KEGG databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef]
- Geraldes, V.; de Medeiros, L.S.; Jacinavicius, F.R.; Long, P.F.; Pinto, E. Development and validation of a rapid LC-MS/MS method for the quantification of mycosporines and mycosporine-like amino acids (MAAs) from cyanobacteria. Algal Res. 2020, 46, 101796. [Google Scholar] [CrossRef]
- Chaumeil, P.A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2019, 36, 1925–1927. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny. fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Dereeper, A.; Audic, S.; Claverie, J.M.; Blanc, G. BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 2010, 10, 8. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Chevenet, F.; Brun, C.; Bañuls, A.L.; Jacq, B.; Christen, R. TreeDyn: Towards dynamic graphics and annotations for analyses of trees. BMC Bioinform. 2006, 7, 439. [Google Scholar] [CrossRef]
- Shoguchi, E. Gene clusters for biosynthesis of mycosporine-like amino acids in dinoflagellate nuclear genomes: Possible recent horizontal gene transfer between species of Symbiodiniaceae (Dinophyceae). J. Phycol. 2022, 58, 1–11. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A handbook of seawater analysis. Bull. Fish. Res. Board Can. 1968, 167, 311. [Google Scholar]
- Kirk, J.T.O.; Allen, R.L. Dependence of chloroplast pigment synthesis on protein synthesis: Effect of actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, V.; de Medeiros, L.S.; Lima, S.T.; Alvarenga, D.O.; Gacesa, R.; Long, P.F.; Fiore, M.F.; Pinto, E. Genetic and biochemical evidence for redundant pathways leading to mycosporine-like amino acid biosynthesis in the cyanobacterium Sphaerospermopsis torques-reginae ITEP-024. Harmful Algae 2020, 35, 177–187. [Google Scholar] [CrossRef]
- Wang, H.; Fewer, D.P.; Sivonen, K. Genome mining demonstrates the widespread occurrence of gene clusters encoding bacteriocins in cyanobacteria. PLoS ONE 2011, 6, e22384. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Rubin, G.M.; Jiang, G.; Raad, Z.; Ding, Y. Biosynthesis and Heterologous Production of Mycosporine-Like Amino Acid Palythines. J. Org. Chem. 2021, 86, 11160–11168. [Google Scholar] [CrossRef]
- Soule, T.; Stout, V.; Swingley, W.D.; Meeks, J.C.; Garcia-Pichel, F. Molecular genetics and genomic analysis of scytonemin biosynthesis in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2007, 189, 4465–4472. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Effects of abiotic stressors on synthesis of the mycosporine-like amino acid shinorine in the cyanobacterium Anabaena variabilis PCC 7937. Photochem. Photobiol. 2008, 84, 1500–1505. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V.; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X.; et al. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef]
- Shang, J.L.; Zhang, Z.C.; Yin, X.Y.; Chen, M.; Hao, F.H.; Wang, K.; Feng, J.L.; Xu, H.F.; Yin, Y.C.; Tang, H.R.; et al. UV-B induced biosynthesis of a novel sunscreen compound in solar radiation and desiccation tolerant cyanobacteria. Environ. Microbiol. 2018, 20, 200–213. [Google Scholar] [CrossRef]
- Le Moigne, D.; Demay, J.; Reinhardt, A.; Bernard, C.; Tiam, S.K.; Marie, B. Dynamics of the Metabolome of Aliinostoc sp. PMC 882.14 in Response to Light and Temperature Variations. Metabolites 2021, 11, 745. [Google Scholar] [CrossRef]
- Sinha, R.P.; Klisch, M.; Häder, D.P. Induction of a mycosporine-like amino acid (MAA) in the rice-field cyanobacterium Anabaena sp. by UV irradiation. J. Photochem. Photobiol. B Biol. 1999, 52, 59–64. [Google Scholar] [CrossRef]
- Mohlin, M.; Wulff, A. Interaction effects of ambient UV radiation and nutrient limitation on the toxic cyanobacterium Nodularia spumigena. Microb. Ecol. 2009, 57, 675–686. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D.; Incharoensakdi, A. Characterization and antioxidant functions of mycosporine-like amino acids in the cyanobacterium Nostoc sp. R76DM. Algal Res. 2016, 16, 110–118. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Greig, C.; Silkina, A.; Kultschar, B.; Hitchings, M.D.; Farnham, G. Mycosporine-like amino acid and aromatic amino acid transcriptome response to UV and far-red light in the cyanobacterium Chlorogloeopsis fritschii PCC 6912. Sci. Rep. 2020, 10, 20638. [Google Scholar] [CrossRef]
- Bairwa, H.K.; Prajapat, G.; Jain, S.; Khan, I.A.; Ledwani, L.; Yadav, P.; Agrawal, A. Evaluation of UV-B protection efficiency of mycosporine like amino acid extracted from the cyanobacteria Anabaenopsis sp. SLCyA isolated from a hypersaline lake. Bioresour. Technol. Rep. 2021, 15, 100749. [Google Scholar] [CrossRef]
- Boucar, M.C.M.; Shen, L.Q.; Wang, K.; Zhang, Z.C.; Qiu, B.S. UV-B irradiation enhances the production of unique mycosporine-like amino acids and carotenoids in the subaerial cyanobacterium Pseudanabaena sp. CCNU1. Eur. J. Phycol. 2021, 56, 316–323. [Google Scholar] [CrossRef]
- Werner, V.R.; Laughinghouse IV, H.D.; Fiore, M.F.; Sant’Anna, C.L.; Hoff, C.; de Souza Santos, K.R.; Neuhaus, E.B.; Molica, R.J.R.; Honda, R.Y.; Echenique, R.O. Morphological and molecular studies of Sphaerospermopsis torques-reginae (Cyanobacteria, Nostocales) from South American water blooms. Phycologia 2012, 51, 228–238. [Google Scholar] [CrossRef]
- Quesada, A.; Vincent, W.F. Strategies of adaptation by Antarctic cyanobacteria to ultraviolet radiation. Eur. J. Phycol. 1997, 32, 335–342. [Google Scholar] [CrossRef]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef]
- Karsten, U. Defense strategies of algae and cyanobacteria against solar ultraviolet radiation. In Algal Chemical Ecology; Amsler, C.D., Ed.; Springer: Berlin, Germany, 2008; pp. 273–296. [Google Scholar]
- Castenholz, R.W.; Garcia-Pichel, F. Cyanobacterial responses to UV radiation. In Ecology of Cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, Germany, 2012; pp. 481–499. [Google Scholar]
- Garcia-Pichel, F.; Mechling, M.; Castenholz, R.W. Diel migrations of microorganisms within a benthic, hypersaline mat community. Appl. Environ. Microbiol. 1994, 60, 1500–1511. [Google Scholar] [CrossRef]
- Graham, G.J. Tandem genes and clustered genes. J. Theor. Biol. 1995, 175, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Soule, T.; Palmer, K.; Gao, Q.; Potrafka, R.M.; Stout, V.; Garcia-Pichel, F. A comparative genomics approach to understanding the biosynthesis of the sunscreen scytonemin in cyanobacteria. BMC Genom. 2009, 10, 336. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Mochimaru, M. Carotenoids and carotenogenesis in cyanobacteria: Unique ketocarotenoids and carotenoid glycosides. Cell. Mol. Life Sci. 2007, 64, 2607–2619. [Google Scholar] [CrossRef] [PubMed]
- Mushir, S.; Fatma, T. Ultraviolet radiation-absorbing mycosporine-like amino acids in cyanobacterium Aulosira fertilissima: Environmental perspective and characterization. Curr. Res. J. Biol. Sci. 2011, 3, 165–171. [Google Scholar]
- Hartmann, A.; Becker, K.; Karsten, U.; Remias, D.; Ganzera, M. Analysis of mycosporine-like amino acids in selected algae and cyanobacteria by hydrophilic interaction liquid chromatography and a novel MAA from the red alga Catenella repens. Mar. Drugs 2015, 13, 6291–6305. [Google Scholar] [CrossRef]
- Aigner, S.; Herburger, K.; Holzinger, A.; Karsten, U. Epilithic Chamaesiphon (Synechococcales, Cyanobacteria) species in mountain streams of the Alps-interspecific differences in photo-physiological traits. J. Appl. Phycol. 2018, 30, 1125–1134. [Google Scholar] [CrossRef]
- Dextro, R.B.; Delbaje, E.; Cotta, S.R.; Zehr, J.P.; Fiore, M.F. Trends in Free-access Genomic Data Accelerate Advances in Cyanobacteria Taxonomy. J. Phycol. 2021, 57, 1392–1402. [Google Scholar] [CrossRef]
- Rueter, J.G.; Petersen, R.R. Micronutrient effects on cyanobacterial growth and physiology. N. Z. J. Mar. Freshwater Res. 1987, 21, 435–445. [Google Scholar] [CrossRef]
- Cordeiro, R.S.; Vaz, I.C.; Magalhaes, S.; Barbosa, F.A. Effects of nutritional conditions on lipid production by cyanobacteria. An. Acad. Bras. Cien. 2017, 89, 2021–2031. [Google Scholar] [CrossRef]
- Karentz, D. Chemical defenses of marine organisms against solar radiation exposure: UV-absorbing mycosporine-like amino acids and scytonemim. In Marine Chemical Ecology, 1st ed.; McClintock, J.B., Baker, B.J., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 481–520. [Google Scholar]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Sulfur deficiency changes mycosporine-like amino acid (MAA) composition of Anabaena variabilis PCC 7937: A possible role of sulfur in MAA bioconversion. Photochem. Photobiol. 2010, 86, 862–870. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Häder, D.P.; Sinha, R.P. Role of various growth media on shinorine (mycosporine-like amino acid) concentration and photosynthetic yield in Anabaena variabilis PCC 7937. World J. Microbiol. Biotechnol. 2008, 24, 3111–3115. [Google Scholar] [CrossRef]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring bioactive properties of marine cyanobacteria isolated from the Portuguese coast: High potential as a source of anticancer compounds. Mar. Drugs 2014, 12, 98–114. [Google Scholar] [CrossRef]
- Monteiro, M.; Costa, M.; Moreira, C.; Vasconcelos, V.M.; Baptista, M. SScreening of BMAA-producing cyanobacteria in cultured isolates and in in situ blooms. J. Appl. Phycol. 2017, 29, 879–888. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for preparation of modified nutrient solution Z8 for algae. Oikos 1972, 11, 5. [Google Scholar]
- Shiio, I.; Nakamori, S. Microbial Production of l-Threonine: Part II. Production by α-Amino-β-hydroxyvaleric Acid Resistant Mutants of Glutamate Producing Bacteria. Agric. Biol. Chem. 1970, 34, 448–456. [Google Scholar] [CrossRef]
- Waditee-Sirisattha, R.; Kageyama, H.; Sopun, W.; Tanaka, Y.; Takabe, T. Identification and upregulation of biosynthetic genes required for accumulation of mycosporine-2-glycine under salt stress conditions in the halotolerant cyanobacterium Aphanothece halophytica. Appl. Environ. Microbiol. 2014, 80, 1763–1769. [Google Scholar] [CrossRef]
- Liu, Z.; Häder, D.P.; Sommaruga, R. Occurrence of mycosporine-like amino acids (MAAs) in the bloom-forming cyanobacterium Microcystis aeruginosa. J. Plankton Res. 2004, 26, 963–966. [Google Scholar] [CrossRef]
- Burford, M.A.; Carey, C.C.; Hamilton, D.P.; Huisman, J.; Paerl, H.W.; Wood, S.A.; Wulff, A. Perspective: Advancing the research agenda for improving understanding of cyanobacteria in a future of global change. Harmful Algae 2020, 91, 101601. [Google Scholar] [CrossRef]
- Lakeman, M.B.; Von Dassow, P.; Cattolico, R.A. The strain concept in phytoplankton ecology. Harmful Algae 2009, 8, 746–758. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Culture Medium | SH (µg mg−1) | P-334 (µg mg−1) | Palythine | Myc-Gly-Ala |
|---|---|---|---|---|---|
| Nodularia spumigena CENA 596 | Z8 ASM-1 BG-11 | 0.062 ± 0.036 0.046 ± 0.018 0.066 ± 0.037 | 4.791 ± 3.639 2.586 ± 1.579 6.169 ± 4.559 | - - - | - - - |
| Sphaerospermopsis torques-reginae ITEP-024 | Z8 ASM-1 BG-11 | 3.143 ± 0.983 2.764 ± 1.322 4.068 ± 1.467 | 0.319 ± 0.179 0.256 ± 0.095 0.356 ± 0.188 | X - X | - - X |
| Geminocystis sp. CENA 526 | Z8 ASM-1 BG-11 | - - - | - - - | - - - | - - - |
| Arthrospira platensis CENA 597 | Z8 | - | - | - | - |
| Arthrospira platensis CENA 650 | Z8 | - | - | - | - |
| Pantanalinema rosanae CENA 516 | Z8 | - | - | - | - |
| Alkalinema pantanalense CENA 528 | Z8 | - | - | - | - |
| Anabaenopsis elenkinii CCIBt 3563 | Z8 modified | - | - | - | - |
| Brasilonema sennae CENA 114 | Z8 (0) | - | - | X | - |
| Brasilonema octagenarum UFV-OR1 | Z8 (0) BG-11 | - - | - - | X X | - - |
| Strain | Setting | SH (µg mg−1) | P-334 (µg mg−1) | Palythine | Myc-Gly-Ala |
|---|---|---|---|---|---|
| Nodularia spumigena CENA596 | Control 72 h after UV | 0.066 ± 0.003 0.066 ± 0.002 | 2.353 ± 0.043 * 7.658 ± 0.451 * | - X | - - |
| Sphaerospermopsis torques-reginae ITEP-024 | Control 72 h after UV | 3.849 ± 0.132 * 9.032 ± 1.121 * | 0.227 ± 0.059 * 1.699 ± 0.127 * | X 0.034 ± 0.005 | - X |
| Brasilonema octagenarum UFV-OR1 | Control 72 h after UV | - - | - - | X 0.278 ± 0.039 | - - |
| Brasilonema sennae CENA114 | Control 72 h after UV | - - | - - | X X | - - |
| Geminocystis sp. CENA526 | Control 72 h after UV | - - | - - | - - | - - |
| Alkalinema pantanalense CENA528 | Control 72 h after UV | - - | - - | - - | - - |
| Anabaenopsis elenkinii CCIBt3563 | Control 72 h after UV | - - | - - | - - | - - |
| Arthrospira platensis CENA597 | Control 72 h after UV | - - | - - | - - | - - |
| Arthrospira platensis CENA650 | Control 72 h after UV | - - | - - | - - | - - |
| Pantanalinema rosanae CENA516 | Control72 h after UV | - - | - - | - - | - - |
| Strain | Culture Medium | Habitat of Origin | Genome Assembly Accession Number | Description and Isolation Reference |
|---|---|---|---|---|
| Nodularia spumigena CENA596 | Z8 | Shrimp production pond (32°12′19″ S, 52°10′42″ W, Rio Grande, Rio Grande do Sul/Brazil) | GCA_001623485.1 | [19] |
| Sphaerospermopsis torques-reginae ITEP-024 | Z8 | Freshwater Tapacurá reservoir (8°02′14″ S 35°09′46″ W, Recife, Pernambuco/Brazil) | GCA_019598945.1 | [20] |
| Geminocystis sp. CENA526 | Z8 | Saline-alkaline lake (19°26′24″ S, 56°05′58″ W, Aquidauana, Mato Grosso do Sul/Brazil) | KF246492 (only 16S RNA available) | [21] |
| Arthrospira platensis CENA597 | Z8 | Saline-alkaline lake (19°27′3.13″ S, 56°7′42.19″ W, Aquidauana, Mato Grosso do Sul/Brazil) | Unpublished | Unpublished |
| Arthrospira platensis CENA650 | Z8 | Saline-alkaline lake (19°22′47.2″ S, 56°18′51.6″ W, Aquidauana, Mato Grosso do Sul/Brazil) | Unpublished | Unpublished |
| Pantanalinema rosanae CENA516 | Z8 | Saline-alkaline lake (19°28′13″ S, 56°03′22″ W, Aquidauana, Mato Grosso do Sul/Brazil) | KF246483 (only 16S RNA available) | [21] |
| Alkalinema pantanalense CENA528 | Z8 | Saline-alkaline lake (19°26′56.0″ S, 56°07′54.8″ W, Aquidauana, Mato Grosso do Sul/Brazil) | KF246494 (only 16S RNA available) | [21] |
| Anabaenopsis elenkinii CCIBt3563 | Z8mod. * | Saline-alkaline lake (18°57′35″ S, 56°37′18″ W, Corumbá, Mato Grosso do Sul/Brazil) | GCA_015245355.1 | [22] |
| Brasilonema sennae CENA114 | Z80 ** | Iron water pipe from spring water (23°46′ S, 46°18′ W, Paranapiacaba, São Paulo/Brazil) | GCA_006968745.1 | [23] |
| Brasilonema octagenarum UFV-OR1 | Z80 ** | Orchid leaf axil (20°20′ S, 41°08′ W, Venda Nova do Imigrante, Espírito Santo/Brazil) | GCA_012912125.1 | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dextro, R.B.; Delbaje, E.; Geraldes, V.; Pinto, E.; Long, P.F.; Fiore, M.F. Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria. Molecules 2023, 28, 1420. https://doi.org/10.3390/molecules28031420
Dextro RB, Delbaje E, Geraldes V, Pinto E, Long PF, Fiore MF. Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria. Molecules. 2023; 28(3):1420. https://doi.org/10.3390/molecules28031420
Chicago/Turabian StyleDextro, Rafael B., Endrews Delbaje, Vanessa Geraldes, Ernani Pinto, Paul F. Long, and Marli F. Fiore. 2023. "Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria" Molecules 28, no. 3: 1420. https://doi.org/10.3390/molecules28031420
APA StyleDextro, R. B., Delbaje, E., Geraldes, V., Pinto, E., Long, P. F., & Fiore, M. F. (2023). Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria. Molecules, 28(3), 1420. https://doi.org/10.3390/molecules28031420