
Biological Activity of Canned Pork Meat Fortified Black Currant Leaf Extract: In Vitro, In Silico, and Molecular Docking Study



Abstract
:1. Introduction
2. Results and Discussion
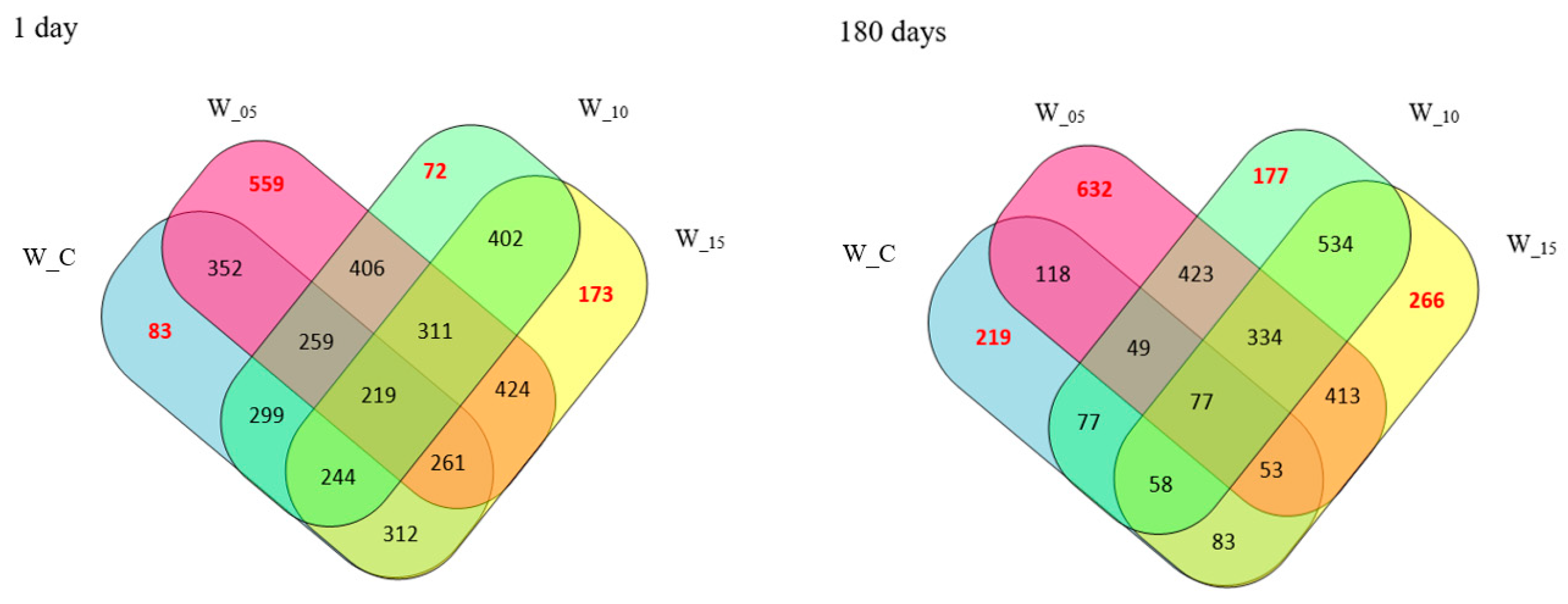
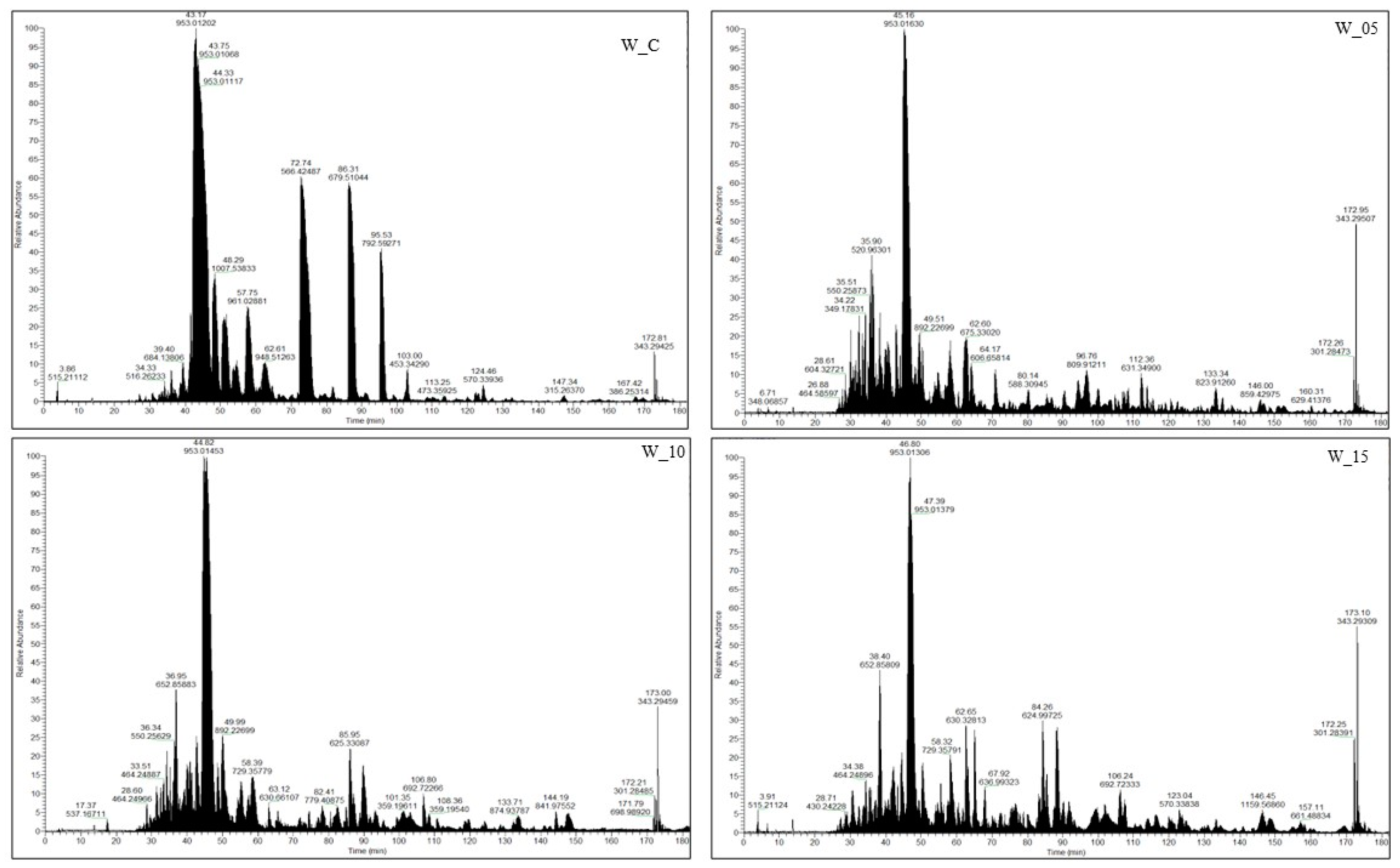
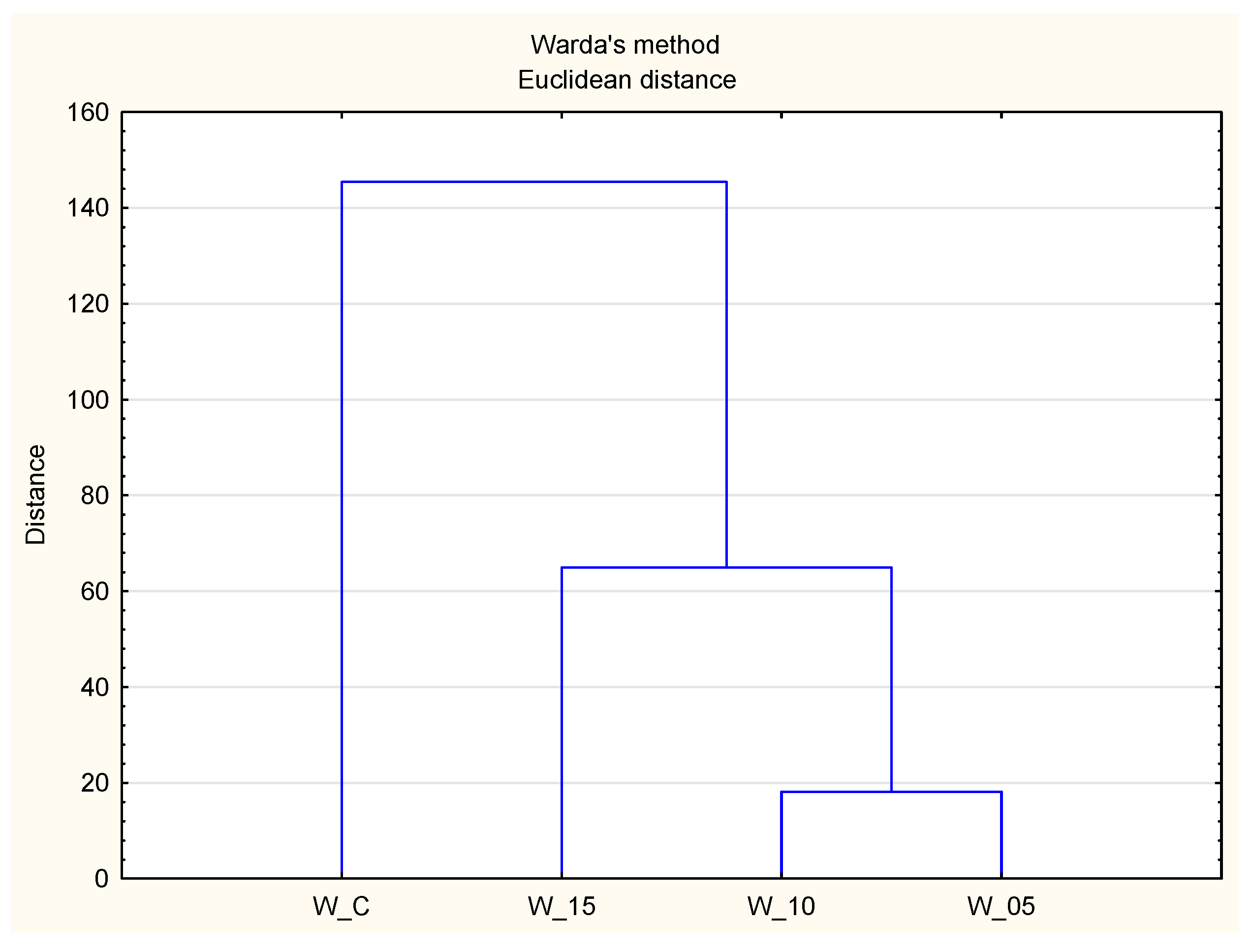
2.1. Spectrometric Characteristic of Peptides Isolated from Canned Meat with Black Currant Leaf Extract
2.2. Antioxidant Properties of Peptides: In Vitro Analysis
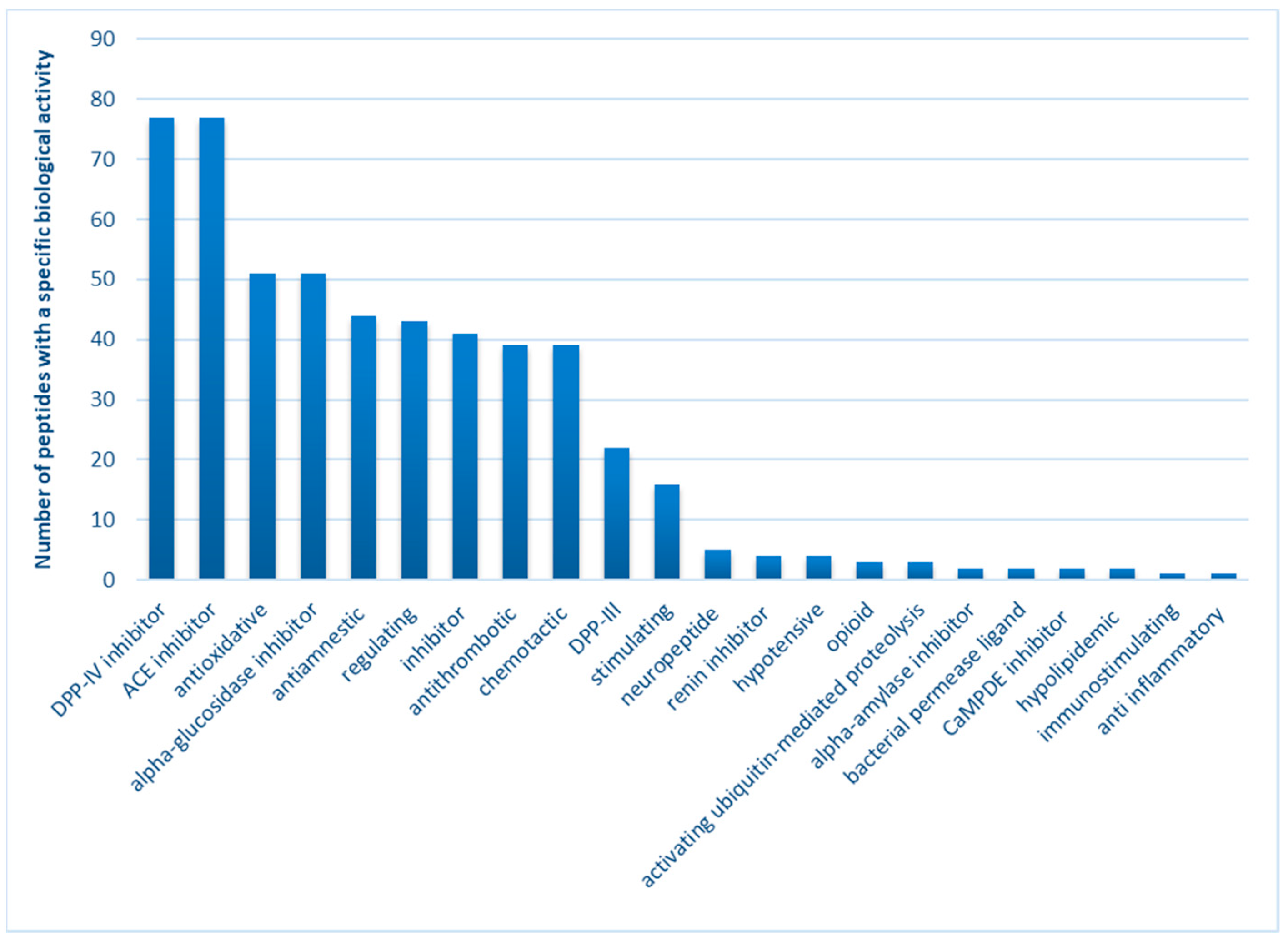
2.3. Bioactive Properties of Peptides from W_C and W_15: In Silico Analysis
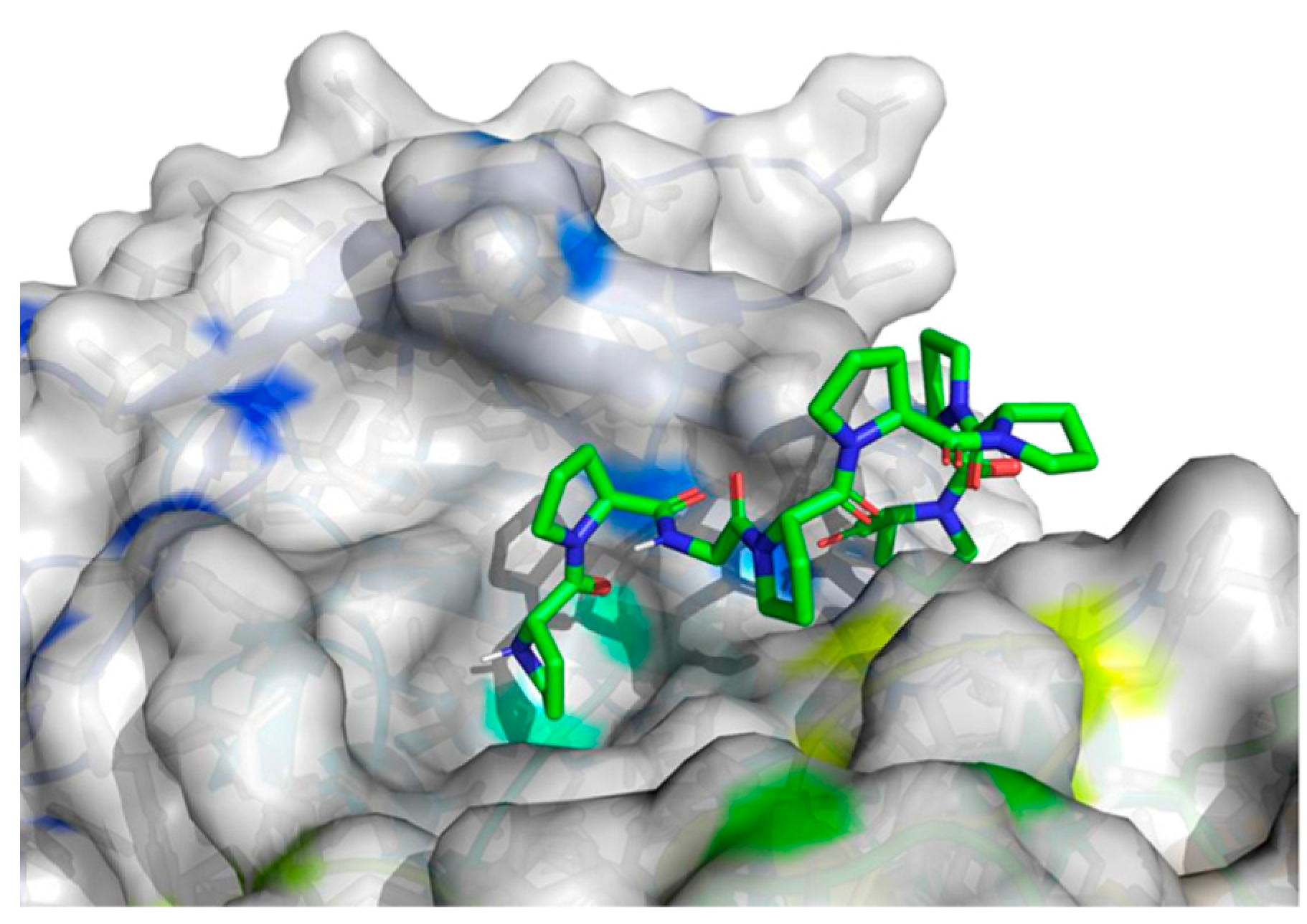
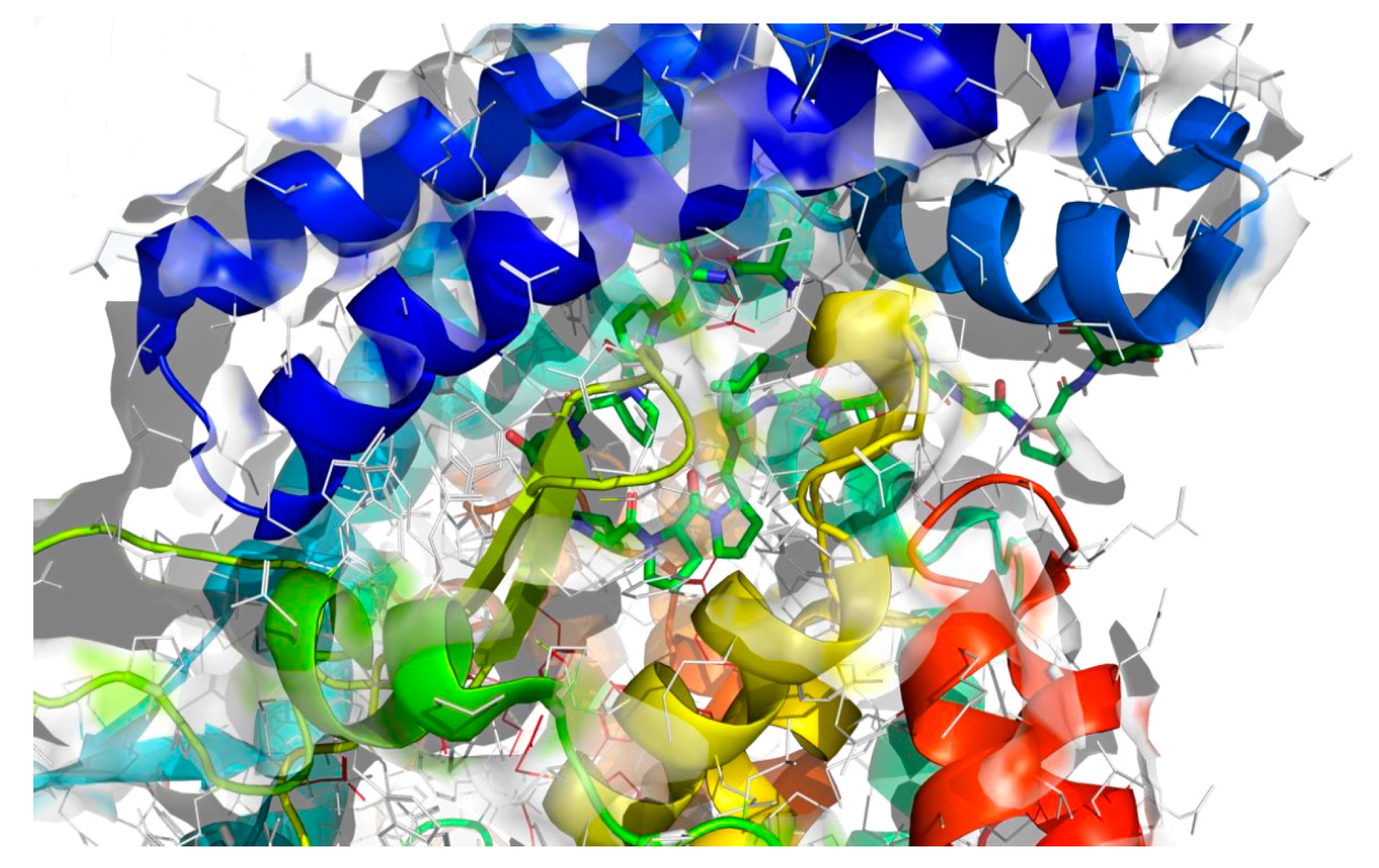
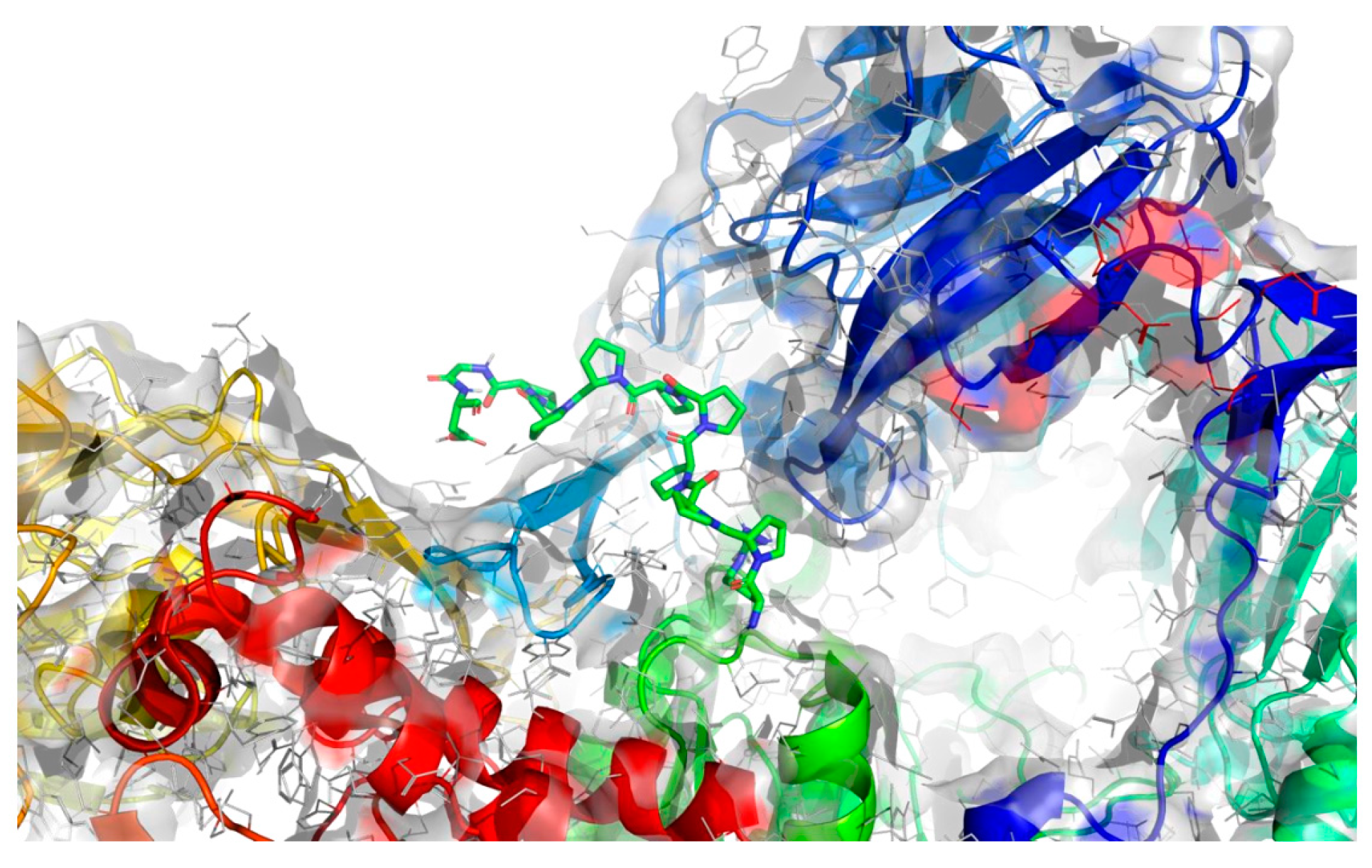
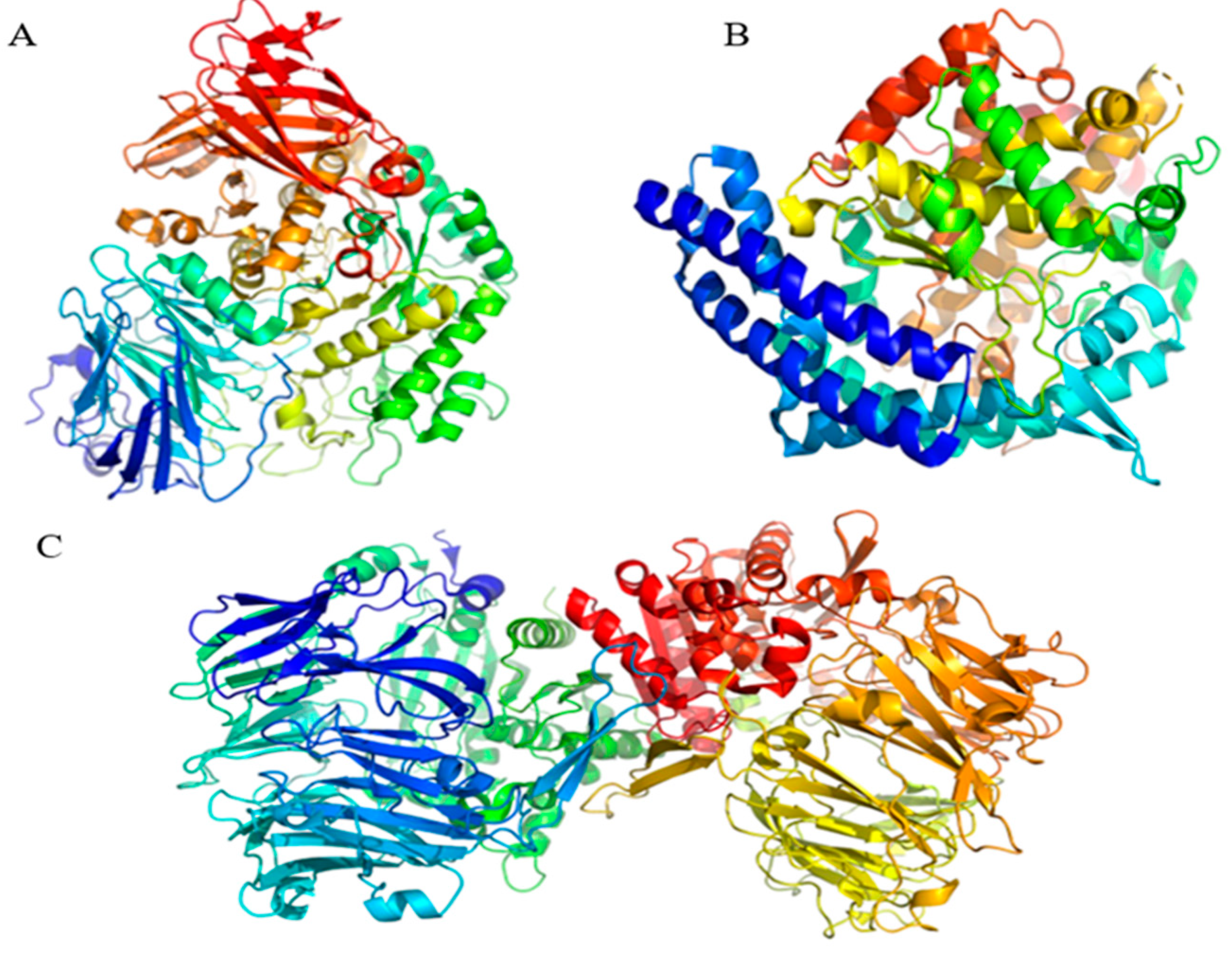
2.4. Molecular Docking Analysis
3. Materials and Methods
3.1. Extract Preparation
3.2. Canned Meat Product Preparation
3.3. Peptides Isolation and Identification
3.4. Multifunctional Properties of Processed Meat Peptides with Currant Leaf Extract—In Vivo Analysis
3.4.1. Evaluation of Biofunctionality of Peptides
3.4.2. Molecular Docking
3.5. Antioxidant Properties of Peptides from Meat Products with Currant Leaf Extract—In Vitro Analysis
3.5.1. The Ability to Neutralize the ABTS Radicals
3.5.2. Ability to Chelate Iron (II) Ions
3.5.3. Ability to Reduce Iron (III) Ions (Reducing Power)
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Godfray, H.C.J.; Aveyard, P.; Garnett, T.; Hall, J.W.; Key, T.J.; Lorimer, J.; Pierrehumbert, R.T.; Scarborough, P.; Springmann, M.; Jebb, S.A. Meat consumption, health, and the environment. Science 2018, 361, eaam5324. [Google Scholar] [CrossRef] [PubMed]
- Geiker, N.R.W.; Bertram, H.C.; Mejborn, H.; Dragsted, L.O.; Kristensen, L.; Carrascal, J.R.; Bügel, S.; Astrup, A. Meat and human health—Current knowledge and research gaps. Foods 2021, 10, 1556. [Google Scholar] [CrossRef] [PubMed]
- Olmedilla-Alonso, B.; Jiménez-Colmenero, F.; Sánchez-Muniz, F.J. Development and assessment of healthy properties of meat and meat products designed as functional foods. Meat Sci. 2013, 95, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Manessis, G.; Kalogianni, A.I.; Lazou, T.; Moschovas, M.; Bossis, I.; Gelasakis, A.I. Plant-derived natural antioxidants in meat and meat products. Antioxidants 2020, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Bosco, S.J.D.; Mir, S.A. Plant extracts as natural antioxidants in meat and meat products. Meat Sci. 2014, 98, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-del-Río, I.; López-Ibáñez, S.; Magadán-Corpas, P.; Fernández-Calleja, L.; Pérez-Valero, Á.; Tuñón-Granda, M.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Terpenoids and polyphenols as natural antioxidant agents in food preservation. Antioxidants 2021, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant polyphenols as antioxidant and antibacterial agents for shelf-life extension of meat and meat products: Classification, structures, sources, and action mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef]
- McCarthy, T.L.; Kerry, J.P.; Kerry, J.F.; Lynch, P.B.; Buckley, D.J. Evaluation of the antioxidant potential of natural food/plant extracts as compared with synthetic antioxidants and vitamin E in raw and cooked pork patties. Meat Sci. 2001, 58, 45–52. [Google Scholar] [CrossRef]
- Nissen, L.R.; Byrne, D.V.; Bertelsen, G.; Skibsted, L.H. The antioxidative activity of plant extracts in cooked pork patties as evaluated by descriptive sensory profiling and chemical analysis. Meat Sci. 2004, 68, 485–495. [Google Scholar] [CrossRef]
- Hassan, O.; Fan, L.S. The anti-oxidation potential of polyphenol extract from cocoa leaves on mechanically deboned chicken meat (MDCM). LWT-Food Sci. Technol. 2005, 38, 315–321. [Google Scholar] [CrossRef]
- Biswas, A.K.; Chatli, M.K.; Sahoo, J. Antioxidant potential of curry (Murraya koenigii L.) and mint (Mentha spicata) leaf extracts and their effect on colour and oxidative stability of raw ground pork meat during refrigeration storage. Food Chem. 2012, 133, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Czyzowska, A.; Efenberger, M.; Krala, L. Polyphenolic extracts of cherry (Prunus cerasus L.) and blackcurrant (Ribes nigrum L.) leaves as natural preservatives in meat products. Food Microbiol. 2016, 59, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Guyon, C.; Meynier, A.; de Lamballerie, M. Protein and lipid oxidation in meat: A review with emphasis on high-pressure treatments. Trends Food Sci. Technol. 2016, 50, 131–143. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Technol. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Wójciak, K.M.; Ferysiuk, K.; Kęska, P.; Materska, M.; Chilczuk, B.; Trząskowska, M.; Kruk, M.; Kołożyn-Krajewska, D.; Domínguez, R. Reduction of Nitrite in Canned Pork through the Application of Black Currant (Ribes nigrum L.) Leaves Extract. Molecules 2023, 28, 1749. [Google Scholar] [CrossRef]
- Kulczyński, B.; Sidor, A.; Gramza-Michałowska, A. Characteristics of selected antioxidative and bioactive compounds in meat and animal origin products. Antioxidants 2019, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, Y.; Li, L.; Qi, B.; Ju, M.; Xu, Y.; Zhang, Y.; Sui, X. Covalent conjugates of anthocyanins to soy protein: Unravelling their structure features and in vitro gastrointestinal digestion fate. Food Res. Int. 2019, 120, 603–609. [Google Scholar] [CrossRef]
- Li, Y.; He, D.; Li, B.; Lund, M.N.; Xing, Y.; Wang, Y.; Li, F.; Cao, X.; Liu, Y.; Chen, X.; et al. Engineering polyphenols with biological functions via polyphenol-protein interactions as additives for functional foods. Trends Food Sci. Technol. 2021, 110, 470–482. [Google Scholar] [CrossRef]
- Ferysiuk, K.; Wójciak, K.M.; Kęska, P. Effect of willow herb (Epilobium angustifolium L.) extract addition to canned meat with reduced amount of nitrite on the antioxidant and other activities of peptides. Food Funct. 2022, 13, 3526–3539. [Google Scholar] [CrossRef]
- Inai, M.; Miura, Y.; Honda, S.; Masuda, A.; Masuda, T. Metmyoglobin reduction by polyphenols and mechanism of the conversion of metmyoglobin to oxymyoglobin by quercetin. J. Agric. Food Chem. 2014, 62, 893–901. [Google Scholar] [CrossRef]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Jongberg, S.; Skov, S.H.; Tørngren, M.A.; Skibsted, L.H.; Lund, M.N. Effect of white grape extract and modified atmosphere packaging on lipid and protein oxidation in chill stored beef patties. Food Chem. 2011, 128, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Zainudin, M.A.M.; Jongberg, S.; Lund, M.N. Combination of light and oxygen accelerates formation of covalent protein-polyphenol bonding during chill storage of meat added 4-methyl catechol. Food Chem. 2021, 334, 127611. [Google Scholar] [CrossRef]
- Liu, R.H. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xing, L.; Fu, Q.; Zhou, G.H.; Zhang, W.G. A review of antioxidant peptides derived from meat muscle and by-products. Antioxidants 2016, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, R.; Cao, S.; Zhang, W.; Guanghong, Z. Meat protein based bioactive peptides and their potential functional activity: A review. Int. J. Food Sci. Technol. 2019, 54, 1956–1966. [Google Scholar] [CrossRef]
- Tavares, T.G.; Malcata, F.X. Whey proteins as source of bioactive peptides against hypertension. In Bioactive Food Peptides in Health and Disease; IntechOpen: London, UK, 2013; p. 75. [Google Scholar]
- Mada, S.B.; Ugwu, C.P.; Abarshi, M.M. Health promoting effects of food-derived bioactive peptides: A review. Int. J. Pept. Res. Ther. 2020, 26, 831–848. [Google Scholar] [CrossRef]
- Antony, P.; Vijayan, R. Bioactive peptides as potential nutraceuticals for diabetes therapy: A comprehensive review. Int. J. Mol. Sci. 2021, 22, 9059. [Google Scholar] [CrossRef]
- Kęska, P.; Wójciak, K.M.; Stadnik, J. Bioactive peptides from beef products fermented by acid whey–in vitro and in silico study. Sci. Agric. 2019, 76, 311–320. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Batista, P.; Gomes, J.E.G.; da Silva, R.; Pintado, M.M. Screening of novel bioactive peptides from goat casein: In silico to in vitro validation. Int. J. Mol. Sci. 2022, 23, 2439. [Google Scholar] [CrossRef] [PubMed]
- Kęska, P.; Stadnik, J.; Łupawka, A.; Michalska, A. Novel α-Glucosidase Inhibitory Peptides Identified In Silico from Dry-Cured Pork Loins with Probiotics through Peptidomic and Molecular Docking Analysis. Nutrients 2023, 15, 3539. [Google Scholar] [CrossRef] [PubMed]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Le Guilloux, V.; Schmidtke, P.; Tuffery, P. Fpocket: An open source platform for ligand pocket detection. BMC Bioinform. 2009, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yuan, Y.; Pei, J.; Lai, L. CAVITY: Mapping the Druggable Binding Site. In Computer-Aided Drug Discovery. Methods in Pharmacology and Toxicology; Zhang, W., Ed.; Humana Press: New York, NY, USA, 2015. [Google Scholar]
- Kawabata, T. Detection of multiscale pockets on protein surfaces using mathematical morphology. Proteins Struct. Funct. Bioinform. 2010, 78, 1195–1211. [Google Scholar] [CrossRef]
- Vajravijayan, S.; Nandhagopal, N.; Anantha, K.D.; Gunasekaran, K. Isolation and characterization of an iridoid, Arbortristoside-C from Nyctanthes arbortristis Linn., a potential drug candidate for diabetes targeting α-glucosidase. J. Biomol. Struct. Dyn. 2022, 40, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE inhibitory peptides derived from simulated gastrointestinal digestion in vitro of sesame (Sesamum indicum L.) protein and molecular docking study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef]
- Ji, W.; Zhang, C.; Ji, H. Purification, identification and molecular mechanism of two dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from Antarctic krill (Euphausia superba) protein hydrolysate. J. Chromatogr. B 2017, 1064, 56–61. [Google Scholar] [CrossRef]
- Mora, L.; Sentandreu, M.A.; Toldrá, F. Identification of small troponin T peptides generated in dry-cured ham. Food Chem. 2010, 123, 691–697. [Google Scholar] [CrossRef]
- Available online: https://biochemia.uwm.edu.pl/biopep-uwm (accessed on 1 April 2022).
- Available online: https://www2.tulane.edu/~biochem/WW/PepDraw (accessed on 1 April 2022).
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of human lysosomal acid α-glucosidase–a guide for the treatment of Pompe disease. Nat. Commun. 2017, 8, 1111. [Google Scholar] [CrossRef]
- Natesh, R.; Schwager, S.L.; Sturrock, E.D.; Acharya, K.R. Crystal structure of the human angiotensin-converting enzyme–lisinopril complex. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef]
- Morris, G.; Huey, R.; Lindstrom, W.; Sanner, M.; Belew, R.; Goodsell, D.; Olson, A. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G. Open Babel: An open chemical toolbox. J. Chem. Inform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.M.; Alhossary, A.A.; Mu, Y.; Kwoh, C.-K. Protein-Ligand Blind Docking Using QuickVina-W With Inter-Process Spatio-Temporal Integration. Sci. Rep. 2017, 7, 15451. [Google Scholar] [CrossRef] [PubMed]
- Arnautova, Y.A.; Jagielska, A.; Scheraga, H.A. A new force field (ECEPP-05) for peptides, proteins, and organic molecules. J. Phys. Chem. B 2006, 110, 5025–5044. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on the product of browning reaction prepared from glucose amine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Time [Days] | Number of Identified Peptide Sequences | |||
|---|---|---|---|---|
| W_C | W_05 | W_10 | W_15 | |
| 1 | 530 | 1129 | 584 | 714 |
| 180 | 385 | 1188 | 821 | 900 |
| TEST | Time [Day] | W_C | W_05 | W_10 | W_15 |
|---|---|---|---|---|---|
| ABTS [%] | 1 | 23.13 ± 2.26 Bd | 46.07 ± 1.84 Bb | 36.43 ± 2.59 Bc | 50.97 ± 1.48 Ba |
| 180 | 27.42 ± 1.02 Ac | 56.81 ± 2.61 Ab | 57.07 ± 3.70 Ab | 67.36 ± 2.72 Aa | |
| Chelate iron (II) ions [%] | 1 | 15.68 ± 1.39 Bb | 16.98 ± 1.81 Bb | 20.24 ± 1.48 Ba | 21.86 ± 0.95 Ba |
| 180 | 21.41 ± 1.65 Ac | 24.19 ± 0.29 Ac | 28.89 ± 1.40 Ab | 46.09 ± 3.23 Aa | |
| Reduction power [A700] | 1 | 1.79 ± 0.006 Ba | 1.74 ± 0.08 Ba | 1.64 ± 0.03 Bb | 1.55 ± 0.02 Bc |
| 180 | 1.80 ± 0.008 Ac | 2.05 ± 0.05 Aa | 1.96 ± 0.05 Ab | 1.81 ± 0.05 Ac |
| No. | Peptides Sequence | A Parameter |
|---|---|---|
| DPP IV inhibiting activity | ||
| 1 | RPPPPPPPPAD 1 | 1.364 |
| 2 | PPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 1.278 |
| 3 | PPPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 1.255 |
| 4 | PPGPPPPP | 1.250 |
| 5 | PPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPP | 1.231 |
| 6 | PPPGPAPPGARPPPGPPPPGPPPPGP | 1.231 |
| 7 | APPGARPPPGPPPPPPGPSPPRPPPGPPPQ | 1.133 |
| 8 | YQEPVLGPVRGPFPILV | 1.118 |
| 9 | KPKKKPPPPAGPPPPGPPSPGP | 1.091 |
| 10 | APPGARPPPPPPPPADEPQQGPAPSGDKPKKKPPPPAGPPPPGPPSPGP | 1.082 |
| ACE-I inhibiting activity | ||
| 1 | ARPPPGPPPLGPPPPGP | 1.529 |
| 2 | PPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 1.417 |
| 3 | PPPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 1.383 |
| 4 | PGPPPPP | 1.375 |
| 5 | RPPPPPPPPAD | 1.364 |
| 6 | PPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPP | 1.359 |
| 7 | PPPGPAPPGARPPPGPPPPGPPPPGP | 1.346 |
| 8 | APPGARPPPGPPPPPPGPSPPRPPPGPPPQ | 1.267 |
| 9 | YQEPVLGPVRGPFPIIV | 1.059 |
| 10 | LLYQEPVLGPVRGPFPIIV | 1.053 |
| Alpha-glucosidase inhibiting activity | ||
| 1 | PPGPPPPP | 0.625 |
| 2 | PPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 0.472 |
| 3 | APPGARPPPGPPPPPPGPSPPRPPPGPPPQ | 0.433 |
| 4 | PPPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPPGPPPPGP | 0.426 |
| 5 | PPPGPAPPGARPPPGPPPPGPPPPGP | 0.423 |
| 6 | ARPPPGPPPLGPPPPGP | 0.412 |
| 7 | PPGPAPPGARPPPGPPPPGPPPPGPAPPGARPPPGPPPP | 0.410 |
| 8 | APPGARPPPPPPPPADEPQQGPAPSGDKPKKKPPPPAGPPPPGPPSPGP | 0.326 |
| 9 | KPKKKPPPPAGPPPPGPPSPGP | 0.318 |
| 10 | RPPPGGGPPRPPPPEESQGEGHQKRPRPPGDGPEQGP | 0.243 |
| Antioxidative activity | ||
| 1 | TLWGIQKDLKDL | 0.500 |
| 2 | LLYQEPVLGPVRGPFPIIV | 0.263 |
| 3 | LLYQEPVLGPVRGPFPILV | 0.211 |
| 4 | KPKKKPPPPAGPPPPGPPSPGP | 0.182 |
| 5 | YQEPVLGPVRGPFPIIV | 0.177 |
| 6 | PLNETVVGLYQK | 0.167 |
| 7 | AAWQKLTNAVANALAHKYH | 0.158 |
| 8 | AIRGDEELDSLIKATIAGGGVIPHIH | 0.154 |
| 9 | PLNETVVGLYQKS | 0.154 |
| 10 | LNLPTGIPIVYEL | 0.154 |
| Peptide Sequences | Peptides 3D Structure | Peptides Characteristic 1 | Receptor (PDB ID) | |
|---|---|---|---|---|
| RPPPPPPPPAD | 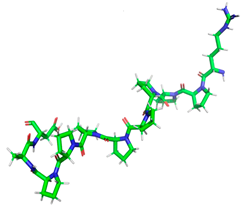 | Length: Mass: Isoelectric point: Net charge: Hydrophobicity: | 11 1136.596 6.80 0 +14.97 Kcal * mol −1 | 2QT9 |
| ARPPPGPPPLGPPPPGP | 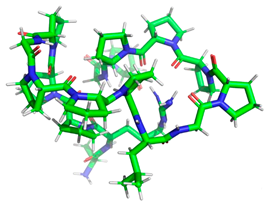 | Length: Mass: Isoelectric point: Net charge: Hydrophobicity: | 17 1596.875 11.56 +1 +19.95 Kcal * mol −1 | 1O86 |
| PPGPPPPP | 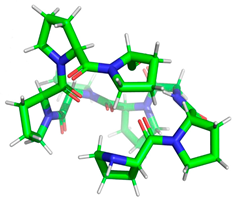 | Length: Mass: Isoelectric point: Net charge: Hydrophobicity: | 8 754.400 5.25 0 +10.03 Kcal * mol −1 | 5NN8 |
| Ligand–Receptor 3D Structure | ∆Gbinding [kcal/mol] |
|---|---|
| The best identified binding regions on the molecular surface of 5NN8 receptor with PPGPPPPP 1 | |
 | −8.4 |
| The best identified binding regions on the molecular surface of 1O86 receptor with ARPPPGPPPLGPPPPGP 2 | |
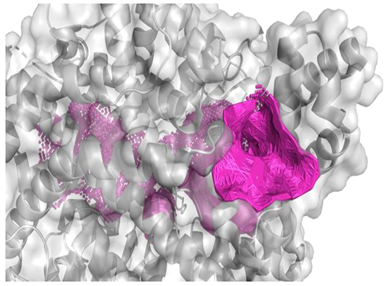 | −9.6 |
| The best identified binding regions on the molecular surface of 2QT9 receptor with RPPPPPPPPAD 3 | |
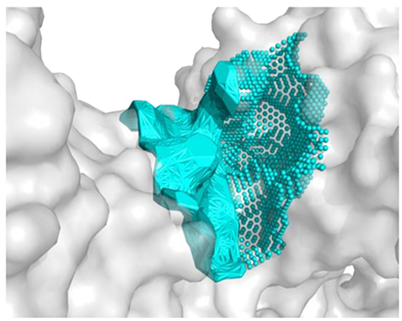 | −9.1 |
| Sample | Extract [mg/kg] | Sodium Nitrate [mg/kg] |
|---|---|---|
| W_05 | 50 | 50 |
| W_10 | 100 | 50 |
| W_15 | 150 | 50 |
| W_C (control) | - | 100 |
| Molecules | The Binding Pose for the Using Receptor is Situated in the Following Regions | References |
|---|---|---|
| α-glucosidase (PDB ID: 5NN8) | Trp376, Tyr378, Leu405, Trp481, Asp518, Met519, Phe525, Asp616, Trp618, Phe649, Leu650, His674, and Leu678. | [38] |
| Angiotensin Converting Enzyme (PDB ID: 1O86) | Ala354, Glu384, Tyr523, Gln281, His353, Lys511, His513, Tyr520, Glu162, His383, His387, Glu411, Ala356, Arg522, Glu123, Asp377, Glu376, Asp377 | [39] |
| Dipeptidyl Peptidase IV/CD26 (PDB ID: 2QT9) | Ser101, Ile102, Glu91, Asn92, Ser93, Thr94, Phe95, Asp96 | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójciak, K.M.; Kęska, P. Biological Activity of Canned Pork Meat Fortified Black Currant Leaf Extract: In Vitro, In Silico, and Molecular Docking Study. Molecules 2023, 28, 8009. https://doi.org/10.3390/molecules28248009
Wójciak KM, Kęska P. Biological Activity of Canned Pork Meat Fortified Black Currant Leaf Extract: In Vitro, In Silico, and Molecular Docking Study. Molecules. 2023; 28(24):8009. https://doi.org/10.3390/molecules28248009
Chicago/Turabian StyleWójciak, Karolina M., and Paulina Kęska. 2023. "Biological Activity of Canned Pork Meat Fortified Black Currant Leaf Extract: In Vitro, In Silico, and Molecular Docking Study" Molecules 28, no. 24: 8009. https://doi.org/10.3390/molecules28248009
APA StyleWójciak, K. M., & Kęska, P. (2023). Biological Activity of Canned Pork Meat Fortified Black Currant Leaf Extract: In Vitro, In Silico, and Molecular Docking Study. Molecules, 28(24), 8009. https://doi.org/10.3390/molecules28248009