


Solid-State 13C Nuclear Magnetic Resonance Study of Soluble and Insoluble β-Glucans Extracted from Candida lusitaniae


Abstract
:1. Introduction
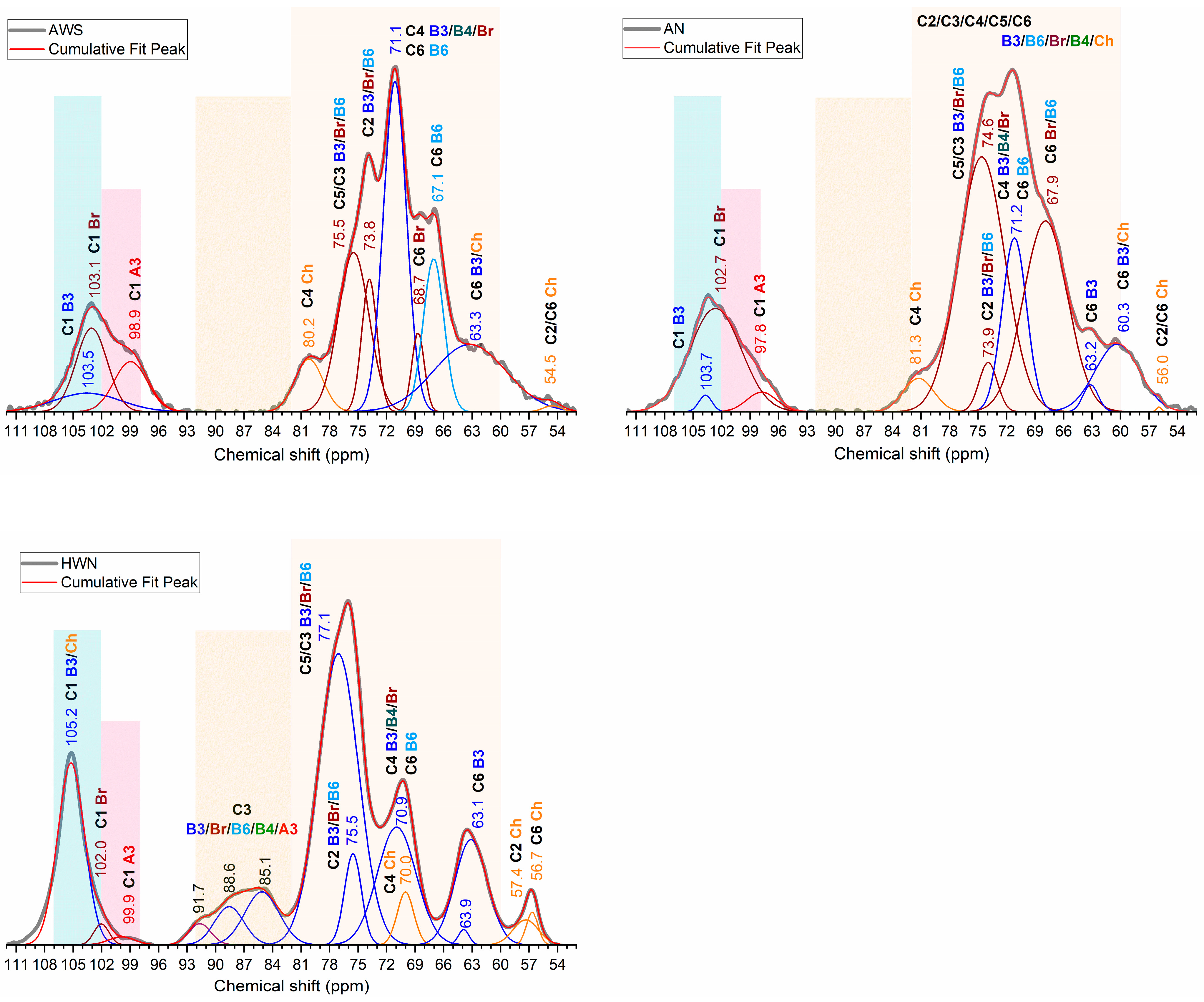
2. Results
3. Discussion
4. Materials and Methods
4.1. Yeast Strain and Culture
4.2. YeastAautolysis
4.3. Hot-Water Extraction
4.4. Alkali Extraction
4.5. Solid-State Nuclear Magnetic Resonance
4.6. Spectral Analysis, Deconvolution and Curve-Fitting
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fernando, L.D.; Dickwella Widanage, M.C.; Shekar, S.C.; Mentink-Vigier, F.; Wang, P.; Wi, S.; Wang, T. Solid-State NMR Analysis of Unlabeled Fungal Cell Walls from Aspergillus and Candida Species. J. Struct. Biol. X 2022, 6, 100070. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kirui, A.; Muszyński, A.; Widanage, M.C.D.; Chen, A.; Azadi, P.; Wang, P.; Mentink-Vigier, F.; Wang, T. Molecular Architecture of Fungal Cell Walls Revealed by Solid-State NMR. Nat. Commun. 2018, 9, 2747. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Novak, M. Structural Analysis of Glucans. Ann. Transl. Med. 2014, 2, 2. [Google Scholar] [CrossRef]
- Aimanianda, V.; Simenel, C.; Garnaud, C.; Clavaud, C.; Tada, R.; Barbin, L.; Mouyna, I.; Heddergott, C.; Popolo, L.; Ohya, Y.; et al. The Dual Activity Responsible for the Elongation and Branching of β-(1,3)-Glucan in the Fungal Cell Wall. mBio 2017, 8, e00619-17. [Google Scholar] [CrossRef]
- Manabe, N.; Yamaguchi, Y. 3D Structural Insights into β-Glucans and Their Binding Proteins. Int. J. Mol. Sci. 2021, 22, 1578. [Google Scholar] [CrossRef] [PubMed]
- Boutros, J.A.; Magee, A.S.; Cox, D. Comparison of Structural DIFFERENCES between Yeast β-Glucan Sourced from Different Strains of Saccharomyces cerevisiae and Processed Using Proprietary Manufacturing Processes. Food Chem. 2022, 367, 130708. [Google Scholar] [CrossRef] [PubMed]
- Neun, B.W.; Cedrone, E.; Potter, T.M.; Crist, R.M.; Dobrovolskaia, M.A. Detection of Beta-Glucan Contamination in Nanotechnology-Based Formulations. Molecules 2020, 25, 3367. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Q.; Wu, X.; Algharib, S.A.; Gong, F.; Hu, J.; Luo, W.; Zhou, M.; Pan, Y.; Yan, Y.; et al. Structure, Preparation, Modification, and Bioactivities of β-Glucan and Mannan from Yeast Cell Wall: A Review. Int. J. Biol. Macromol. 2021, 173, 445–456. [Google Scholar] [CrossRef]
- Lowman, D.W.; Greene, R.R.; Bearden, D.W.; Kruppa, M.D.; Pottier, M.; Monteiro, M.A.; Soldatov, D.V.; Ensley, H.E.; Cheng, S.-C.; Netea, M.G.; et al. Novel Structural Features in Candida albicans Hyphal Glucan Provide a Basis for Differential Innate Immune Recognition of Hyphae Versus Yeast. J. Biol. Chem. 2014, 289, 3432–3443. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; De Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Ibe, C.; Munro, C.A. Fungal Cell Wall Proteins and Signaling Pathways Form a Cytoprotective Network to Combat Stresses. J. Fungi 2021, 7, 739. [Google Scholar] [CrossRef] [PubMed]
- Lowman, D.W.; Sameer Al-Abdul-Wahid, M.; Ma, Z.; Kruppa, M.D.; Rustchenko, E.; Williams, D.L. Glucan and Glycogen Exist as a Covalently Linked Macromolecular Complex in the Cell Wall of Candida albicans and Other Candida Species. Cell Surf. 2021, 7, 100061. [Google Scholar] [CrossRef]
- Xin, Y.; Ji, H.; Cho, E.; Roh, K.-B.; You, J.; Park, D.; Jung, E. Immune-Enhancing Effect of Water-Soluble Beta-Glucan Derived from Enzymatic Hydrolysis of Yeast Glucan. Biochem. Biophys. Rep. 2022, 30, 101256. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Li, F.; Ding, M.; Zhang, R.; Shi, T.; Lu, Y.; Jiang, W. Molecular Dynamic Simulation: Study on the Recognition Mechanism of Linear β-(1→3)-D-Glucan by Dectin-1. Carbohydr. Polym. 2022, 286, 119276. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Verma, R.; Byun, S.; Jeun, E.-J.; Kim, G.-C.; Lee, S.; Kang, H.-J.; Kim, C.J.; Sharma, G.; Lahiri, A.; et al. Structural Specificities of Cell Surface β-Glucan Polysaccharides Determine Commensal Yeast Mediated Immuno-Modulatory Activities. Nat. Commun. 2021, 12, 3611. [Google Scholar] [CrossRef]
- Li, J.; Cai, C.; Zheng, M.; Hao, J.; Wang, Y.; Hu, M.; Fan, L.; Yu, G. Alkaline Extraction, Structural Characterization, and Bioactivities of (1→6)-β-D-Glucan from Lentinus Edodes. Molecules 2019, 24, 1610. [Google Scholar] [CrossRef]
- Lowman, D.W.; West, L.J.; Bearden, D.W.; Wempe, M.F.; Power, T.D.; Ensley, H.E.; Haynes, K.; Williams, D.L.; Kruppa, M.D. New Insights into the Structure of (1→3,1→6)-β-D-Glucan Side Chains in the Candida Glabrata Cell Wall. PLoS ONE 2011, 6, e27614. [Google Scholar] [CrossRef]
- Avramia, I.; Amariei, S. Spent Brewer’s Yeast as a Source of Insoluble β-Glucans. Int. J. Mol. Sci. 2021, 22, 825. [Google Scholar] [CrossRef]
- Chakraborty, A.; Fernando, L.D.; Fang, W.; Dickwella Widanage, M.C.; Wei, P.; Jin, C.; Fontaine, T.; Latgé, J.-P.; Wang, T. A Molecular Vision of Fungal Cell Wall Organization by Functional Genomics and Solid-State NMR. Nat. Commun. 2021, 12, 6346. [Google Scholar] [CrossRef]
- Han, B.; Baruah, K.; Cox, E.; Vanrompay, D.; Bossier, P. Structure-Functional Activity Relationship of β-Glucans from the Perspective of Immunomodulation: A Mini-Review. Front. Immunol. 2020, 11, 658. [Google Scholar] [CrossRef]
- Yuan, H.; Lan, P.; He, Y.; Li, C.; Ma, X. Effect of the Modifications on the Physicochemical and Biological Properties of β-Glucan—A Critical Review. Molecules 2019, 25, 57. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Kang, J.; Xu, Z.; Guo, Q.; Zhang, L.; Ning, H.; Cui, S.W. Triple-Helix Polysaccharides: Formation Mechanisms and Analytical Methods. Carbohydr. Polym. 2021, 262, 117962. [Google Scholar] [CrossRef]
- Du, L.; Zhang, X.; Wang, C.; Xiao, D. Preparation of Water Soluble Yeast Glucan by Four Kinds of Solubilizing Processes. Engineering 2012, 4, 184–188. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Zhou, Q.; Zhang, X.; Chen, R. Transition from Triple Helix to Coil of Lentinan in Solution Measured by SEC, Viscometry, and 13C NMR. Polym. J. 2002, 34, 443–449. [Google Scholar] [CrossRef]
- Ehren, H.L.; Appels, F.V.W.; Houben, K.; Renault, M.A.M.; Wösten, H.A.B.; Baldus, M. Characterization of the Cell Wall of a Mushroom Forming Fungus at Atomic Resolution Using Solid-State NMR Spectroscopy. Cell Surf. 2020, 6, 100046. [Google Scholar] [CrossRef]
- Kremmyda, A.; MacNaughtan, W.; Arapoglou, D.; Eliopoulos, C.; Metafa, M.; Harding, S.E.; Israilides, C. The Detection, Purity and Structural Properties of Partially Soluble Mushroom and Cereal β-D-Glucans: A Solid-State NMR Study. Carbohydr. Polym. 2021, 266, 118103. [Google Scholar] [CrossRef]
- Lamon, G.; Lends, A.; Valsecchi, I.; Wong, S.S.W.; Duprès, V.; Lafont, F.; Tolchard, J.; Schmitt, C.; Mallet, A.; Grélard, A.; et al. Solid-State NMR Molecular Snapshots of Aspergillus fumigatus Cell Wall Architecture during a Conidial Morphotype Transition. Proc. Natl. Acad. Sci. USA 2023, 120, e2212003120. [Google Scholar] [CrossRef]
- Saito, H.; Yokoi, M. High-Resolution 13C NMR Study of (1→3)-β-D-Glucans in the Solid State: DMSO-Induced Conformational Change and Conformational Characterization by Spin-Relaxation Measurements. Bull. Chem. Soc. Jpn. 1989, 62, 392–398. [Google Scholar] [CrossRef]
- Wu, K.; Ma, F.; Wei, C.; Gan, F.; Du, C. Rapid Determination of Nitrate Nitrogen Isotope in Water Using Fourier Transform Infrared Attenuated Total Reflectance Spectroscopy (FTIR-ATR) Coupled with Deconvolution Algorithm. Molecules 2023, 28, 567. [Google Scholar] [CrossRef]
- Aimanianda, V.; Clavaud, C.; Simenel, C.; Fontaine, T.; Delepierre, M.; Latgé, J.-P. Cell Wall β-(1,6)-Glucan of Saccharomyces cerevisiae: Structural characterization and in situ synthesis. J. Biol. Chem. 2009, 284, 13401–13412. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Rutckeviski, R.; Villalva, M.; Abreu, H.; Soler-Rivas, C.; Santoyo, S.; Iacomini, M.; Smiderle, F.R. Isolation and Comparison of α- and β-D-Glucans from Shiitake Mushrooms (Lentinula edodes) with Different Biological Activities. Carbohydr. Polym. 2020, 229, 115521. [Google Scholar] [CrossRef]
- Usui, T.; Yamaoka, N.; Matsuda, K.; Tuzimura, K.; Sugiyama, H.; Seto, S. 13C and 1H Nuclear Magnetic Resonance for Structural Investigation of Some Glucans. Agric. Biol. Chem. 1975, 39, 1071–1076. [Google Scholar] [CrossRef]
- Kolbe, F.; Ehren, H.L.; Kohrs, S.; Butscher, D.; Reiß, L.; Baldus, M.; Brunner, E. Solid-State NMR Spectroscopic Studies of 13C,15N,29Si-Enriched Biosilica from the Marine Diatom Cyclotella cryptica. Discov. Mater. 2021, 1, 3. [Google Scholar] [CrossRef]
- Vaithanomsat, P.; Boonlum, N.; Trakunjae, C.; Apiwatanapiwat, W.; Janchai, P.; Boondaeng, A.; Phalinphattharakit, K.; Nimitkeatkai, H.; Jarerat, A. Functionality of Yeast β-Glucan Recovered from Kluyveromyces Marxianus by Alkaline and Enzymatic Processes. Polymers 2022, 14, 1582. [Google Scholar] [CrossRef]
- Okobira, T.; Miyoshi, K.; Uezu, K.; Sakurai, K.; Shinkai, S. Molecular Dynamics Studies of Side Chain Effect on the β-1,3-D-Glucan Triple Helix in Aqueous Solution. Biomacromolecules 2008, 9, 783–788. [Google Scholar] [CrossRef]
- Gopal, P.K.; Shepherd, M.G.; Sullivan, P.A. Analysis of Wall Glucans from Yeast, Hyphal and Germ-Tube Forming Cells of Candida albicans. Microbiology 1984, 130, 3295–3301. [Google Scholar] [CrossRef]
- Fairweather, J.K. Structural Characterization by 13C-NMR Spectroscopy of Products Synthesized in vitro by Polysaccharide Synthases Using 13C-Enriched Glycosyl Donors: Application to a UDP-Glucose:(1→3)-β-D-Glucan Synthase from Blackberry (Rubus fruticosus). Glycobiology 2004, 14, 775–781. [Google Scholar] [CrossRef]
- Spěváček, J.; Brus, J. Solid-State NMR Studies of Polysaccharide Systems. Macromol. Symp. 2008, 265, 69–76. [Google Scholar] [CrossRef]
- Qu, Y.; Zhao, X.; Guo, H.; Meng, Y.; Wang, Y.; Zhou, Y.; Sun, L. Structural Analysis and Macrophage Activation of a Novel Β-glucan Isolated from Cantharellus cibarius. Int. J. Mol. Med. 2021, 47, 50. [Google Scholar] [CrossRef] [PubMed]
- Kono, H.; Kondo, N.; Hirabayashi, K.; Ogata, M.; Totani, K.; Ikematsu, S.; Osada, M. Two-Dimensional NMR Data of a Water-Soluble β-(1→3,1→6)-Glucan from Aureobasidium pullulans and Schizophyllan from Schizophyllum commune. Data Brief 2017, 15, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Kollár, R.; Reinhold, B.B.; Petráková, E.; Yeh, H.J.C.; Ashwell, G.; Drgonová, J.; Kapteyn, J.C.; Klis, F.M.; Cabib, E. Architecture of the Yeast Cell Wall. J. Biol. Chem. 1997, 272, 17762–17775. [Google Scholar] [CrossRef] [PubMed]
- Bikmurzin, R.; Bandzevičiūtė, R.; Maršalka, A.; Maneikis, A.; Kalėdienė, L. FT-IR Method Limitations for β-Glucan Analysis. Molecules 2022, 27, 4616. [Google Scholar] [CrossRef] [PubMed]
- Fernando, L.D.; Dickwella Widanage, M.C.; Penfield, J.; Lipton, A.S.; Washton, N.; Latgé, J.-P.; Wang, P.; Zhang, L.; Wang, T. Structural Polymorphism of Chitin and Chitosan in Fungal Cell Walls from Solid-State NMR and Principal Component Analysis. Front. Mol. Biosci. 2021, 8, 727053. [Google Scholar] [CrossRef] [PubMed]
- Feás, X.; Vázquez-Tato, M.P.; Seijas, J.A.; Pratima, G.; Nikalje, A.; Fraga-López, F. Extraction and Physicochemical Characterization of Chitin Derived from the Asian Hornet, Vespa Velutina Lepeletier 1836 (Hym.: Vespidae). Molecules 2020, 25, 384. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O’Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; et al. The Structural Unit of Melanin in the Cell Wall of the Fungal Pathogen Cryptococcus Neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Prados-Rosales, R.; Itin, B.; Casadevall, A.; Stark, R.E. Solid-State NMR Reveals the Carbon-Based Molecular Architecture of Cryptococcus Neoformans Fungal Eumelanins in the Cell Wall. J. Biol. Chem. 2015, 290, 13779–13790. [Google Scholar] [CrossRef]
- Walker, C.A.; Gómez, B.L.; Mora-Montes, H.M.; Mackenzie, K.S.; Munro, C.A.; Brown, A.J.P.; Gow, N.A.R.; Kibbler, C.C.; Odds, F.C. Melanin Externalization in Candida albicans Depends on Cell Wall Chitin Structures. Eukaryot. Cell 2010, 9, 1329–1342. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What Do We Know About Structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak, S.; Kawarska, A.; Stasiak-Różańska, L.; Gientka, I.; Majewska, E. Evaluation of the Efficiency of Different Disruption Methods on Yeast Cell Wall Preparation for β-Glucan Isolation. Molecules 2014, 19, 20941–20961. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Q.; Cui, S.; Liu, H. A New Isolation Method of β-D-Glucans from Spent Yeast Saccharomyces cerevisiae. Food Hydrocoll. 2008, 22, 239–247. [Google Scholar] [CrossRef]
- Pengkumsri, N.; Sivamaruthi, B.S.; Sirilun, S.; Peerajan, S.; Kesika, P.; Chaiyasut, K.; Chaiyasut, C. Extraction of β-Glucan from Saccharomyces cerevisiae: Comparison of Different Extraction Methods and in Vivo Assessment of Immunomodulatory Effect in Mice. Food Sci. Technol. 2016, 37, 124–130. [Google Scholar] [CrossRef]
- Xing, Y.; Chen, C.; Sun, W.; Zhang, B.; Sang, Y.; Xiu, Z.; Dong, Y. An Environment-Friendly Approach to Isolate and Purify Glucan from Spent Cells of Recombinant Pichia pastoris and the Bioactivity Characterization of the Purified Glucan. Eng. Life Sci. 2018, 18, 227–235. [Google Scholar] [CrossRef]
- Varelas, V.; Tataridis, P.; Liouni, M.; Nerantzis, E.T. Application of Different Methods for the Extraction of Yeast β-Glucan. E-J. Sci. Technol. 2016, 15, 75–89. [Google Scholar]
- Shokri, H.; Asadi, F.; Khosravi, A.R. Isolation of β-glucan from the Cell Wall of Saccharomyces cerevisiae. Nat. Prod. Res. 2008, 22, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Ferro, M.; Mannu, A.; Panzeri, W.; Theeuwen, C.H.J.; Mele, A. An Integrated Approach to Optimizing Cellulose Mercerization. Polymers 2020, 12, 1559. [Google Scholar] [CrossRef]
- Malissa, A.; Cappa, F.; Schreiner, M.; Marchetti-Deschmann, M. Spectral Features Differentiate Aging-Induced Changes in Parchment—A Combined Approach of UV/VIS, µ-ATR/FTIR and µ-Raman Spectroscopy with Multivariate Data Analysis. Molecules 2023, 28, 4584. [Google Scholar] [CrossRef]
- Geminiani, L.; Campione, F.; Corti, C.; Luraschi, M.; Motella, S.; Recchia, S.; Rampazzi, L. Differentiating between Natural and Modified Cellulosic Fibres Using ATR-FTIR Spectroscopy. Heritage 2022, 5, 4114–4139. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Carbon Assignment | Fraction | C1 | C2 | C3 | C4 | C5 | C6 | CH3 | C=O |
|---|---|---|---|---|---|---|---|---|---|
| β-1,3-glucans | AN | 103.7 | 73.9 | n.d. * | 71.2 | 74.6 | 63.2 | ||
| AWS | 103.5 | 73.8 | n.d. * | 71.1 | 75.5 | 63.3 | |||
| HWN | 105.2 | 75.5 | 82–92 | 70.9 | 77.1 | 63.1 | |||
| β-1,3/1,6-glucans | AN | 102.7 | ov. ** | 67.9 | |||||
| AWS | 103.1 | 68.7 | |||||||
| HWN | 102.0 | 70.9 | |||||||
| Chitin | AN | ov. ** | 56.0 | ov. ** | 81.3 | ov. ** | 60.3 | 25.3 | 181.1 |
| AWS | ov. ** | 54.5 | ov. ** | 80.2 | ov. ** | 60.3 | 25.3 | 181.7 | |
| HWN | ov. ** | 57.4 | ov. ** | 70.0 | ov. ** | 56.7 | 24.3 | 175.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bikmurzin, R.; Maršalka, A.; Kalėdienė, L. Solid-State 13C Nuclear Magnetic Resonance Study of Soluble and Insoluble β-Glucans Extracted from Candida lusitaniae. Molecules 2023, 28, 8066. https://doi.org/10.3390/molecules28248066
Bikmurzin R, Maršalka A, Kalėdienė L. Solid-State 13C Nuclear Magnetic Resonance Study of Soluble and Insoluble β-Glucans Extracted from Candida lusitaniae. Molecules. 2023; 28(24):8066. https://doi.org/10.3390/molecules28248066
Chicago/Turabian StyleBikmurzin, Ruslan, Arūnas Maršalka, and Lilija Kalėdienė. 2023. "Solid-State 13C Nuclear Magnetic Resonance Study of Soluble and Insoluble β-Glucans Extracted from Candida lusitaniae" Molecules 28, no. 24: 8066. https://doi.org/10.3390/molecules28248066
APA StyleBikmurzin, R., Maršalka, A., & Kalėdienė, L. (2023). Solid-State 13C Nuclear Magnetic Resonance Study of Soluble and Insoluble β-Glucans Extracted from Candida lusitaniae. Molecules, 28(24), 8066. https://doi.org/10.3390/molecules28248066